Influence of Traditional vs Alternative Dietary Carbohydrates Sources on the Large Intestinal Microbiota in Post-Weaning Piglets



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Former Food Product Ingredients
2.2. Animals, Housing and Treatment
2.3. Experimental Diets
2.4. Sample Collection
2.5. DNA Extraction and Sequencing
2.6. Growth Performance Data
2.7. NGS Data Analysis
3. Results
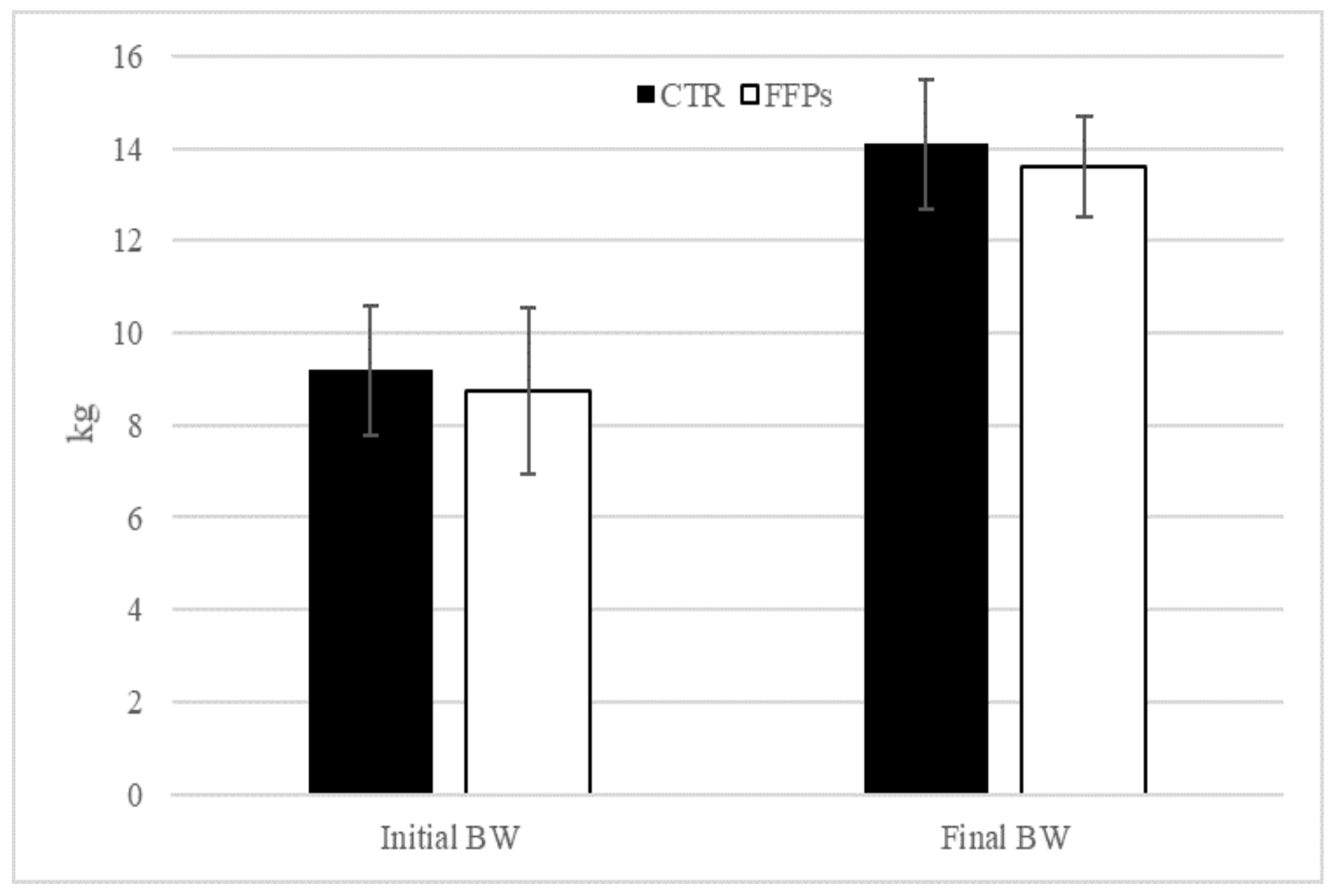
3.1. Growth Performance
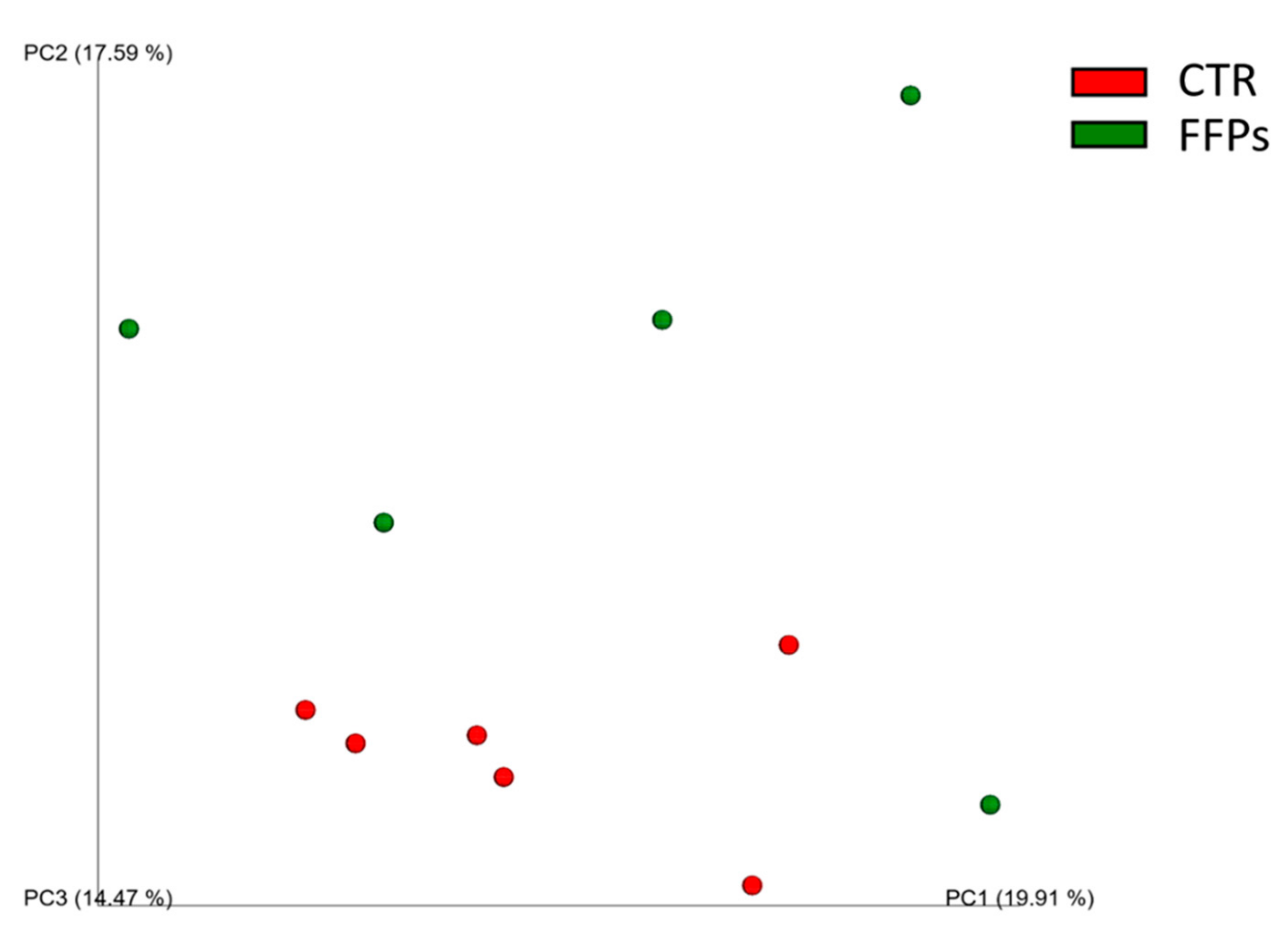
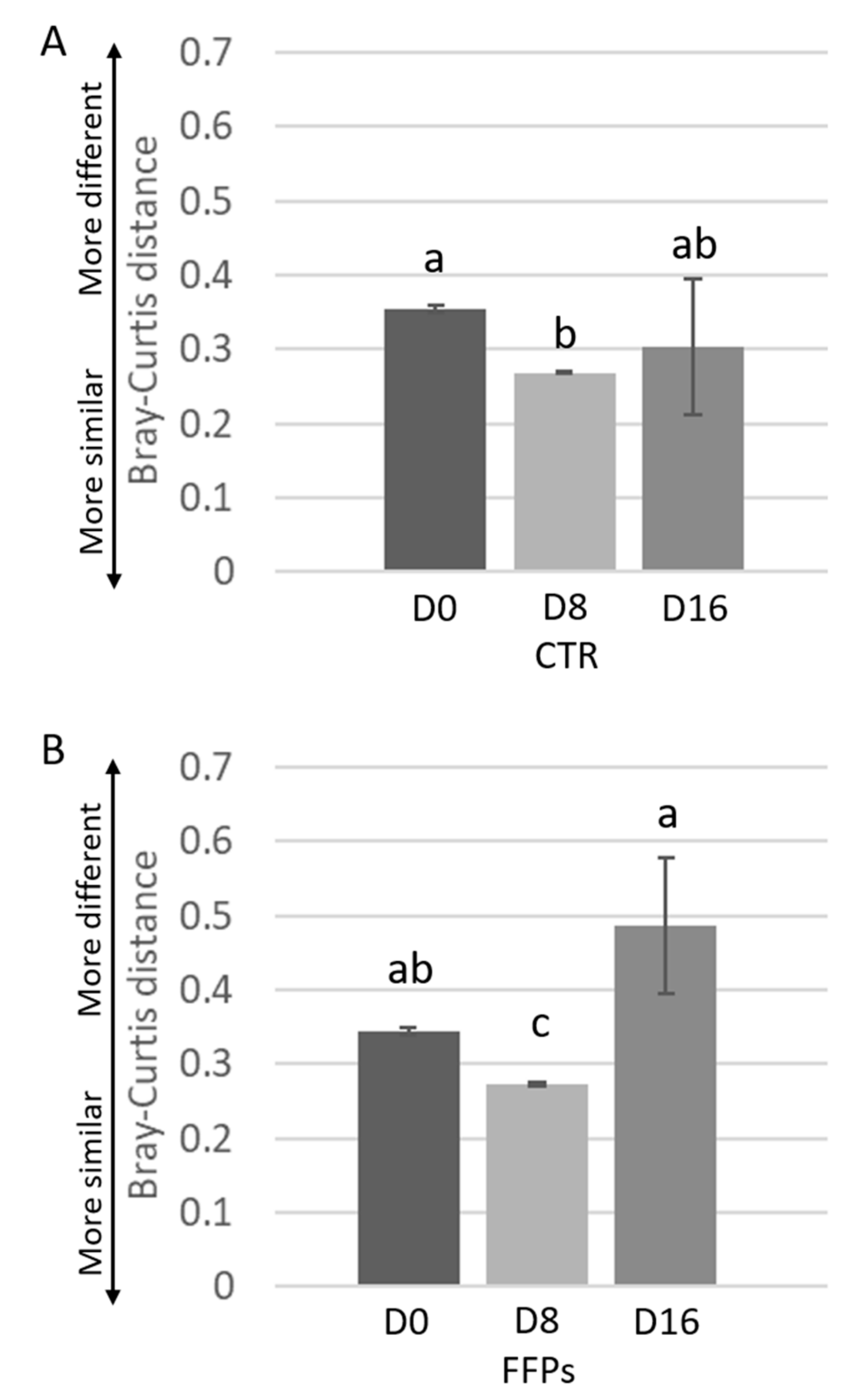
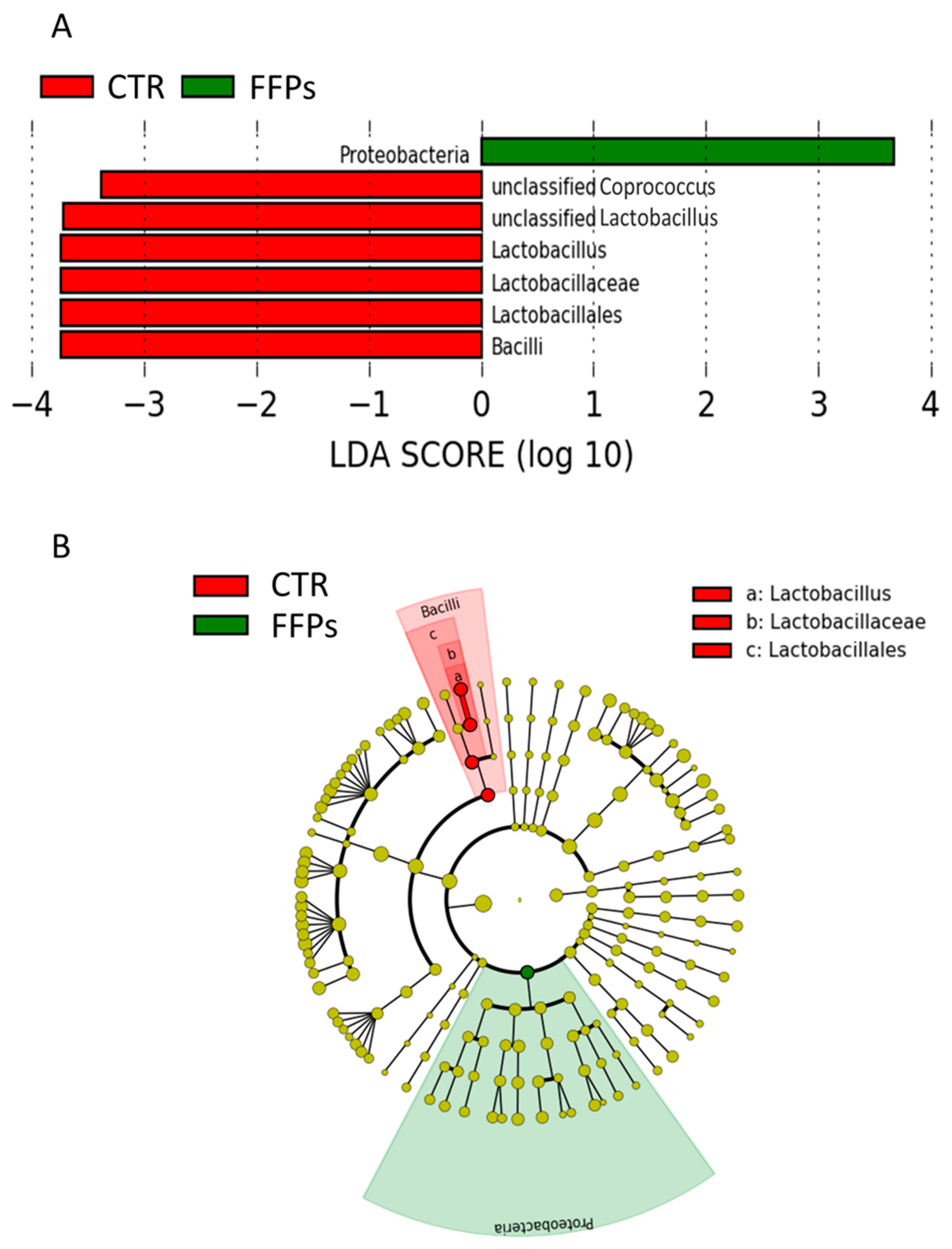
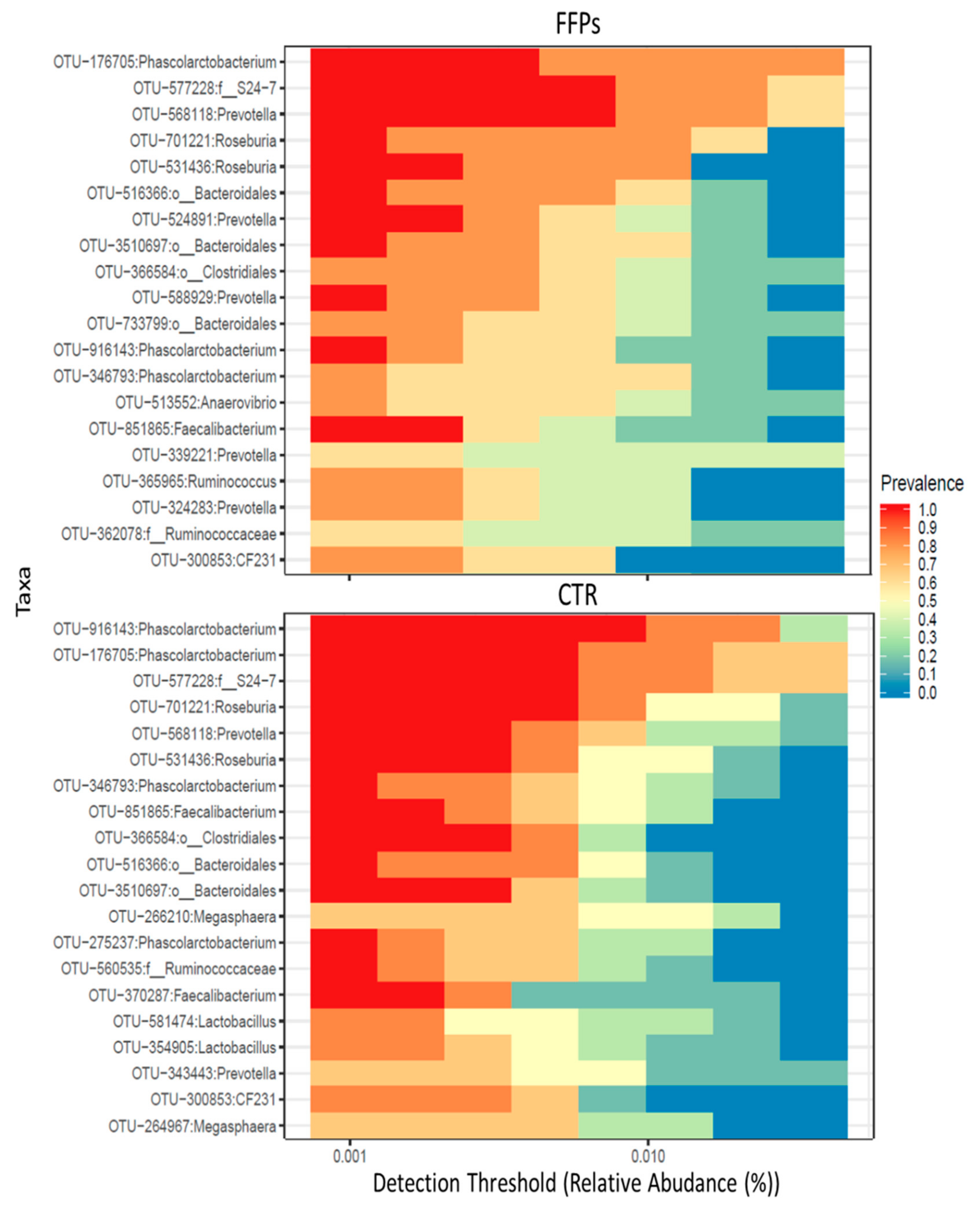
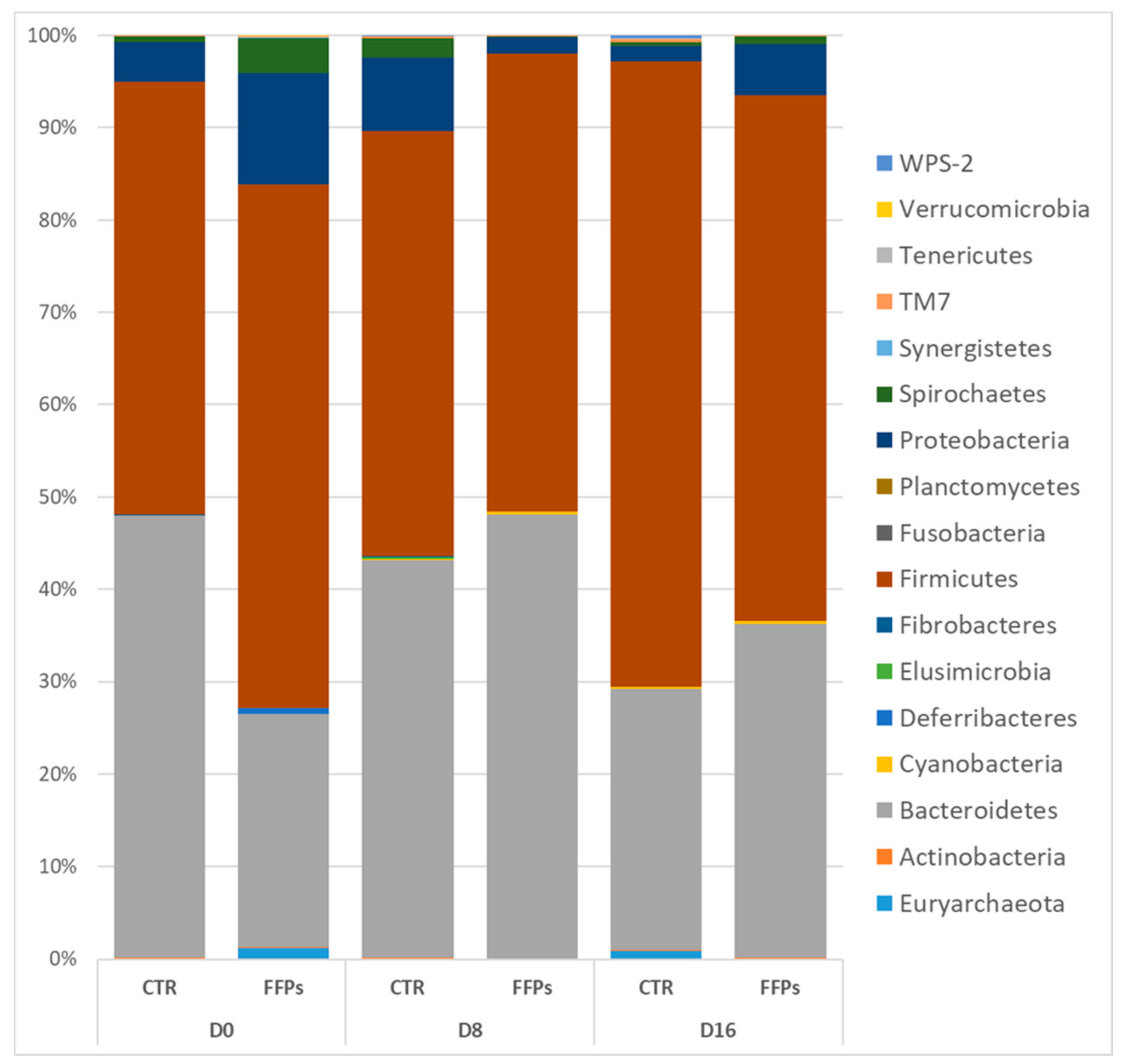
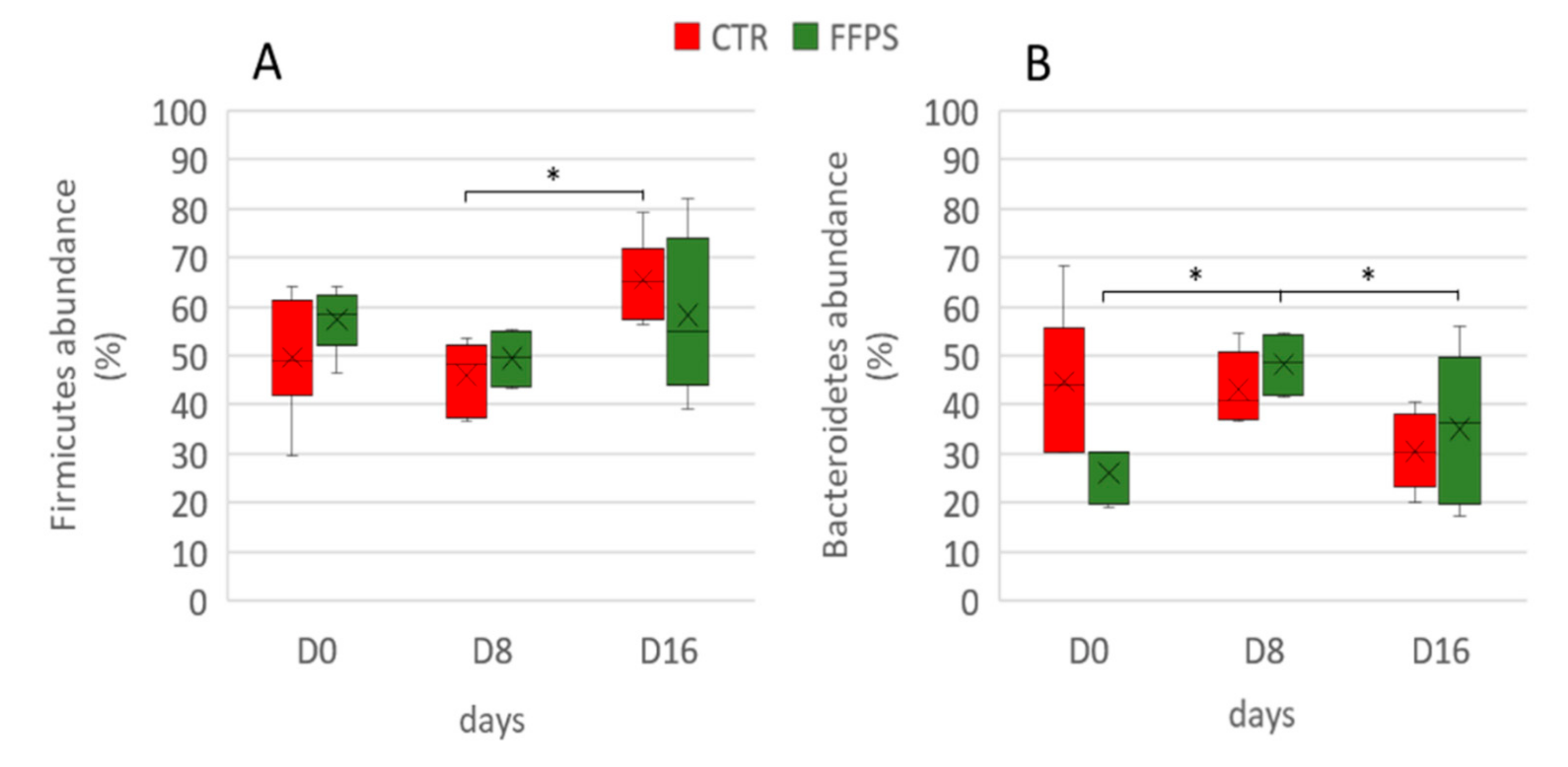
3.2. Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pinotti, L.; Giromini, C.; Ottobon, i M.; Tretola, M.; Marchis, D. Insects and former foodstuffs for upgrading food waste biomasses/streams to feed ingredients for farm animals. Animal 2019, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tretola, M.; Di Rosa, A.; Tirloni, E.; Ottoboni, M.; Giromini, C.; Leone, F.; Pinotti, L. Former food products safety: microbiological quality and computer vision evaluation of packaging remnants contamination. Food Addit. Contam. Part. A 2017, 34, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Tretola, M.; Ottoboni, M.; Di Rosa, A.; Giromini, C.; Fusi, E.; Rebucci, R.; Pinotti, L. Former food products safety evaluation: Computer vision as an innovative approach for the packaging remnants detection. J. Food Qual. 2017, 2017, 6. [Google Scholar] [CrossRef]
- Giromini, C.; Ottoboni, M.; Tretola, M.; Marchis, D.; Gottardo, D.; Caprarulo, V.; Baldi, A.; Pinotti, L. Nutritional evaluation of former food products (ex-food) intended for pig nutrition. Food Addit. Contam. Part. A 2017, 34, 1436–1445. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jha, R.; Stein, H. Nutritional composition, gross energy concentration, and in vitro digestibility of dry matter in 46 sources of bakery meals. J. Anim. Sci. 2018, 96, 4685–4692. [Google Scholar] [CrossRef]
- Ottoboni, M.; Tretola, M.; Luciano, A.; Giuberti, G.; Gallo, A.; Pinotti, L. Carbohydrate digestion and predicted glycemic index of bakery/confectionary ex-food intended for pig nutrition. Ital. J. Anim. Sci. 2019, 18, 838–849. [Google Scholar] [CrossRef]
- Giuberti, G.; Gallo, A.; Masoero, F. In vitro starch digestion and predicted glycemic index of cereal grains commonly utilized in pig nutrition. Anim. Feed Sci. Tech. 2012, 174, 163–173. [Google Scholar] [CrossRef]
- Guo, J.Y.; Phillips, C.E.; Coffey, M.T.; Kim, S.W. Efficacy of a supplemental candy coproduct as an alternative carbohydrate source to lactose on growth performance of newly weaned pigs in a commercial farm condition. J. Anim. Sci. 2015, 93, 5304–5312. [Google Scholar] [CrossRef]
- Prandini, A.; Sigolo, S.; Moschini, M.; Giuberti, G.; Morlacchini, M. Effect of the inclusion of dry pasta by-products at different levels in the diet of typical Italian finishing heavy pigs: Performance, carcass characteristics, and ham quality. Meat Sci. 2016, 114, 38–45. [Google Scholar] [CrossRef]
- Fouhse, J.; Zijlstra, R.; Willing, B. The role of gut microbiota in the health and disease of pigs. Anim. Front. 2016, 6, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Lalles, J.; Bosi, P.; Smidt, H.; Stokes, C. Nutritional management of gut health in pigs around weaning. Proc. Nutr. Soc. 2007, 66, 260–268. [Google Scholar] [CrossRef]
- AOAC Association of Official Analytical Chemists. Official Methods of Analysis, 18th ed.; Scientific Research: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Noblet, J.; Perez, J. Prediction of digestibility of nutrients and energy values of pig diets from chemical analysis. J. Anim. Sci. 1993, 71, 3389–3398. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Swine; National Academy of Sciences: Washington, DC, USA.
- Caporaso, J.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.; Costello, E.; Fierer, N.; Pena, A.; Goodrich, J.; Gordon, J. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods. 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.; Subramanian, S.; Faith, J.; Gevers, D.; Gordon, J.; Knight, R.; Mills, D.; Caporaso, J. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods. 2013, 10, 57–59. [Google Scholar] [CrossRef]
- McDonald, D.; Price, M.n.; Goodrich, J.; Nawrock, E.P.; DeSantis, T.Z.; Probst, A.; Anderson, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Tretola, M.; Ottoboni, M.; Luciano, A.; Rossi, L.; Baldi, A.; Pinotti, L. Former food products have no detrimental effects on diet digestibility, growth performance and selected plasma variables in post-weaning piglets. Ital. J. Anim. Sci. 2019, 18, 987–996. [Google Scholar] [CrossRef]
- Ran, S.; Mu, C.; Zhu, W. Diversity and community pattern of sulfate-reducing bacteria in piglet gut. J. Anim. Sci. Biotechno. 2019, 10, 40. [Google Scholar] [CrossRef]
- Giuberti, G.; Gallo, A.; Masoero, F.; Ferraretto, L.; Hoffman, P.; Shaver, R. Factors affecting starch utilization in large animal food production system: A review. Starch Stärke 2014, 66, 72–90. [Google Scholar] [CrossRef]
- Williams, B.; Mikkelsen, D.; Flanagan, B.; Gidley, M. “Dietary fibre”: moving beyond the "soluble/insoluble" classification for monogastric nutrition, with an emphasis on humans and pigs. J. Anim. Sci. Biotechno. 2019, 10, 45. [Google Scholar] [CrossRef]
- McCann, K. The diversity±stability debate. Nature 2000, 405, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Dillon, R.; Vennard, C.; Buckling, A.; Charnley, A. Diversity of locust gut bacteria protects against pathogen invasion. Ecol. Lett. 2005, 8, 1291–1298. [Google Scholar] [CrossRef]
- Stecher, B.; Chaffron, S.; Käppeli, R.; Hapfelmeier, S.; Freedrich, S.; Weber, T.C.; Kirundi, J.; Suar, M.; McCoy, K.D.; Mering, C.V.; et al. Like will to like: Abundances of closely related species can predict susceptibility to intestinal colonization by pathogenic and commensal Bacteria. PLoS Pathog. 2010, 6, e1000711. [Google Scholar] [CrossRef] [PubMed]
- Werner, J.J.; Garcia, M.L.; Perkins, S.D.; Yarasheski, K.E.; Smith, S.R.; Muegge, B.D.; Stadermann, F.J.; DeRito, C.M.; Floss, C.; Madsen, E.L.; et al. Microbial community dynamics and stability during an ammonia-induced shift to syntrophic acetate oxidation. Appl. Environ. Microbiol. 2014, 8, 3375–3383. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, R.; Kim, H. The intestinal microbiome of the pig. Anim. Health Res. Rev. 2012, 13, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xu, Y.; Chen, X.; Fang, C.; Zhao, L.; Chen, F. The maturing development of gut microbiota in commercial piglets during the weaning transition. Front. Microbiol. 2017, 8, 1688. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Borewicz, K.; White, B.; Singer, R.; Sreevatsan, S.; Tu, Z.; Isaacson, R. Longitudinal investigation of the age-related bacterial diversity in the feces of commercial pigs. Vet. Microb. 2011, 153, 24–133. [Google Scholar] [CrossRef] [PubMed]
- Looft, T.; Johnson, T.; Allen, H.; Bayles, D.; Alt, D.; Stedtfeld, R.; Hashsham, S. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. 2012, 109, 1691–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Nie, Y.; Chen, J.; Zhang, Y.; Wang, Z.; Fan, Q.; Yan, X. Gradual changes of gut microbiota in weaned miniature piglets. Front. Microbiol. 2016, 7, 1727. [Google Scholar] [CrossRef]
- Park, H.; Lee, S.; Kim, H.; Isaacson, R.; Seo, S.; Song, K. Association of obesity with serum leptin, adiponectin, and serotonin and gut microflora in beagle dogs. J. Vet. Intern. Med. 2015, 29, 43–50. [Google Scholar] [CrossRef]
- Vázquez-Baeza, Y.; Hyde, E.; Suchodolski, J.; Knight, R. Dog and human inflammatory bowel disease rely on overlapping yet distinct dysbiosis networks. Nat. Microbiol. 2016, 1, 16177. [Google Scholar] [CrossRef] [PubMed]
- Suchodolski, J.; Foster, M.; Sohail, M.; Leutenegger, C.; Queen, E.; Steiner, J.; Marks, S. The fecal microbiome in cats with diarrhea. PloS ONE 2015, 10, e0127378. [Google Scholar] [CrossRef]
- Harmsen, H.; Elfferich, P.; Schut, F.; Welling, G. A 16S rRNA-targeted probe for detection of lactobacilli and enterococci in faecal samples by fluorescent in situ hybridization. Microb. Ecol. Health Dis. 1999, 11, 3–12. [Google Scholar]
- Azad, M.; Kalam, A.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. Biomed. Res. Int. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Guo, X.; Zhang, J.; Zhang, M.; Ou, Z.; Peng, Y. Phascolarctobacterium faecium abundant colonization in human gastrointestinal tract. Exp. Ther. Med. 2017, 14, 3122–3126. [Google Scholar] [CrossRef] [PubMed]
- Ormerod, K.L.; Wood, D.L.A.; Lachner, N.; Gellatly, S.L.; Daly, J.N.; Parsons, J.D.; Dal’Molin, C.G.O.; Palfreyman, R.W.; Nielsen, L.K.; Cooper, M.A.; et al. Genomic characterization of the uncultured Bacteroidales family S24–7 inhabiting the guts of homeothermic animals. Microbiome 2016, 4, 36. [Google Scholar] [CrossRef]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alpha Diversity Indexes | D 0 | D 8 | D 16 | p-Values 1 | |||||
|---|---|---|---|---|---|---|---|---|---|
| CTR | FFPs | CTR | FFPs | CTR | FFPs | T | G | TxG | |
| Shannon’s | 6.19 ± 0.19 AB | 6.39 ± 0.16 AB | 6.46 ± 0.04 A | 5.96 ± 0.2 B | 6.25 ± 0.10 A | 5.76 ± 0.13 B | 0.57 | 0.001 | 0.71 |
| Chao1 | 549.71 ± 20.1 | 556.7 ± 37.8 | 603.9 ± 16.3 | 581.2 ± 22.3 | 609.3 ± 10.8 | 541.3 ± 22.7 | 0.67 | 0.10 | 0.55 |
| OTUs | 470.6 ± 19.4 ab | 489 ± 35.2 ab | 529.3 ± 14.2 ab | 500.0 ± 25.5 ab | 534.1 ± 12.1 a | 435.1 ± 19.1 b | 0.78 | 0.03 | 0.16 |
| PD-whole tree | 35.3 ± 1.06 | 36.5 ± 1.67 | 38.1 ± 0.93 | 36.4 ± 1.07 | 37.8 ± 0.72 | 33.2 ± 1.06 | 0.70 | 0.10 | 0.28 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tretola, M.; Luciano, A.; Ottoboni, M.; Baldi, A.; Pinotti, L. Influence of Traditional vs Alternative Dietary Carbohydrates Sources on the Large Intestinal Microbiota in Post-Weaning Piglets. Animals 2019, 9, 516. https://doi.org/10.3390/ani9080516
Tretola M, Luciano A, Ottoboni M, Baldi A, Pinotti L. Influence of Traditional vs Alternative Dietary Carbohydrates Sources on the Large Intestinal Microbiota in Post-Weaning Piglets. Animals. 2019; 9(8):516. https://doi.org/10.3390/ani9080516
Chicago/Turabian StyleTretola, Marco, Alice Luciano, Matteo Ottoboni, Antonella Baldi, and Luciano Pinotti. 2019. "Influence of Traditional vs Alternative Dietary Carbohydrates Sources on the Large Intestinal Microbiota in Post-Weaning Piglets" Animals 9, no. 8: 516. https://doi.org/10.3390/ani9080516