

Response of Parasite Community Composition to Aquatic Pollution in Common Carp (Cyprinus carpio L.): A Semi-Experimental Study
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Study Area and Experimental Design
2.2. Fish Processing and Chemical Analysis
2.3. Parasite Collection and Identification
2.4. Data Analysis
3. Results
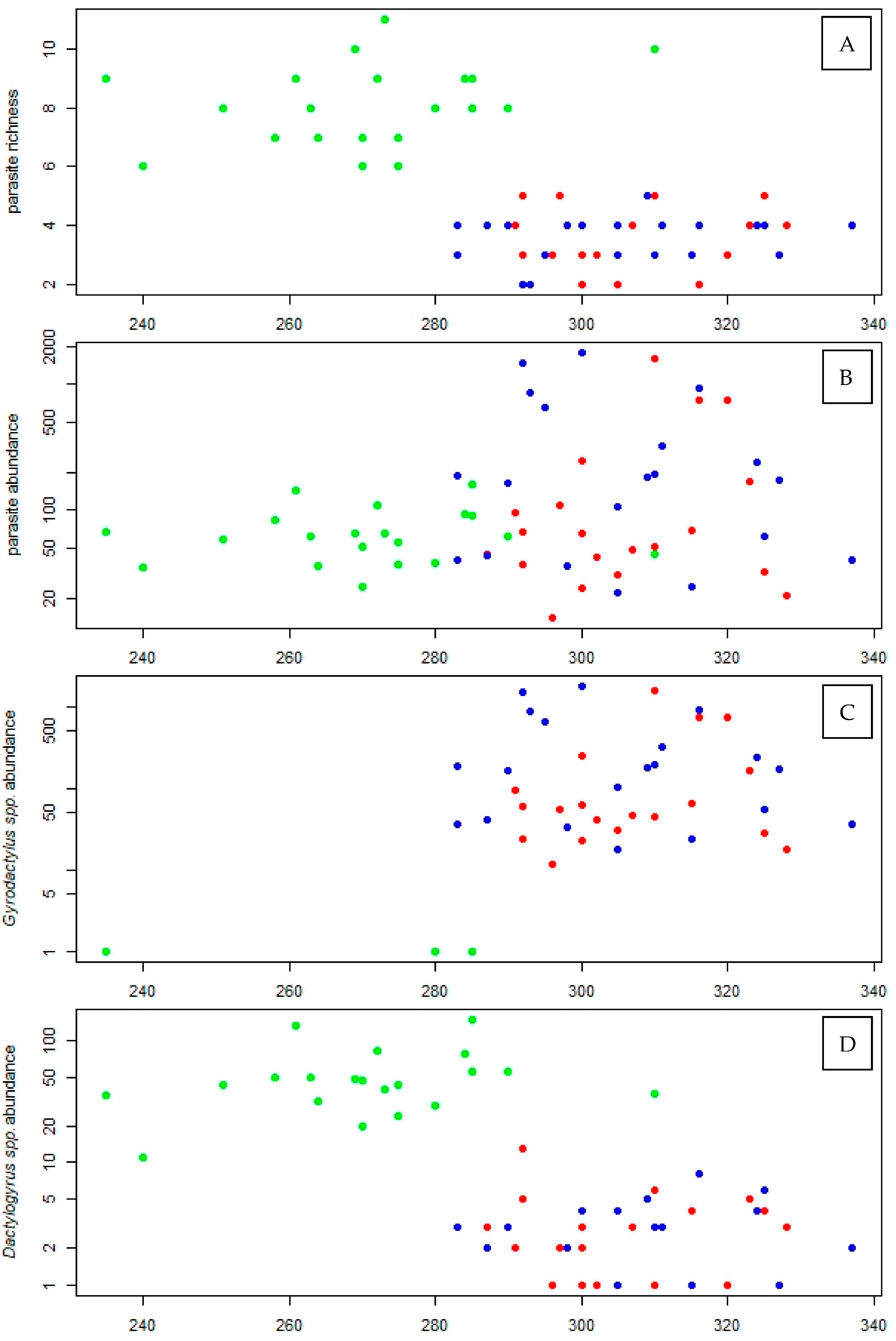
3.1. Parasite Community
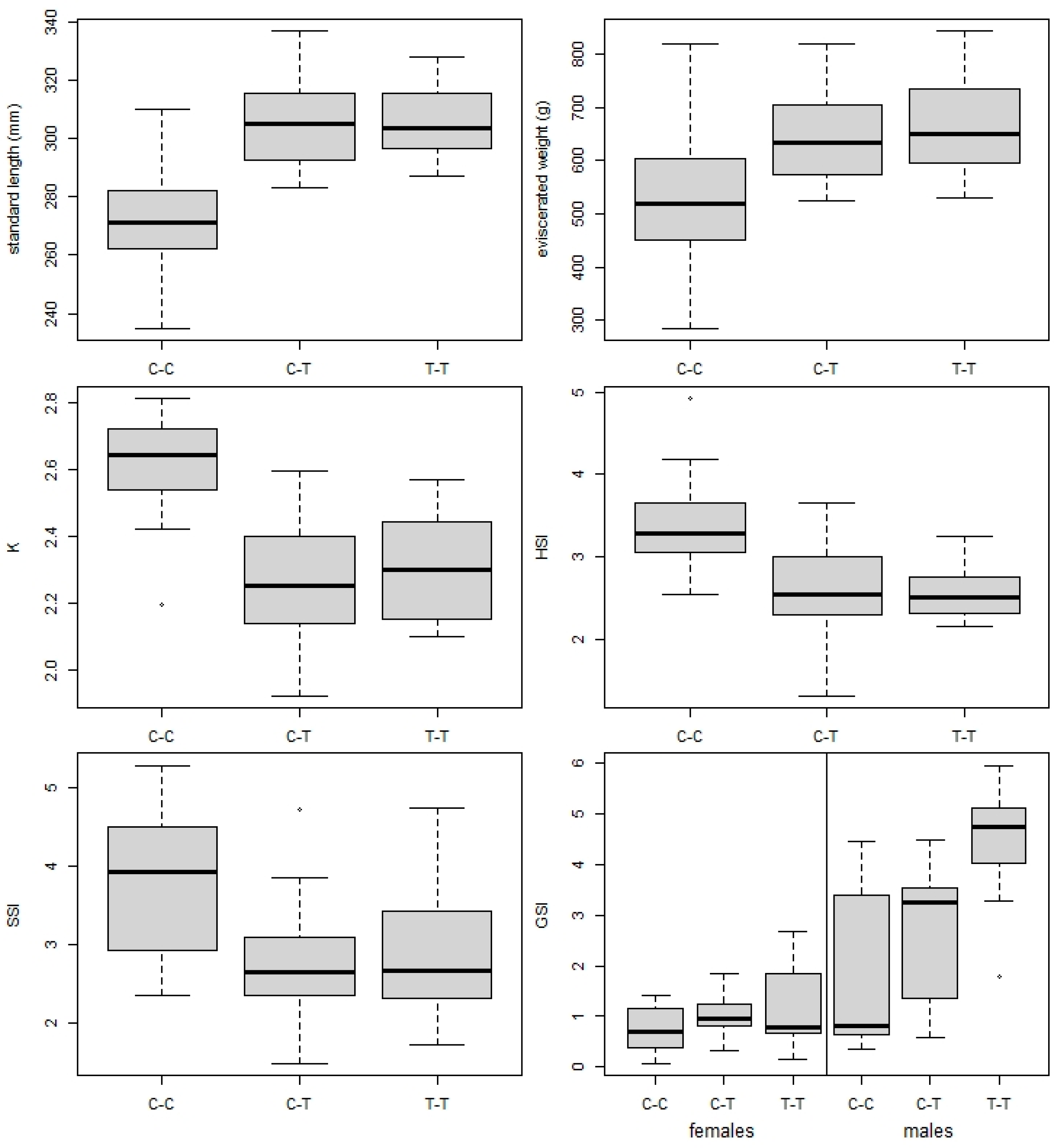
3.2. Fish Biometric Parameters
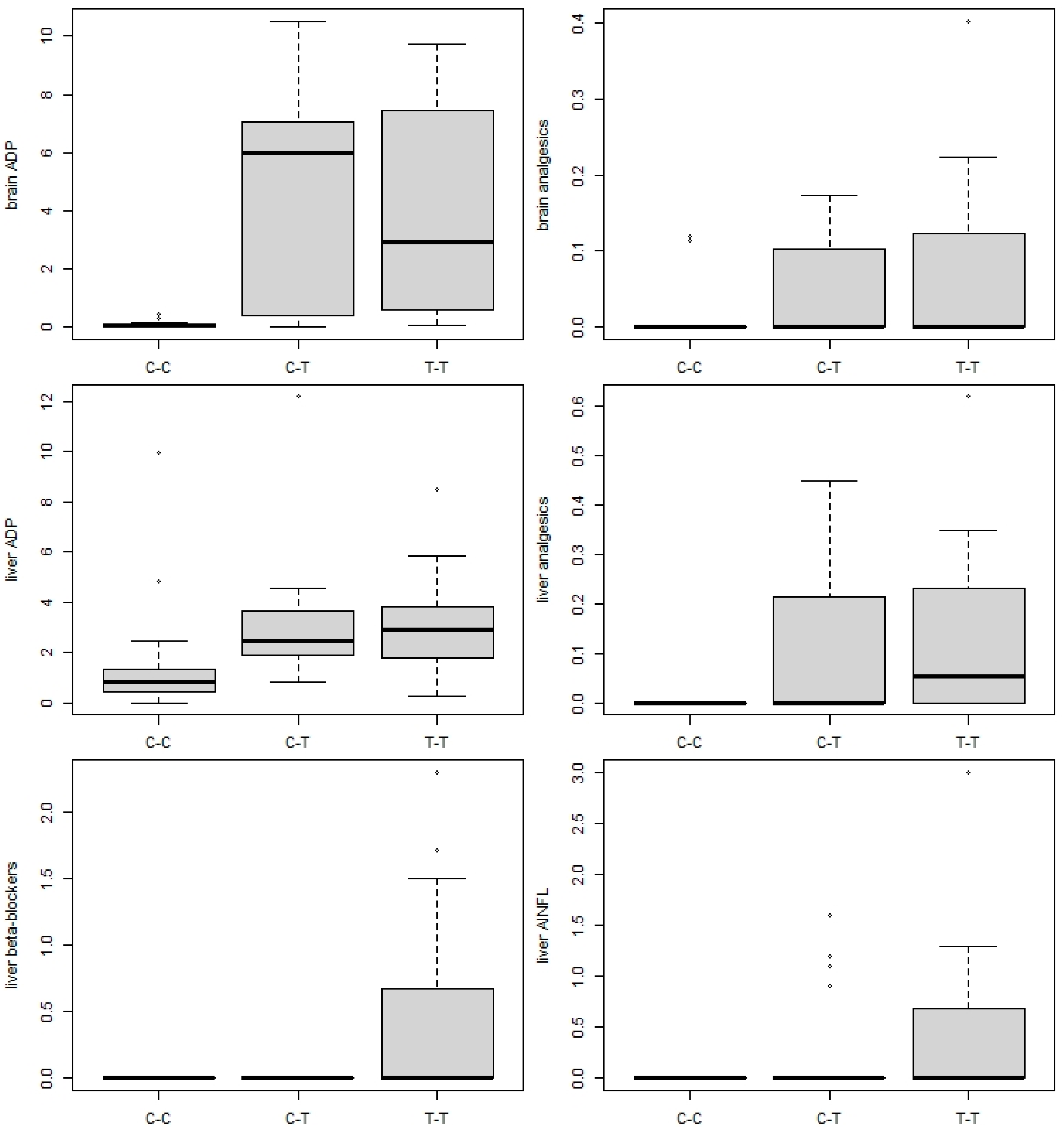
3.3. Presence of Pharmaceuticals in Fish Tissues
3.4. Association between Parasite Infection and Pharmaceutical Load
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carpenter, S.R.; Stanley, E.H.; Vander Zanden, M.J. State of the World’s Freshwater Ecosystems: Physical, Chemical, and Biological Changes. Annu. Rev. Environ. Resour. 2011, 36, 75–99. [Google Scholar] [CrossRef]
- Blettler, M.C.M.; Abrial, E.; Khan, F.R.; Sivri, N.; Espinola, L.A. Freshwater plastic pollution: Recognising research biases and identifying knowledge gaps. Water Res. 2018, 143, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Amoatey, P.; Baawain, M.S. Effects of pollution on freshwater aquatic organisms. Water Environ. Res. 2019, 91, 1272–1287. [Google Scholar] [CrossRef]
- Ebele, A.J.; Abdallah, M.A.; Harrad, S. Pharmaceuticals and personal care products (PPCPs) in the freshwater aquatic environment. Emerg. Contam. 2017, 3, 1–16. [Google Scholar] [CrossRef]
- Fekadu, S.; Alemayehu, E.; Dewil, R.; Van der Bruggen, B. Pharmaceuticals in freshwater aquatic environments: A comparison of the African and European challenge. Sci. Total Environ. 2019, 654, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Khan, M.Z.A.; Shahane, S.; Rai, D.; Chauhan, D.; Kant, C.; Chaudhary, V.K. Emerging Pollutants in Aquatic Environment: Source, Effect, and Challenges in Biomonitoring and Bioremediation—A Review. Pollution 2020, 6, 99–113. [Google Scholar]
- Fent, K.; Weston, A.A.; Caminada, D. Ecotoxicology of human pharmaceuticals. Aquat. Toxicol. 2006, 76, 122–159. [Google Scholar] [CrossRef]
- Morley, N.J. Environmental risk and toxicology of human and veterinary waste pharmaceutical exposure to wild aquatic host-parasite relationships. Environ. Toxicol. Pharmacol. 2009, 27, 161–175. [Google Scholar] [CrossRef]
- Hamilton, W.L.; Doyle, C.; Halliwell-Ewen, M.; Lambert, G. Public health interventions to protect against falsified medicines: A systematic review of international, national and local policies. Health Policy Plan. 2016, 31, 1448–1466. [Google Scholar] [CrossRef]
- Nilsen, E.; Smalling, K.L.; Ahrens, L.; Gros, M.; Miglioranza, K.S.; Pico, Y.; Schoenfuss, H.L. Critical review: Grand challenges in assessing the adverse effects of contaminants of emerging concern on aquatic food webs. Environ. Toxicol. Chem. 2019, 38, 46–60. [Google Scholar] [CrossRef]
- Marcogliese, D.J.; Pietrock, M. Combined effects of parasites and contaminants on animal health: Parasites do matter. Trends Parasitol. 2011, 27, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Sures, B.; Nachev, M.; Selbach, C.; Marcogliese, D.J. Parasite responses to pollution: What we know and where we go in ‘Environmental Parasitology’. Parasit. Vectors 2017, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Marcogliese, D.J. Parasites of the superorganism: Are they indicators of ecosystem health? Int. J. Parasitol. 2005, 35, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Blanar, C.A.; Marcogliese, D.J.; Couillard, C.M. Natural and anthropogenic factors shape metazoan parasite community structure in mummichog (Fundulus heteroclitus) from two estuaries in New Brunswick, Canada. Folia Parasitol. 2011, 58, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Pietrock, M.; Marcogliese, D.J. Free-living endohelminth stages: At the mercy of environmental conditions. Trends Parasitol. 2003, 19, 293–299. [Google Scholar] [CrossRef]
- Hua, J.; Buss, N.; Kim, J.; Orlofske, S.A.; Hoverman, J.T. Population-specific toxicity of six insecticides to the trematode Echinoparyphium sp. Parasitology 2016, 143, 542–550. [Google Scholar] [CrossRef]
- Gilbert, B.M.; Avenant-Oldewage, A. Effects of altered water quality and trace elements on the infection variables of Paradiplozoon ichthyoxanthon (Monogenea: Diplozoidae) from two sites in the Vaal River system, South Africa. Acta Parasitol. 2016, 61, 52–62. [Google Scholar] [CrossRef]
- Möller, H. Pollution and parasitism in the aquatic environment. Int. J. Parasitol. 1987, 17, 353–361. [Google Scholar] [CrossRef]
- Gilbert, B.M.; Avenant-Oldewage, A. Monogeneans as bioindicators: A meta-analysis of effect size of contaminant exposure toward Monogenea (Platyhelminthes). Ecol. Indic. 2021, 130, 108062. [Google Scholar] [CrossRef]
- Marcogliese, D.J. Parasites: Small players with crucial roles in the ecological theatre. EcoHealth 2004, 1, 151–164. [Google Scholar] [CrossRef]
- Blanar, C.A.; Munkittrick, K.R.; Houlahan, J.; MacLatchy, D.L.; Marcogliese, D.J. Pollution and parasitism in aquatic animals: A meta-analysis of effect size. Aquat. Toxicol. 2009, 93, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Koskivaara, M.; Valtonen, E.T.; Prost, M. Seasonal occurrence of gyrodactylid monogeneans on the roach (Rutilus rutilus) and variations between four lakes of differing water quality in Finland. Aqua Fenn. 1991, 21, 47–55. [Google Scholar]
- Brázová, T.; Torres, J.; Eira, C.; Hanzelová, V.; Miklisová, D.; Šalamún, P. Perch and its parasites as heavy metal biomonitors in a freshwater environment: The Case Study of the Ružín Water Reservoir, Slovakia. Sensors 2012, 12, 3068–3081. [Google Scholar] [CrossRef]
- Pravdova, M.; Kolarova, J.; Grabicova, K.; Mikl, L.; Blaha, M.; Randak, T.; Kvach, Y.; Jurajda, P.; Ondrackova, M. Associations between pharmaceutical contaminants, parasite load and health status in brown trout exposed to sewage effluent in a small stream. Ecohydrol. Hydrobiol. 2021, 21, 233–243. [Google Scholar] [CrossRef]
- Pravdova, M.; Kolarova, J.; Grabicova, K.; Randak, T.; Janac, M.; Kvach, Y.; Jurajda, P.; Ondrackova, M. Pharmaceutical contamination and biotic factors affecting parasitism in common carp (Cyprinus carpio). Aquacult. Res. 2022, 53, 4116–4127. [Google Scholar] [CrossRef]
- Giang, P.; Sakalli, S.; Fedorova, G.; Sarvenaz, K.T.; Bakal, T.; Najmanova, L.; Grabicova, K.; Kolarova, J.; Sampels, S.; Zamaratskaia, G.; et al. Biomarker response, health indicators, and intestinal microbiome composition in wild brown trout (Salmo trutta m. fario L.) exposed to a sewage treatment plant effluent-dominated stream. Sci. Total Environ. 2018, 625, 1494–1509. [Google Scholar]
- Grabicova, K.; Grabic, R.; Fedorova, G.; Stanova, A.; Blaha, M.; Randak, T.; Brooks, B.; Zlabek, V. Water reuse and aquaculture: Pharmaceutical bioaccumulation by fish during tertiary treatment in a wastewater stabilization pond. Environ. Pollut. 2020, 267, 115593. [Google Scholar] [CrossRef]
- Fedorova, G.; Grabic, R.; Grabicova, K.; Turek, J.; Van Nguyen, T.; Randak, T.; Brooks, B.W.; Zlabek, V. Water reuse for aquaculture: Comparative removal efficacy and aquatic hazard reduction of pharmaceuticals by a pond treatment system during a one-year study. J. Hazard. Mater. 2022, 421, 126712. [Google Scholar] [CrossRef]
- Kares, M. Do Fish Kept in a Biological Pond Supplied with Water from a WWTP Meet the Requirements for Hygienic Meat Quality? Bachelor’s Thesis, University of South Bohemia, Ceske Budejovice, Czech Republic, 2019. [Google Scholar]
- Rohlenová, K.; Morand, S.; Hyršl, P.; Tolarová, S.; Flajšhans, M.; Šimková, A. Are fish immune systems really affected by parasites? An immunoecological study of common carp (Cyprinus carpio). Parasites Vectors 2011, 4, 120. [Google Scholar] [CrossRef]
- Giang, P.T.; Burkina, V.; Sakalli, S.; Schmidt-Posthaus, H.; Rasmussen, M.K.; Randak, T.; Grabic, R.; Grabicova, K.; Fedorova, G.; Koba, O.; et al. Effects of multi-component mixtures from sewage treatment plant effluent on common carp (Cyprinus carpio) under fully realistic condition. Environ. Manage. 2019, 63, 466–484. [Google Scholar] [CrossRef]
- Koubova, A.; Van Nguyen, T.; Grabicova, K.; Burkina, V.; Aydin, F.G.; Grabic, R.; Novakova, P.; Svecova, H.; Lepic, P.; Fedorova, G.; et al. Metabolome adaptation and oxidative stress response of common carp (Cyprinus carpio) to altered water pollution levels. Environ. Pollut. 2022, 303, 119117. [Google Scholar] [CrossRef] [PubMed]
- Ricker, W.E. Computation and interpretation of biological statistics of fish populations. In Bulletin of the Fisheries Research Board of Canada; Department of the Environment, Fisheries and Marine Service: Ottawa, ON, Canada, 1975; Volume 191, pp. 1–382. [Google Scholar]
- Grabicova, K.; Vojs Stanova, A.; Koba Ucun, O.; Borik, A.; Randak, T.; Grabic, R. Development of a robust extraction procedure for the HPLC-ESI-HRPS determination of multi-residual pharmaceuticals in biota samples. Anal. Chim. Acta 2018, 1022, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, B.; Biserkov, V.; Genov, T. In toto staining method for cestodes with iron acetocarmine. Helminthologia 1986, 23, 279–281. [Google Scholar]
- Georgieva, S.; Soldánová, M.; Pérez-del-Olmo, A.; Dangel, D.R.; Sitko, J.; Sures, B.; Kostadinova, A. Molecular prospecting for European Diplostomum (Digenea: Diplostomidae) reveals cryptic diversity. Int. J. Parasitol. 2013, 43, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Bauer, O.N. Key to the Parasites of Freshwater Fishes of the USSR; Nauka: Leningrad, Russia, 1987; pp. 1–584. (In Russian) [Google Scholar]
- Gusev, A.V.; Dubinina, M.N.; Raikova, E.V.; Khotenkovskiy, I.A.; Pugachev, O.N.; Ergens, R. Metazoan parasites, Part Monogenea. In Key to the Parasites of Freshwater Fishes of the USSR; Bauer, O.N., Ed.; Nauka: Leningrad, Russia, 1985; pp. 1–425. (In Russian) [Google Scholar]
- Moravec, F. Parasitic Nematodes of Freshwater Fishes of Europe, 2nd ed.; Academia Prague: Prague, Czech Republic, 2013; pp. 1–601. [Google Scholar]
- Bush, A.O.; Lafferty, K.; Lotz, M.J.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring biological diversity. Cur. Biol. 2021, 31, R1174–R1177. [Google Scholar] [CrossRef]
- Hammer, R.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. J. Math. Methods Biosci. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Doledec, S.; Chessel, D. Co-inertia analysis: An alternative method for studying species-environment relationships. Freshwater Biol. 1994, 31, 277–294. [Google Scholar] [CrossRef]
- Dray, S.; Chessel, D.; Thioulouse, J. Co-inertia analysis and the linking of ecological data tables. Ecology 2003, 84, 3078–3089. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; ISBN 3-900051-07-0. Available online: http://www.Rproject.org/ (accessed on 22 September 2022).
- Chapman, J.M.; Marcogliese, D.J.; Suski, C.D.; Cooke, S.J. Variation in parasite communities and health indices of juvenile Lepomis gibbosus across a gradient of watershed land-use and habitat quality. Ecol. Indic. 2015, 57, 564–572. [Google Scholar] [CrossRef]
- Marcogliese, D.J.; Gendron, A.D.; Plante, C.; Fournier, M.; Cyr, D. Parasites of spottail shiners (Notropis hudsonius) in the St. Lawrence River: Effects of municipal effluents and habitat. Can. J. Zool. 2006, 84, 1461–1481. [Google Scholar] [CrossRef]
- Nachev, M.; Sures, B. The endohelminth fauna of barbel (Barbus barbus) correlates with water quality of the Danube River in Bulgaria. Parasitology 2009, 136, 545–552. [Google Scholar] [CrossRef]
- Wenger, M.; Ondrackova, M.; Machala, M.; Neca, J.; Hyrsl, P.; Simkova, A.; Jurajda, P.; von der Ohe, P.; Segner, H. Assessing relationships between chemical exposure, parasite infection, fish health, and fish ecological status: A case study using chub (Leuciscus cephalus) in the Bílina River, Czech Republic. Environ. Toxicol. Chem. 2010, 29, 453–466. [Google Scholar] [CrossRef]
- Hanzelová, V.; Oros, M.; Scholz, T. Pollution and diversity of fish parasites: Impact of pollution on the diversity of fish parasites in the Tisa River in Slovakia. In Species Diversity and Extinction; Tepper, G.H., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2011; pp. 265–296. [Google Scholar]
- Morley, N.J.; Lewis, J.W.; Hoole, D. Pollutant-induced effects on immunological and physiological interactions in aquatic host-trematode systems: Implications for parasite transmission. J. Helminthol. 2006, 80, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Austin, B. The effects of pollution on fish health. J. Appl. Microbiol. 1998, 85 (Suppl. 1), 234–242. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, A.; Seth, P.C.; Iqbal, S.A. Trace metal concentration in ambient air at selected sites of Mandideep Township, in vicinity of Bhopal (India). Pollut. Res. 2001, 20, 125–128. [Google Scholar]
- Evering, T.; Weiss, L.M. The immunology of parasite infections in immunocompromised hosts. Parasite Immunol. 2006, 28, 549–565. [Google Scholar] [CrossRef]
- Buchmann, K.; Bresciani, J. Microenvironment of Gyrodactylus derjavini on rainbow trout Oncorhynchus mykiss: Association between mucous cell density in skin and site selection. Parasitol. Res. 1997, 84, 17–24. [Google Scholar] [CrossRef]
- Gheorgiu, C.; Marcogliese, D.J.; Scott, M. Concentration-dependent effects of waterborne zinc on population dynamics of Gyrodactylus turnbulli (Monogenea) on isolated guppies (Poecilia reticulata). Parasitology 2006, 132, 225–232. [Google Scholar] [CrossRef]
- Bakke, T.A.; Harris, P.D.; Jansen, P.A. The susceptibility of Salvelinus fontinalis (Mitchill) to Gyrodactylus salaris Malmberg (Platyhelminthes; Monogenea) under experimental conditions. J. Fish Biol. 1992, 41, 499–507. [Google Scholar] [CrossRef]
- Marcogliese, D.J.; Nagler, J.J.; Cyr, D.G. Effects of exposure to contaminated sediments on the parasite fauna of American plaice (Hippoglossoides platessoides). Bull. Environ. Contamin. Toxicol. 1998, 61, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Madanire-Moyo, G.N.; Luus-Powell, W.J.; Olivier, P.A. Diversity of metazoan parasites of the Mozambique tilapia, Oreochromis mossambicus (Peters, 1852), as indicators of pollution in the Limpopo and Olifants River systems. Onderstepoort J. Vet. Res. 2012, 79, 1–9. [Google Scholar] [CrossRef]
- Shah, H.B.; Yousuf, A.R.; Chishti, M.Z.; Ahmad, F. Seasonal changes in infrapopulations of Diplozoon kashmirensis Kaw, 1950 (Monogenea: Diplozoidae) along a eutrophic gradient. Parasitol. Res. 2013, 112, 3347–3356. [Google Scholar] [CrossRef]
- aus der Beek, T.; Weber, F.A.; Bergmann, A.; Hickmann, S.; Ebert, I.; Hein, A.; Küster, A. Pharmaceuticals in the environment-Global occurrences and perspectives. Environ. Toxicol. Chem. 2016, 35, 823–835. [Google Scholar] [CrossRef]
- Vasquez, M.I.; Lambrianides, A.; Schneider, M.; Kümmerer, K.; Fatta-Kassinos, D. Environmental side effects of pharmaceutical cocktails: What we know and what we should know. J. Hazard. Mater. 2014, 279, 169–189. [Google Scholar] [CrossRef] [PubMed]
- Sueiro, M.C.; Bagnato, E.; Palacios, M.G. Parasite infection and immune and health-state in wild fish exposed to marine pollution. Mar. Pollut. Bull. 2017, 119, 320–324. [Google Scholar] [CrossRef]
- Grabicova, K.; Lindberg, R.H.; Östman, M.; Grabic, R.; Randak, T.; Larsson, D.J. Tissue-specific bioconcentration of antidepressants in fish exposed to effluent from a municipal sewage treatment plant. Sci. Total Environ. 2014, 488, 46–50. [Google Scholar] [CrossRef]
- Dzieweczynski, T.L.; Hebert, O.L. Fluoxetine alters behavioral consistency of aggression and courtship in male Siamese fighting fish, Betta splendens. Physiol. Behav. 2012, 107, 92–97. [Google Scholar] [CrossRef]
- Barry, M.J. Effects of fluoxetine on the swimming and behavioural responses of the Arabian killifish. Ecotoxicology 2013, 22, 425–432. [Google Scholar] [CrossRef]
- Melvin, S.D.; Wilson, S.P. The utility of behavioral studies for aquatic toxicology testing: A meta-analysis. Chemosphere 2013, 93, 2217–2223. [Google Scholar] [CrossRef] [PubMed]
- Brodin, T.; Fick, J.; Jonsson, M.; Klaminder, J. Dilute concentrations of a psychiatric drug alter behavior of fish from natural populations. Science 2013, 339, 814–815. [Google Scholar] [CrossRef] [PubMed]
- Brodin, T.; Piovano, S.; Fick, J.; Klaminder, J.; Heynen, M.; Jonsson, M. Ecological effects of pharmaceuticals in aquatic systems—Impacts through behavioural alterations. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130580. [Google Scholar] [CrossRef] [PubMed]
- Bagge, A.M.; Valtonen, E.T. Experimental study on the influence of paper and pulp mill effluent on the gill parasite communities of roach (Rutilus rutilus). Parasitology 1996, 112, 499–508. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T-T | C-C | C-T | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Parasite Taxa | p (%) | A | Range | p (%) | A | Range | p (%) | A | Range |
| Ciliophora | |||||||||
| Ichthiophthirius multifiliis | 15 | 0.20 | (1–2) | ||||||
| Trichodina spp. | 80 | 100 | 95 | ||||||
| Monogenea | |||||||||
| Gyrodactylus sprostonae | 100 | 207.75 | (12–1594) | 15 | 0.15 | (1) | 100 | 374.95 | (18–1778) |
| Dactylogyrus falciformis | 10 | 0.15 | (1–2) | 100 | 34.45 | (2–105) | 15 | 0.15 | (1) |
| Dactylogyrus achmerowi | 75 | 1.60 | (1–8) | 95 | 10.05 | (2–31) | 65 | 1.50 | (1–4) |
| Dactylogyrus extensus | 65 | 1.25 | (1–5) | 70 | 3.00 | (1–10) | 55 | 0.95 | (1–4) |
| Dactylogyrus molnari | 70 | 5.20 | (2–24) | ||||||
| Eudiplozoon nipponicum | 5 | 0.05 | (1) | 95 | 3.25 | (1–7) | 5 | 0.05 | (1) |
| Trematoda | |||||||||
| Diplostomum pseudospathaceum | 5 | 0.05 | (1) | 5 | 0.05 | (1) | |||
| Cestoda | |||||||||
| Atractolytocestus huronensis | 5 | 2.55 | (51) | 20 | 1.95 | (1–25) | 5 | 0.10 | (2) |
| Khawia sinensis | 5 | 0.10 | (2) | ||||||
| Valipora campylancristrota | 5 | 0.05 | (1) | 15 | 0.15 | (1) | |||
| Crustacea | |||||||||
| Argulus coregoni | 60 | 0.7 | (1–3) | ||||||
| Argulus foliaceus | 45 | 0.6 | (1–3) | ||||||
| Argulus japonicus | 10 | 0.1 | (1) | ||||||
| Ergasilus sieboldi | 5 | 0.05 | (1) | 90 | 9.45 | (1–29) | 5 | 0.05 | (1) |
| Bivalvia | |||||||||
| Anodonta sp. | 5 | 0.05 | (1) | ||||||
| Hirudinea | |||||||||
| Piscicola geometra | 5 | 0.05 | (1) | ||||||
| T-T | C-T | C-C | |
|---|---|---|---|
| Parasite species richness | 10 | 10 | 16 |
| Overall parasite abundance | 4302 | 7600 | 1439 |
| Dominance index | 0.933 | 0.974 | 0.277 |
| Shannon–Wiener index | 0.202 | 0.090 | 1.721 |
| Equitability | 0.088 | 0.039 | 0.621 |
| Df | SS | MS | F | R2 | p | |
|---|---|---|---|---|---|---|
| C-T vs. T-T | 1 | 0.089 | 0.089 | 2.92 | 0.071 | 0.058 |
| Residuals | 38 | 1.160 | 0.031 | 0.929 | ||
| Total | 39 | 1.249 | 1 | |||
| C-C vs. C-T | 1 | 3.855 | 3.856 | 172.57 | 0.820 | 0.001 |
| Residuals | 38 | 0.849 | 0.022 | 0.180 | ||
| Total | 39 | 4.704 | 1 | |||
| C-C vs. T-T | 1 | 3.740 | 3.740 | 156.35 | 0.804 | 0.001 |
| Residuals | 38 | 0.909 | 0.024 | 0.196 | ||
| Total | 39 | 4.648 | 1 |
| T-T | C-C | C-T | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | Mean ± S | n | Range | Mean ± SD | n | Range | Mean ± SD | n | Range |
| SL (mm) | 306 ± 12 | 20 | (287–328) | 271 ± 17 | 20 | (235–310) | 305 ± 15 | 20 | (245–359) |
| We (g) | 663 ± 90 | 20 | (530–845) | 528 ± 116 | 20 | (285–820) | 646 ± 82 | 20 | (323–820) |
| K | 2.31 ± 0.15 | 20 | (2.1–2.6) | 2.62 ± 0.14 | 20 | (2.2–2.8) | 2.27 ± 0.18 | 20 | (1.9–2.6) |
| HSI | 2.56 ± 0.29 | 20 | (2.2–3.2) | 3.39 ± 0.58 | 20 | (2.5–4.9) | 2.62 ± 0.52 | 20 | (1.3–3.7) |
| SSI | 2.87 ± 0.82 | 20 | (1.7–4.7) | 3.85 ± 0.88 | 20 | (2.4–5.3) | 2.77 ± 0.76 | 20 | (1.5–4.7) |
| GSI (m) | 4.38 ± 1.88 | 9 | (1.8–6.0) | 1.87 ± 1.14 | 9 | (0.3–4.5) | 2.75 ± 1.31 | 10 | (0.6–4.5) |
| GSI (f) | 1.22 ± 1.84 | 11 | (0.1–2.7) | 0.73 ± 1.28 | 10 | (0.1–1.4) | 1.00 ± 1.32 | 9 | (0.3–1.8) |
| Df | SS | MS | F | R2 | p | |
|---|---|---|---|---|---|---|
| C-T vs. T-T | 1 | 12.190 | 12.187 | 1.485 | 0.038 | 0.179 |
| Residuals | 38 | 311.940 | 8.209 | 0.962 | ||
| Total | 39 | 324.140 | 1 | |||
| C-C vs. C-T | 1 | 59.663 | 59.663 | 10.228 | 0.211 | 0.001 |
| Residuals | 38 | 221.671 | 5.833 | 0.788 | ||
| Total | 39 | 281.334 | 1 | |||
| C-C vs. T-T | 1 | 44.089 | 44.089 | 12.158 | 0.242 | 0.001 |
| Residuals | 38 | 137.799 | 3.626 | 0.758 | ||
| Total | 39 | 181.888 | 1 |
| Parasites vs. Pharmaceuticals | Parasites vs. Biometry | Pharmaceuticals vs. Biometry | ||||
|---|---|---|---|---|---|---|
| Group | RV | p | RV | p | RV | p |
| T-T | 0.334 | 0.037 | 0.151 | 0.597 | 0.112 | 0.741 |
| C-T | 0.380 | 0.036 | 0.185 | 0.526 | 0.196 | 0.560 |
| C-C | 0.278 | 0.135 | 0.204 | 0.317 | 0.110 | 0.711 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pravdová, M.; Kolářová, J.; Grabicová, K.; Janáč, M.; Randák, T.; Ondračková, M. Response of Parasite Community Composition to Aquatic Pollution in Common Carp (Cyprinus carpio L.): A Semi-Experimental Study. Animals 2023, 13, 1464. https://doi.org/10.3390/ani13091464
Pravdová M, Kolářová J, Grabicová K, Janáč M, Randák T, Ondračková M. Response of Parasite Community Composition to Aquatic Pollution in Common Carp (Cyprinus carpio L.): A Semi-Experimental Study. Animals. 2023; 13(9):1464. https://doi.org/10.3390/ani13091464
Chicago/Turabian StylePravdová, Markéta, Jitka Kolářová, Kateřina Grabicová, Michal Janáč, Tomáš Randák, and Markéta Ondračková. 2023. "Response of Parasite Community Composition to Aquatic Pollution in Common Carp (Cyprinus carpio L.): A Semi-Experimental Study" Animals 13, no. 9: 1464. https://doi.org/10.3390/ani13091464