
Chronic Stress Induces Type 2b Skeletal Muscle Atrophy via the Inhibition of mTORC1 Signaling in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Protocol
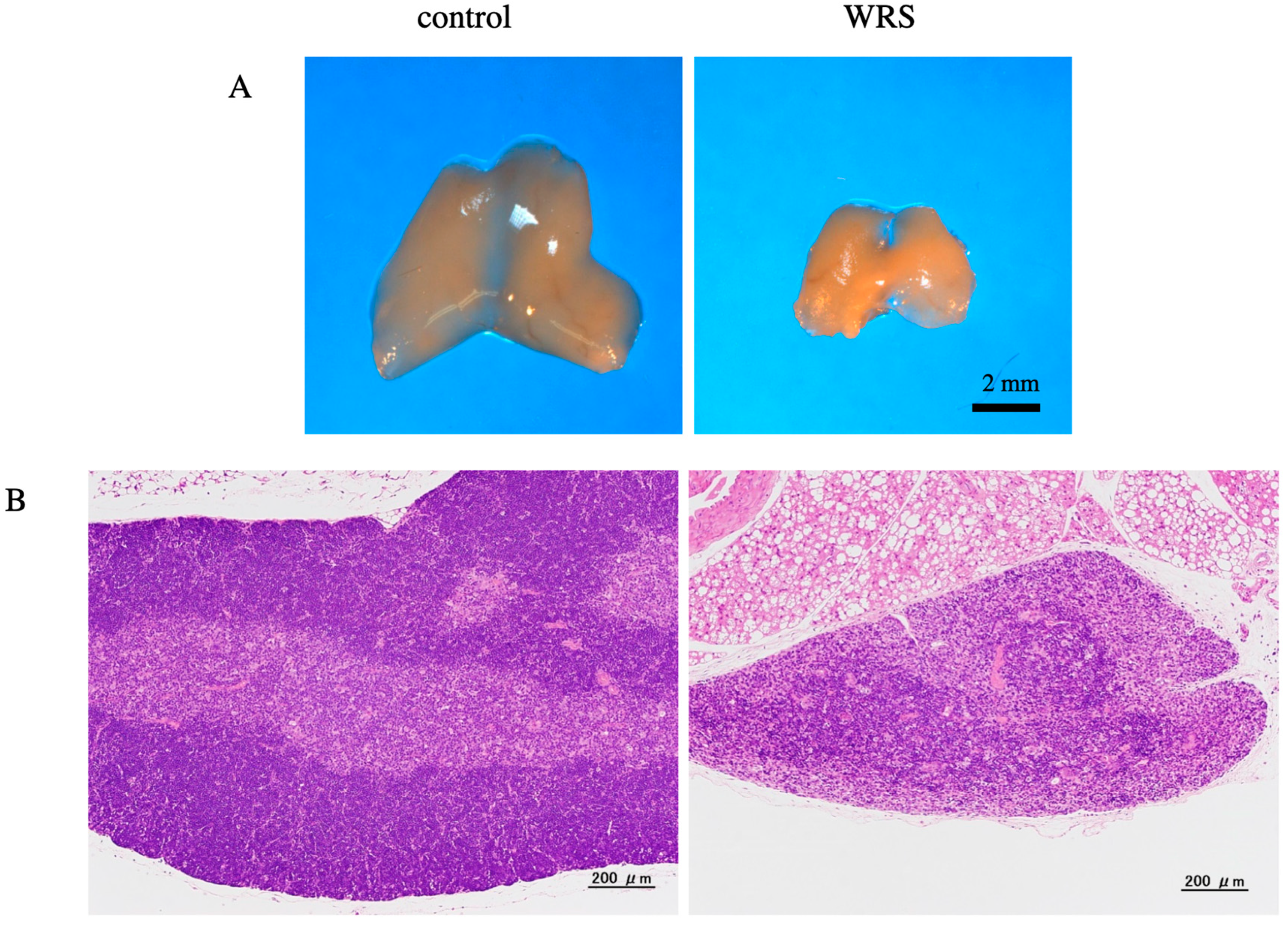
2.2. Histological Analysis of the Thymus
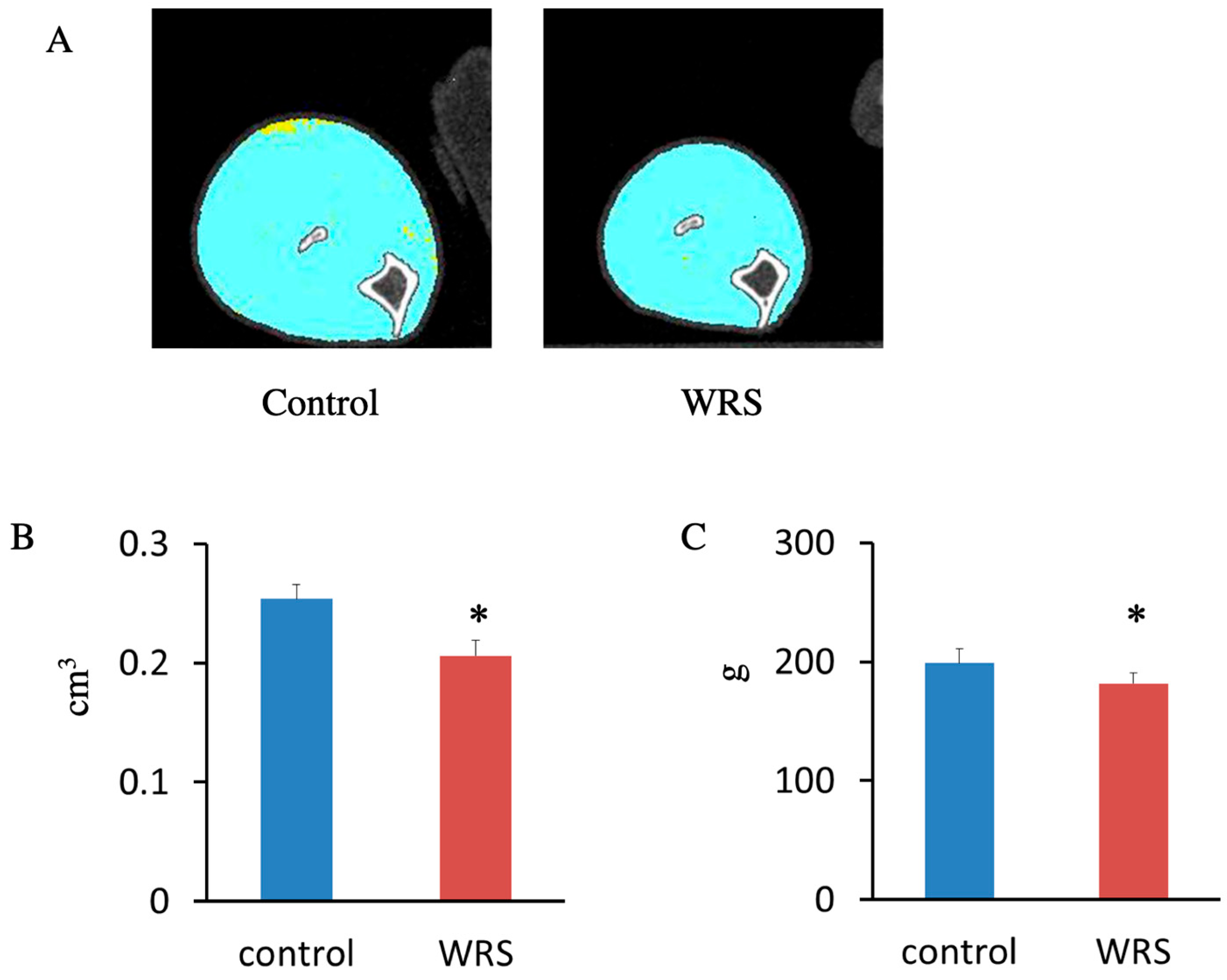
2.3. Measurement of BMD
2.4. Biochemical Markers and Bone Turnover Markers
2.5. Grip Strength Test
2.6. Measurement of Muscle Volume
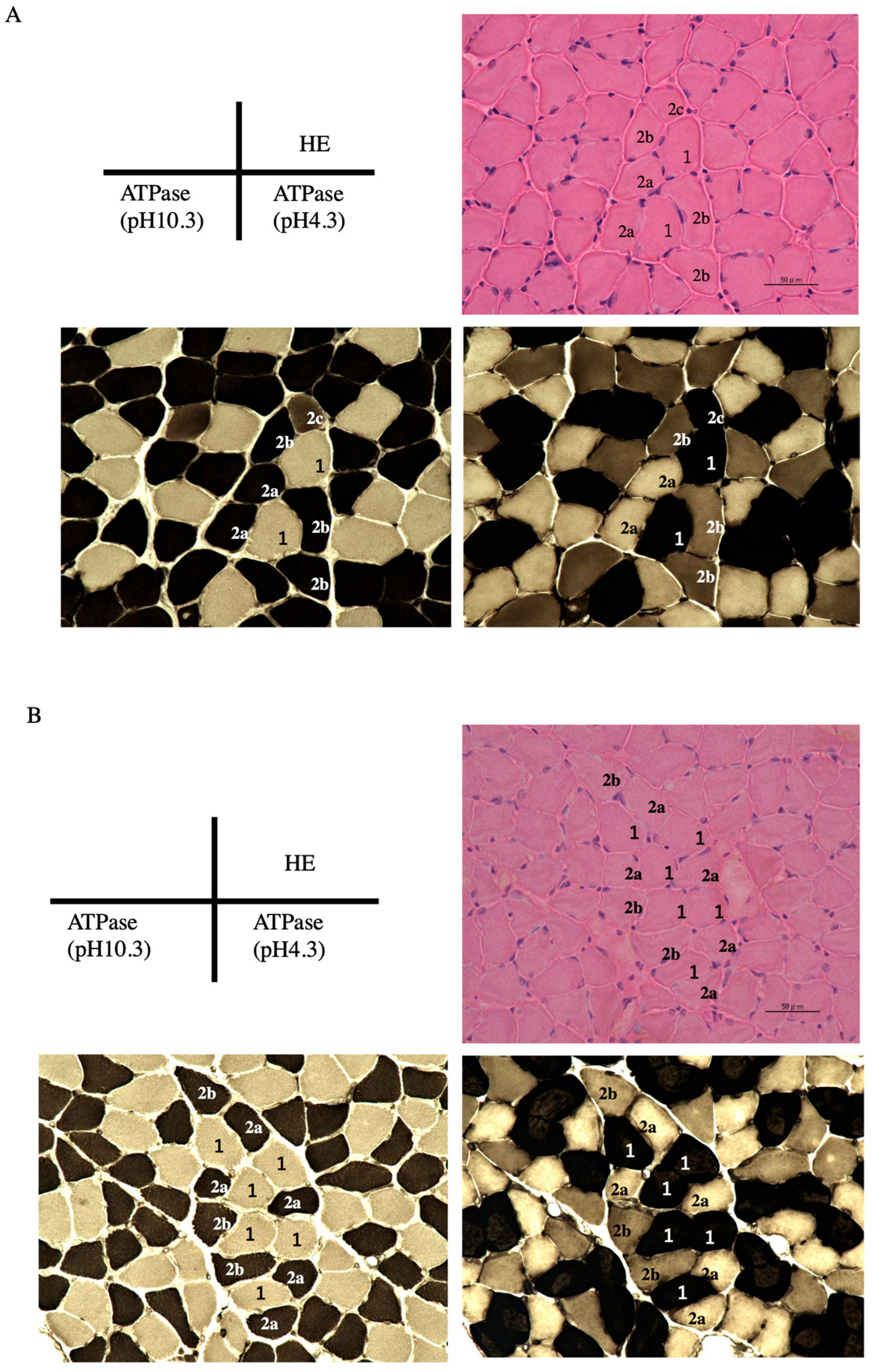
2.7. Adenosine Triphosphate (ATPase) Staining to Identify Muscle Fiber Type
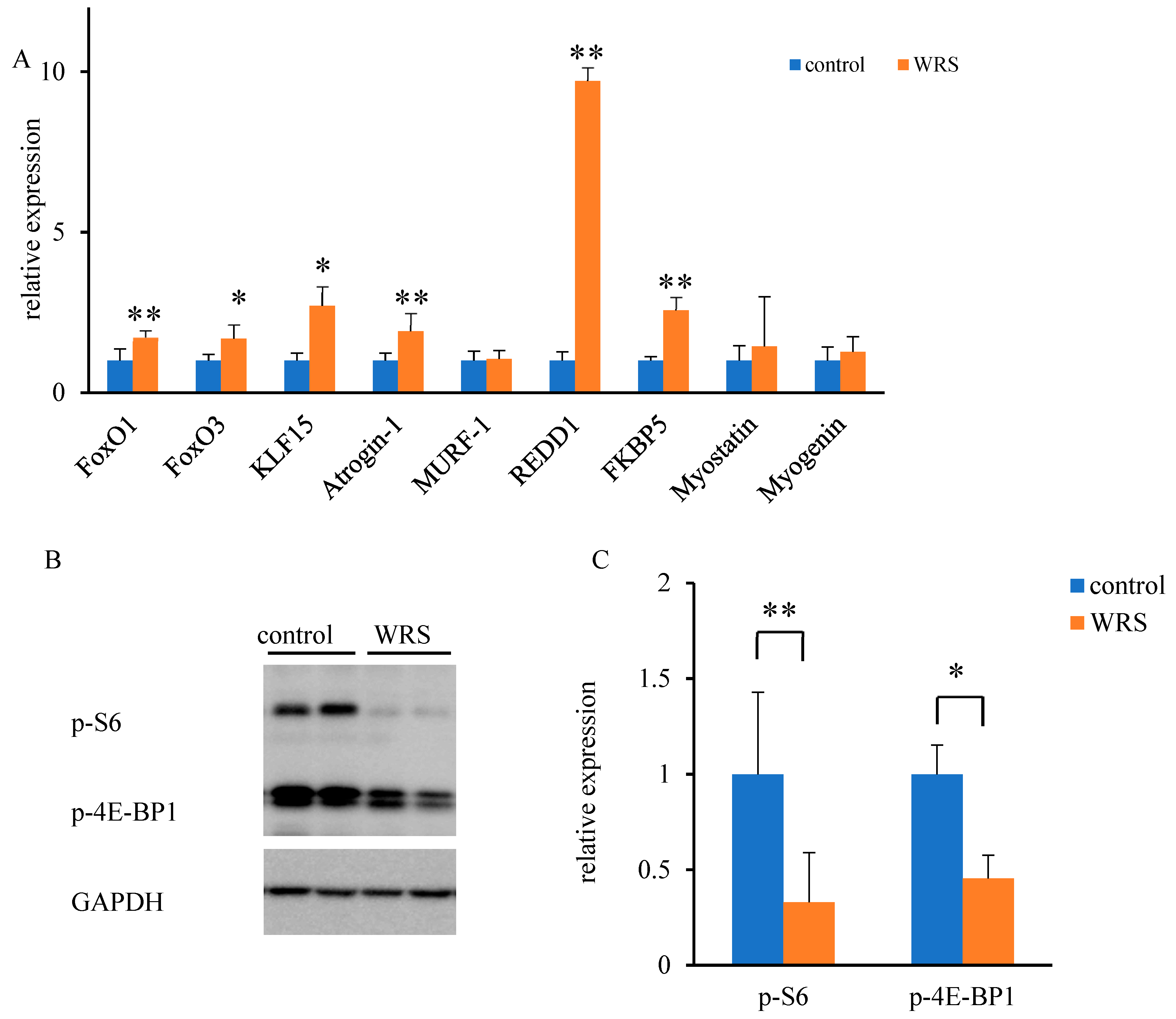
2.8. Quantitative Real-Time Polymerase Chain Reaction (PCR)
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
3.1. Influence of Chronic Stress on Mice Growth
3.2. Influence of Chronic Stress on Corticosterone Levels, Thymus Mass, and BMD
3.3. Impact of Chronic Stress on Hind Limb Muscle Size and Muscle Strength
3.4. Influence of Chronic Stress on the Soleus Muscle
3.5. Molecular Responses to Chronic Stress within the Soleus Muscle
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schakman, O.; Gilson, H.; Kalista, S.; Thissen, J.P. Mechanisms of muscle atrophy induced by glucocorticoids. Horm. Res. 2009, 72 (Suppl. 1), 36–41. [Google Scholar] [CrossRef]
- Jackman, R.W.; Kandarian, S.C. The molecular basis of skeletal muscle atrophy. Am. J. Physiol. 2004, 287, C834–C843. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.J. Skeletal muscle loss: Cachexia, sarcopenia, and inactivity. Am. J. Clin. Nutr. 2010, 91, 1123S–1127S. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J.; Uchida, H.; Kofuji, A.; Ito, J.; Shimizu, M.; Kim, H.; Sekiguchi, Y.; Kushibe, S. Molecular regulation of skeletal muscle mass and the contribution of nitric oxide: A review. FASEB BioAdvances 2019, 1, 364–374. [Google Scholar] [CrossRef]
- Mishra, S.; Cosentino, C.; Tamta, A.K.; Khan, D.; Srinivasan, S.; Ravi, V.; Abbotto, E.; Arathi, P.B.; Kumar, S.; Jain, A.; et al. Sirtuin 6 inhibition protects against glucocorticoid-induced skeletal muscle atrophy by regulating IGF/PI3K/AKT signaling. Nat. Commun. 2022, 13, 5415. [Google Scholar] [CrossRef] [PubMed]
- Cid-Diaz, T.; Leal-Lopez, S.; Fernandez-Barreiro, F.; Gonzalez-Sanchez, J.; Santos-Zas, I.; Andrade-Bulos, L.J.; Rodriguez-Fuentes, M.E.; Mosteiro, C.S.; Mouly, V.; Casabiell, X.; et al. Obestatin signaling counteracts glucocorticoid-induced skeletal muscle atrophy via NEDD4/KLF15 axis. J. Cachexia Sarcopenia Muscle 2021, 12, 493–505. [Google Scholar] [CrossRef]
- Totsuka, Y.; Nagao, Y.; Horii, T.; Yonekawa, H.; Imai, H.; Hatta, H.; Izaike, Y.; Tokunaga, T.; Atomi, Y. Physical performance and soleus muscle fiber composition in wild-derived and laboratory inbred mouse strain. J. Appl. Physiol. 2003, 95, 720–727. [Google Scholar] [CrossRef]
- Reider, L.; Beck, T.; Alley, D.; Miller, R.; Shardell, M.; Schumacher, J.; Magaziner, J.; Cawthon, P.M.; Barbour, K.E.; Cauley, J.A.; et al. Evaluating the relationship between muscle and bone modeling response in older adults. Bone 2016, 90, 152–158. [Google Scholar] [CrossRef]
- Katsuyama, H.; Fushimi, S.; Yamane, K.; Watanabe, Y.; Shimoya, K.; Okuyama, T.; Katsuyama, M.; Saijoh, K.; Tomita, M. Effect of vitamin K2 on the development of stress-induced osteopenia in a growing senescence-accelerated mouse prone 6 strain. Exper. Ther. Med. 2015, 10, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Kubo, H.; Yashiro, K.; Ohashi, K.; Tsuzuki, Y.; Wada, N.; Yamamoto, Y.; Saito, K. Effect of water-immersion restraint stress on tryptophan catabolism through the kynurenine pathway in rat tissues. J. Physiol. Sci. 2017, 67, 361–372. [Google Scholar] [CrossRef]
- Birsen, I.; Gemici, B.; Acar, N.; Ustunel, I.; Izgut-Uysal, V.N. The role of apelin in the healing of water-immersion and restraint stress-induced gastric damage. J. Physiol. Sci 2017, 67, 373–385. [Google Scholar] [CrossRef]
- Yasugaki, S.; Liu, C.Y.; Kashiwagi, M.; Kanuka, M.; Honda, T.; Miyata, S.; Yanagisawa, M.; Hayashi, Y. Effects of 3 weeks of water immersion and restraint stress on sleep in mice. Front. Neurosci. 2019, 13, 1072. [Google Scholar] [CrossRef]
- Klenerova, V.; Sery, O.; Hynie, S. Corticotropin-releasing hormone receptor subtypes in the rat anterior pituitary after two types of restraint stress. Ann. N. Y. Acad. Sci. 2008, 1148, 415–420. [Google Scholar] [CrossRef]
- Tomita, M.; Katsuyama, H.; Okuyama, T.; Watanabe, Y.; Hidaka, K.; Otsuki, T.; Nata, M. The effect of CAG repeat polymorphism in the glucocorticoid receptor on stress response of mice exposed to water-immersion restraint stress. Int. J. Mol. Med. 2010, 25, 415–420. [Google Scholar] [CrossRef]
- Fushimi, S.; Nohno, T.; Nagatsuka, H.; Katsuyama, H. Involvement of miR-140-3p in Wnt3a and TGFb3 signaling pathways during osteoblast differentiation in MC3T3-E1 cells. Genes Cells 2018, 23, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Fleshner, M.; Deak, T.; Spencer, R.L.; Laudenslager, M.L.; Watkins, L.R.; Maier, S.F. A long-term increase in basal levels of corticosterone and a decrease in corticosteroid-binding globulin after acute stressor exposure. Endocrinology 2015, 136, 5336–5342. [Google Scholar] [CrossRef] [PubMed]
- Selye, H. Forty years of stress research: Principal remaining problems and misconceptions. Can. Med. Assoc. J. 1976, 115, 53–56. [Google Scholar] [PubMed]
- Bonald, P.; Sandri, M. Cellular and molecular mechanisms of muscle atrophy. Dis. Model. Mech. 2013, 6, 25–39. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatinni, D.M. mTOR signaling in growth, metabolism, and disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Bordine, S.C.; Stitt, T.N.; Gonzalez, M.; Kline, W.O.; Stover, G.L.; Bauerlein, R.; Zlotchenko, E.; Scrimgeour, A.; Lawrence, J.C.; Glass, D.J.; et al. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nature Cell Biol. 2001, 3, 1014–1019. [Google Scholar] [CrossRef]
- Schakman, O.; Kalista, S.; Barbe, C.; Loumaye, A.; Thissen, J.P. Glucocorticoid-induced skeletal muscle atrophy. Int. J. Biochem. Cell Biol. 2013, 45, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Tong, T.; Park, T. Corticotropin releasing factor-overexpressing mouse is a model of chronic stress-induced muscle atrophy. PLoS ONE 2020, 15, e0229048. [Google Scholar] [CrossRef] [PubMed]
- Kemp, J.D.; Blazev, R.; Stephenson, D.G.; Stephenson, G.M.M. Morphological and biochemical alterations of skeletal muscles from the genetically obese (ob/ob) mouse. Int. J. Obes. 2009, 33, 831–841. [Google Scholar] [CrossRef]
- Hitomi, Y.; Kizaki, T.; Watanabe, S.; Matsumura, G.; Fujioka, Y.; Haga, S.; Izawa, T.; Taniguchi, N.; Ohno, H. Seven skeletal muscles rich in slow muscle fibers may function to sustain neural position in the rodent hindlimb. Comp. Biochem. Physiol. B 2005, 140, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Kamei, Y.; Miura, S.; Suzuki, M.; Kai, Y.; Mizukami, J.; Taniguchi, T.; Mochida, K.; Hata, T.; Matsuda, J.; Aburatani, H.; et al. Skeletal muscle FOXO1 (FKHR) transgenic mice have less skeletal muscle mass, down-regulated type I (slow twitch/red muscle) fiber genes, and impaired glycemic control. J. Biol. Chem. 2004, 279, 41114–41123. [Google Scholar] [CrossRef] [PubMed]
- Dankbar, B.; Fennen, M.; Brunert, D.; Hayer, S.; Frank, S.; Wehmeyer, C.; Beckmann, D.; Paruzel, P.; Bertrand, J.; Redlich, K.; et al. Myostain is a direct regulator of osteoclast differentiation and its inhibition reduces inflammatory joint destruction in mice. Nature Med. 2015, 21, 1085–1090. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.L.; McCall, G.E.; Loh, A.S.; Madden, M.C.; Mehan, R.S. Acute daily psychological stress causes increased atrophic gene expression and myostatin-dependent muscle atrophy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R889–R898. [Google Scholar] [CrossRef]
- Ganassi, M.; Badodi, S.; Quiroga, H.P.O.; Zammit, P.S.; Hinits, Y.; Hughes, S.M. Myogenin promotes myocyte fusion to balance fibre number and size. Nat. Comm. 2018, 9, 4232. [Google Scholar] [CrossRef]
- Depke, M.; Fusch, G.; Domanska, G.; Geffers, R.; Volker, U.; Schuett, C.; Kiank, C. Hypermetabolic syndrome as a consequence of repeated psychological stress in mice. Endocrinology 2008, 149, 2714–2723. [Google Scholar] [CrossRef]
- Paddon-Jones, D.; Sheffield-Moore, M.; Cree, M.G.; Hewlings, S.J.; Aarsland, A.; Wolfe, R.; Ferrando, A.A. Atrophy and impaired muscle protein synthesis during prolonged inactivity and stress. J. Clin. Endocrinol. Metab. 2006, 91, 4836–4841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karsenty, G.; Mera, P. Molecular basis of the crosstalk between bone and muscle. Bone 2018, 115, 43–49. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Assay ID |
|---|---|
| FoxO1 | Mm00490672_m1 |
| FoxO3 | Mm01185722_m1 |
| KLF15 | Mm00517792_m1 |
| Atrogin-1 | Mm00499523_m1 |
| MuRF1 | Mm01185221_m1 |
| REDD1 | Mm00512504_g1 |
| FKBP5 | Mm00487406_m1 |
| Myostatin | Mm01254559_m1 |
| Myogenin | Mm00446195_g1 |
| GAPDH | Mm99999915_g1 |
| Control | WRS | |
|---|---|---|
| body weight (g) | 25.2 ± 0.3 | 24.0 ± 0.4 * |
| urinary creatinine (U-CRE) (mg/dL) | 21.43 ± 4.78 | 8.67 ± 3.18 ** |
| aspartate aminotransferase (AST) (IU/L) | 155.0 ± 34.2 | 192.0 ± 53.6 |
| lactate dehydrogenase (LDH) (IU/L) | 647 ± 190 | 753 ± 290 |
| creatinine kinase (CK) (IU/L) | 1758 ± 715 | 1847 ± 889 |
| blood urea nitrogen (BUN) (mg/dL) | 36.8 ± 2.0 | 38.8 ± 8.1 |
| serum creatinine (CRE) (mg/dL) | 0.13 ± 0.02 | 0.14 ± 0.10 |
| corticosterone level (ng/mL) | ||
| day 2 | 168.1 ± 65.1 | 370.6 ± 44.1 ** |
| week 2 | 71.6 ± 36.8 | 295.0 ± 61.4 ** |
| week 4 | 82.4 ± 34.6 | 249.4 ± 45.8 ** |
| Control, n = 5 | WRS, n = 8 | |
|---|---|---|
| total BMD (mg/cm3) | 391 ± 22 | 328 ± 29 ** |
| trabecular BMD (mg/cm3) | 188 ± 15 | 146 ± 30 * |
| cortical BMD (mg/cm3) | 1052 ± 55 | 982 ± 32 * |
| ALP (IU/L) | 329.0 ± 26.3 | 402.3 ± 70.9 |
| Gla (ng/mL) | 25.8 ± 6.2 | 11.3 ± 3.5 ** |
| CTX (ng/mM Cre) | 6.1 ± 0.7 | 26.9 ± 7.9 ** |
| TRACP-5b (U/L) | 12.5 ± 1.1 | 16.4 ± 3.0 * |
| Control, n = 5 | WRS, n = 8 | |
|---|---|---|
| ratio of fiber type (%) | ||
| Type 1 | 34.0 ± 4.3 | 35.4 ± 4.3 |
| Type 2a | 46.9 ± 5.2 | 44.8 ± 2.1 |
| Type 2b | 17.4 ± 3.0 | 15.8 ± 8.7 |
| Type 2c | 1.2 ± 1.1 | 3.4 ± 5.5 |
| area of fiber type (mm2) | ||
| Type 1 | 1312 ± 156 | 1265 ± 172 |
| Type 2a | 1135 ± 115 | 990 ± 85 # |
| Type 2b | 1296 ± 152 | 1084 ± 97 * |
| Type 2c | 836 ± 446 | 532 ± 467 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fushimi, S.; Nohno, T.; Katsuyama, H. Chronic Stress Induces Type 2b Skeletal Muscle Atrophy via the Inhibition of mTORC1 Signaling in Mice. Med. Sci. 2023, 11, 19. https://doi.org/10.3390/medsci11010019
Fushimi S, Nohno T, Katsuyama H. Chronic Stress Induces Type 2b Skeletal Muscle Atrophy via the Inhibition of mTORC1 Signaling in Mice. Medical Sciences. 2023; 11(1):19. https://doi.org/10.3390/medsci11010019
Chicago/Turabian StyleFushimi, Shigeko, Tsutomu Nohno, and Hironobu Katsuyama. 2023. "Chronic Stress Induces Type 2b Skeletal Muscle Atrophy via the Inhibition of mTORC1 Signaling in Mice" Medical Sciences 11, no. 1: 19. https://doi.org/10.3390/medsci11010019