
Exploring Efflux as a Mechanism of Reduced Susceptibility towards Biocides and Fluoroquinolones in Staphylococcus pseudintermedius

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Antibiotic, Biocides and Efflux Inhibitors
2.3. Determination of Minimum Inhibitory Concentrations
2.4. Determination of Cut-Off (COWT) Values
2.5. Detection of Efflux Activity by Real Time Fluorometry
2.6. Detection of Plasmid-Encoded Efflux Pump Genes
2.7. Statistical Analysis
3. Results
3.1. Relation between Reduced Susceptibility to TPP and EtBr and Efflux Activity
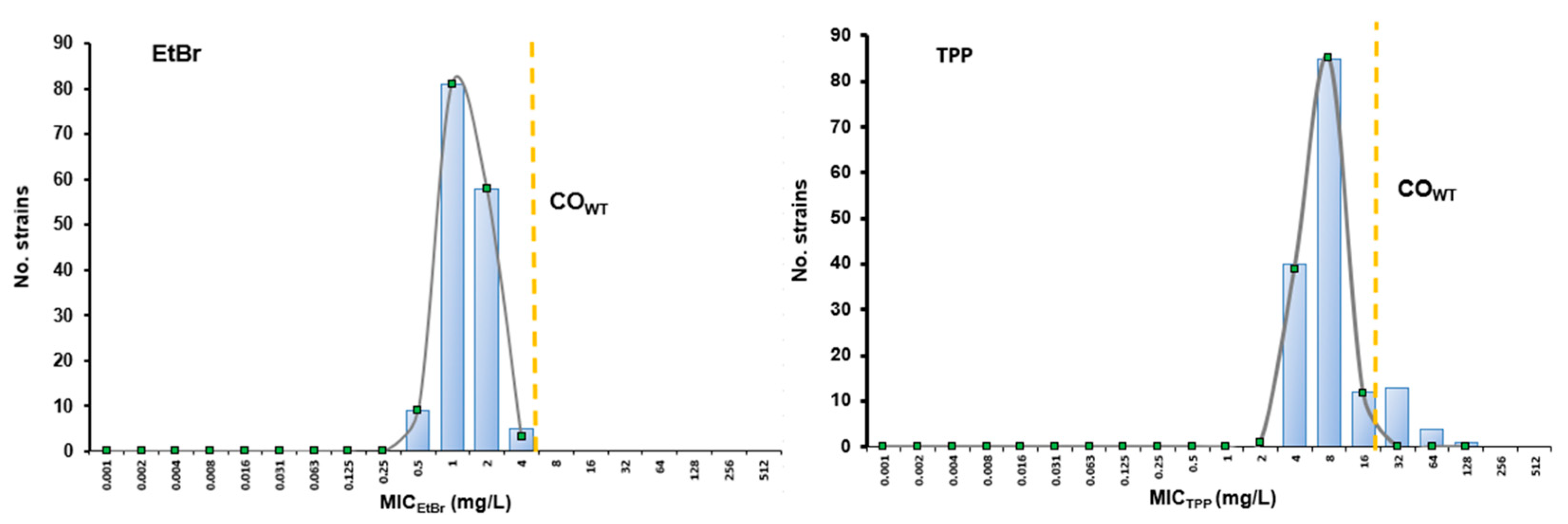
3.1.1. Susceptibility Profiling towards EtBr and TPP
3.1.2. Cut-Off Values (COWT) and Identification of S. pseudintermedius NWT Populations for EtBr and TPP
3.1.3. Assessment of Efflux Activity in S. pseudintermedius Strains
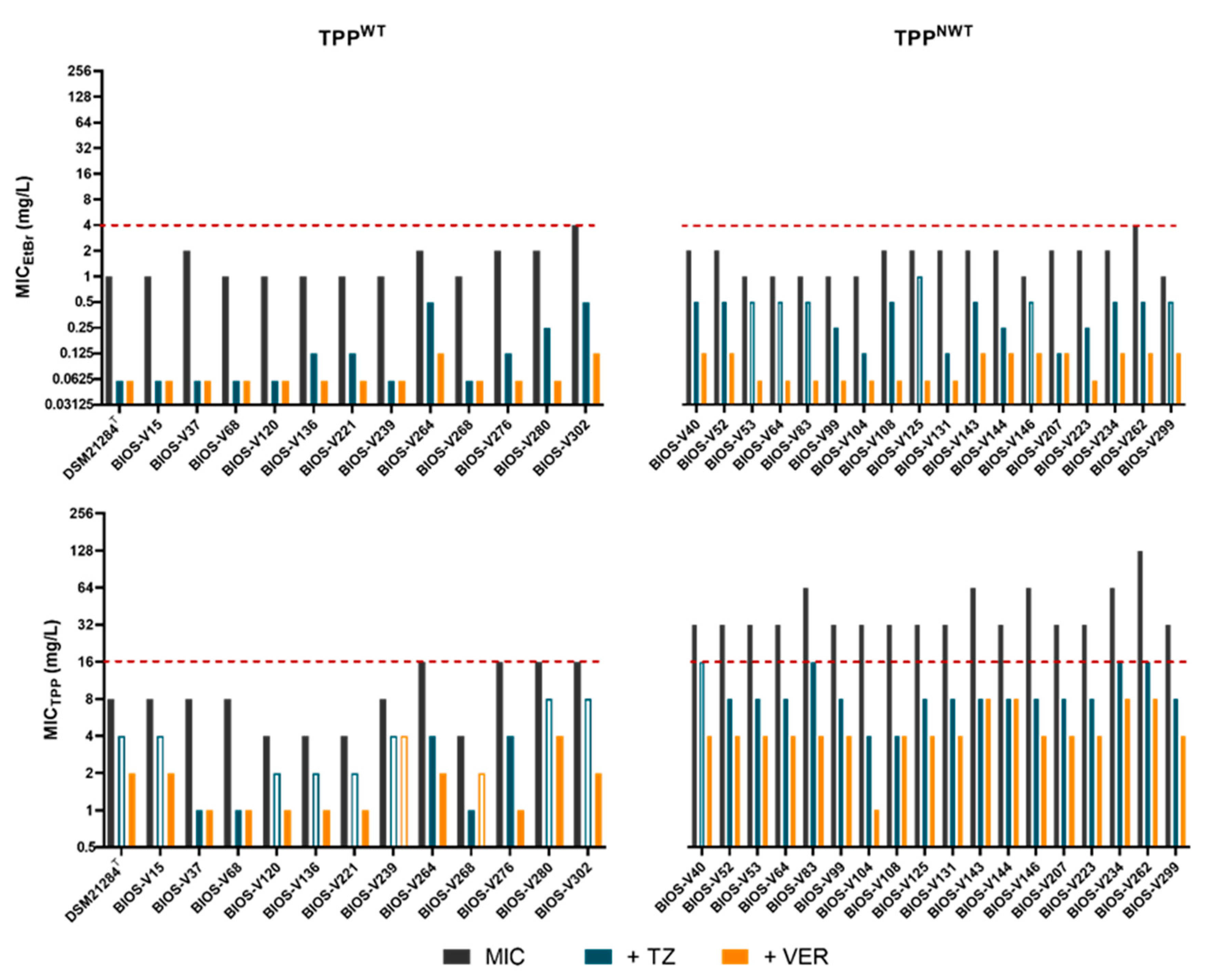
Effect of Efflux Inhibitors (EIs) on MICEtBr and MICTPP
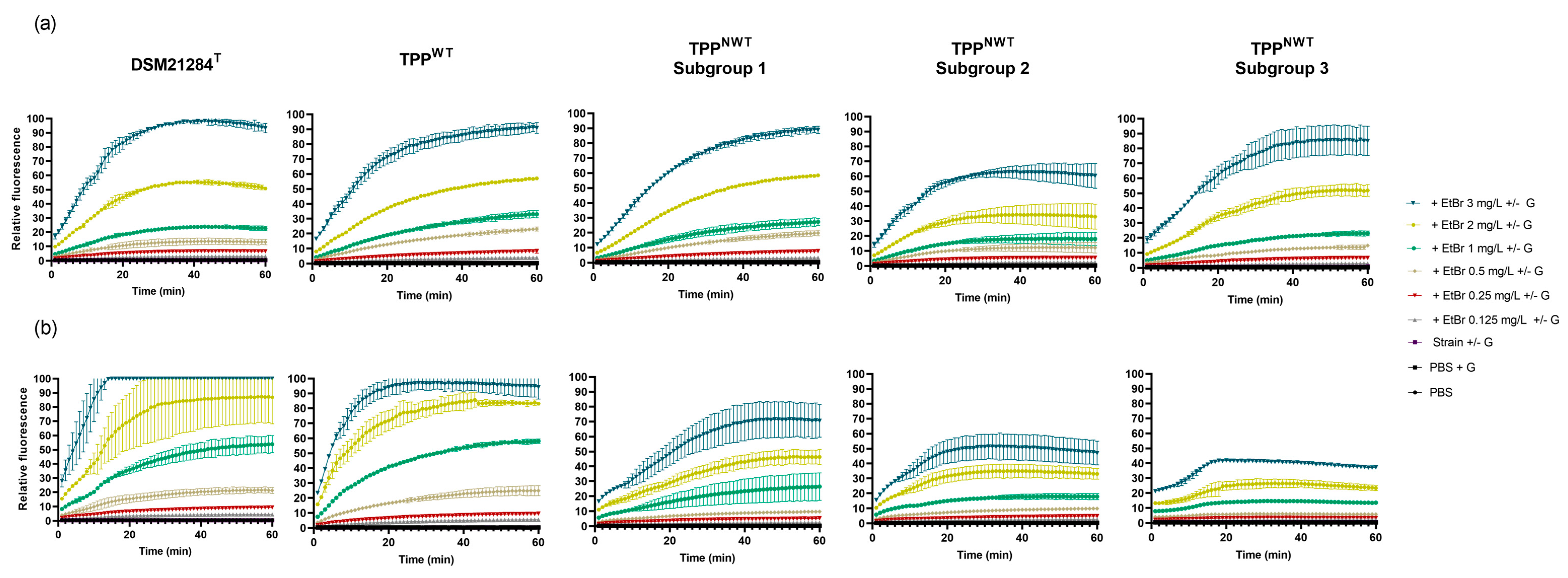
Detection and Analysis of Efflux Activity by Real-Time Fluorometry
3.1.4. Screening of Plasmid-Encoded Efflux Pump Genes qacA/B and smr
3.2. Relation between Resistance to Fluoroquinolones and Efflux Activity
Effect of Efflux Inhibitors on Ciprofloxacin (CIP) Susceptibility Levels
4. Discussion
4.1. Contribution of Efflux to Reduced Susceptibility to Biocides, EtBr and Resistance to Fluoroquinolones in S. pseudintermedius
4.2. Misuse of TPP May Promote Emergence of Efflux-Mediated Resistance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhooshan, S.; Negi, V.; Khatri, P. Staphylococcus pseudintermedius: An undocumented, emerging pathogen in humans. GMS Hyg. Infect. Control. 2020, 15, 1–11. [Google Scholar] [CrossRef]
- Ruiz-Ripa, L.; Simón, C.; Ceballos, S.; Ortega, C. S. pseudintermedius and S. aureus lineages with transmission ability circulate as causative agents of infections in pets for years. BMC Vet. Res. 2021, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.S.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Rojas, J.L.G.; Gortázar, C.; Herskin, M.; Michel, V.; et al. Assessment of listing and categorization of animal diseases within the framework of the Animal Health Law (Regulation (EU) No 2016/429): Antimicrobial-resistant Staphylococcus pseudintermedius in dogs and cats. EFSA J. Eur. Food Saf. Auth. 2022, 20, e07310. [Google Scholar] [CrossRef]
- Lynch, S.A.; Helbig, K.J. The complex diseases of Staphylococcus pseudintermedius in canines: Where to next? Vet. Sci. 2021, 8, 11. [Google Scholar] [CrossRef]
- Frank, L.; Loeffler, A. Meticillin-resistant Staphylococcus pseudintermedius: Clinical challenge and treatment options. Vet. Dermatol. 2012, 23, 283–291. [Google Scholar] [CrossRef]
- Hillier, A.; Lloyd, D.H.; Weese, J.; Blondeau, J.; Boothe, D.; Breitschwerdt, E.; Guardabassi, L.; Papich, M.J.; Rankin, S.; Turnidge, J.T.; et al. Guidelines for the diagnosis and antimicrobial therapy of canine superficial bacterial folliculitis (Antimicrobial Guidelines Working Group of the International Society for Companion Animal Infectious Diseases). Vet. Dermatol. 2014, 25, 163-e43. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.O.; Loeffler, A.; Davis, M.F.; Guardabassi, L.; Weese, J.S. Recommendations for approaches to meticillin-resistant staphylococcal infections of small animals: Diagnosis, therapeutic considerations and preventative measures. Vet. Dermatol. 2017, 28, 304-e69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, K.M.; Waisglass, S.E.; Dick, H.L.N.; Weese, J.S. Prevalence of meticillin-resistant Staphylococcus pseudintermedius (MRSP) from skin and carriage sites of dogs after treatment of their meticillin-resistant or meticillin-sensitive staphylococcal pyoderma. Vet. Dermatol. 2012, 23, 369–375. [Google Scholar] [CrossRef]
- Bierowiec, K.; Miszczak, M.; Korzeniowska-Kowal, A.; Wzorek, A.; Płókarz, D.; Gamian, A. Epidemiology of Staphylococcus pseudintermedius in cats in Poland. Sci. Rep. 2021, 11, 18898. [Google Scholar] [CrossRef]
- Pomba, C.; Rantala, M.; Greko, C.; Baptiste, K.E.; Catry, B.; van Duijkeren, E.; Mateus, A.; Moreno, M.A.; Pyorala, S.; Ruzauskas, M.; et al. Public health risk of antimicrobial resistance transfer from companion animals. J. Antimicrob. Chemother. 2017, 72, 957–968. [Google Scholar] [CrossRef]
- Loeffler, A.; Lloyd, D.H. What has changed in canine pyoderma? A narrative review. Vet. J. 2018, 235, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feßler, A.; Scholtzek, A.; Schug, A.; Kohn, B.; Weingart, C.; Schink, A.; Bethe, A.; Lübke-Becker, A.; Schwarz, S. Antimicrobial and biocide resistance among feline and canine Staphylococcus aureus and Staphylococcus pseudintermedius isolates from diagnostic submissions. Antibiotics 2022, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Gagetti, P.; Rosato, R.R.; Rosato, A.E. Staphylococcus pseudintermedius’s PBP4 is directly associated with the dissociated oxacillin and cefoxitin phenotype. Antibiotics 2021, 10, 1299. [Google Scholar] [CrossRef] [PubMed]
- Borjesson, S.; Gómez-Sanz, E.; Ekstrom, K.; Torres, C.; Gronlund, U. Staphylococcus pseudintermedius can be misdiagnosed as Staphylococcus aureus in humans with dog bite wounds. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Somayajin, R.; Rubin, J.E.; Priyantha, M.; Church, D. Exploring Staphylococcus pseudintermedius: An emerging zoonotic pathogen? Future Microbiol. 2016, 11, 1371–1374. [Google Scholar] [CrossRef] [Green Version]
- Frosini, S.M.; Bond, R.; McCarthy, A.J.; Feudi, C.; Schwarz, S.; Lindsay, J.A.; Loeffler, A. Genes on the move: In vitro transduction of antimicrobial resistance genes between human and canine Staphylococcal pathogens. Microorganisms 2020, 8, 2031. [Google Scholar] [CrossRef]
- Costa, S.S.; Falcão, C.; Viveiros, M.; Machado, D.; Martins, M.; Melo-Cristino, J.; Amaral, L.; Couto, I. Exploring the contribution of efflux on the resistance to fluoroquinolones in clinical isolates of Staphylococcus aureus. BMC Microbiol. 2011, 11, 241. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.S.; Viveiros, M.; Amaral, L.; Couto, I. Multidrug Efflux Pumps in Staphylococcus aureus: An update. Open Microbiol. J. 2013, 7, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.S.; Junqueira, E.; Palma, C.; Viveiros, M.; Melo-Cristino, J.; Amaral, L.; Couto, I. Resistance to antimicrobials mediated by efflux pumps in Staphylococcus aureus. Antibiotics 2013, 2, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.S.; Viveiros, M.; Rosato, A.; Melo-Cristino, J.; Couto, I. Impact of efflux in the development of multidrug resistance phenotypes in Staphylococcus aureus. BMC Microbiol. 2015, 15, 232. [Google Scholar] [CrossRef] [Green Version]
- Blair, J.M.A.; Piddock, L.J.V. How to measure export via bacterial multidrug resistance efflux pumps. ASM 2016, 7, e00840-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeMarco, C.E.; Cushing, L.A.; Frempong-Manso, E.; Seo, S.M.; Jaravaza, T.A.A.; Kaatz, G.W. Efflux-related resistance to norfloxacin, dyes, and biocides in bloodstream isolates of Staphylococcus aureus. Antimicrob. Agents Chemother. 2007, 51, 3235–3239. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.S.; Viveiros, M.; Pomba, C.; Couto, I. Active antimicrobial efflux in Staphylococcus epidermidis: Building up of resistance to fluoroquinolones and biocides in a major opportunistic pathogen. J. Antimicrob. Chemother. 2018, 73, 320–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rampacci, E.; Felicetii, T.; Pietrella, D.; Sabatini, S.; Passamonti, F. Drug efflux transporters in Staphylococcus pseudintermedius: In silico prediction and characterization of resistance. JAC 2022, 77, 3283–3290. [Google Scholar] [CrossRef]
- Couto, N.; Belas, A.; Oliveira, M.; Almeida, P.; Clemente, C.; Pomba, C. Comparative RNA-seq-based transcriptome analysis of the virulence characteristics of methicillin-resistant and -susceptible Staphylococcus pseudintermedius strains isolated from small animals. Antimicrob. Agents. Chemother. 2016, 60, 962–967. [Google Scholar] [CrossRef] [Green Version]
- Worthing, K.A.; Marcus, A.; Abraham, S.; Trott, D.J.; Norris, J.M. Qac genes and biocide tolerance in clinical veterinary methicillin-resistant and methicillin-susceptible Staphylococcus aureus and Staphylococcus pseudintermedius. Vet. Microbiol. 2018, 216, 153–158. [Google Scholar] [CrossRef]
- Morais, C.; Costa, S.S.; Leal, M.; Ramos, B.; Andrade, M.; Ferreira, C.; Abrantes, P.; Pomba, C.; Couto, I. Genetic diversity and antimicrobial resistance profiles of Staphylococcus pseudintermedius associated to skin and soft-tissue infections in companion animals in Lisbon, Portugal. Front. Microbiol. 2023; in press. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing. CLSI Supplement M100, 32nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022. [Google Scholar]
- Clinical and Laboratory Standards Institute (VET01S CLSI). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals. CLSI Supplement VET01S, 5th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Couto, I.; Costa, S.S.; Viveiros, M.; Martins, M.; Amaral, L. Efflux-mediated response of Staphylococcus aureus exposed to ethidium bromide. J. Antimicrob. Chemother. 2008, 62, 504–513. [Google Scholar] [CrossRef] [Green Version]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Media Preparation for Disk Diffusion Testing and for Determination of MIC Values by the Broth Microdilution Method, Version 7.0; European Committee on Antimicrobial Susceptibility Testing (EUCAST): Växjö, Sweden, 2022. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Standard Operating Procedure. MIC Distributions and the Setting of Epidemiological Cutoff (ECOFF) Values, EUCAST SOP 10.2 2021. Available online: http://www.eucast.org (accessed on 15 February 2023).
- Turnidge, J.; Kahlmeter, G.; Kronvall, G. Statistical characterisation of bacterial wildtype MIC value distributions and the determination of epidemiological cut-off values. Clin. Microbiol. Infect. 2006, 12, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Viveiros, M.; Martins, M.; Couto, I.; Rodrigues, L.; Spengler, G.; Martins, A. New methods for the identification of efflux mediated MDR bacteria, genetic assessment of regulators and efflux pump constituents, characterization of efflux systems and screening for inhibitors of efflux pumps. Curr. Drugs Targets 2008, 9, 760–778. [Google Scholar] [CrossRef]
- Costa, S.S.; Lopes, E.; Azzali, E.; Machado, D.; Coelho, T.; Silva, P.; Viveiros, M.; Pieroni, M.; Couto, I. An experimental model for the rapid screening of compounds with potential use against Mycobacteria. Assay Drug Dev. Technol. 2016, 14, 524–534. [Google Scholar] [CrossRef]
- Costa, S.S.; Palma, C.; Kadlec, K.; Fessler, A.T.; Viveiros, M.; Melo-Cristino, J.; Schwarz, S.; Couto, I. Plasmid-borne antimicrobial resistance of Staphylococcus aureus isolated in a hospital in Lisbon, Portugal. Microb. Drug Resist. 2016, 22, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Anthonisen, I.L.; Sunde, M.; Steinum, T.M.; Sidhu, M.S.; Sorum, H. Organization of the antiseptic resistance gene qacA and Tn552-related-lactamase genes in multidrug- resistant Staphylococcus haemolyticus strains of animal and human origins. Antimicrob. Agents. Chemother. 2002, 46, 3606–3612. [Google Scholar] [CrossRef] [Green Version]
- Bjorland, J.; Sunde, M.; Waage, S. Plasmid-borne smr gene causes resistance to quaternary ammonium compounds in bovine Staphylococcus aureus. J. Clin. Microbiol. 2001, 39, 3999–4004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, M.; McCusker, M.; Viveiros, M.; Couto, I.; Fanning, S.; Pagès, J.M. A Simple Method for Assessment of MDR Bacteria for Over-Expressed Efflux Pumps. Open Microbiol. J. 2013, 7, 72–82. [Google Scholar] [CrossRef]
- Andrade, M.; Oliveira, K.; Morais, C.; Abrantes, P.; Pomba, C.; Rosato, A.E.; Couto, I.; Costa, S.S. Virulence potential of biofilm producing Staphylococcus pseudintermedius, Staphylococcus aureus and Staphylococcus coagulans causing skin infections in companion animals. Antibiotics 2022, 11, 1339. [Google Scholar] [CrossRef]
- Descloux, S.; Rossano, A.; Perreten, V. Characterization of new Staphylococcal Cassette Chromosome mec (SCCmec) and topoisomerase genes in fluoroquinolone- and methicillin-Resistant Staphylococcus pseudintermedius. J. Clin. Microbiol. 2008, 46, 1818–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, D. Emerging mechanisms of fluoroquinolones resistance. Emerg Infect. Dis. 2001, 7, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kizerwetter-Świda, M.; Chrobak-Chmiel, D.; Rzewuska, M.; Binek, M. Resistance of canine methicillin-resistant Staphylococcus pseudintermedius strains to pradofloxacin. J. Vet. Diagnostic. Investig. 2016, 28, 514–518. [Google Scholar] [CrossRef] [Green Version]
- Azzariti, S.; Bond, R.; Loeffler, A.; Zendri, F.; Timofte, D.; Chang, Y.; Pelligand, L. Investigation of in vitro susceptibility and resistance mechanisms in skin pathogens: Perspectives for fluoroquinolone therapy in canine pyoderma. Antibiotics 2022, 11, 1204. [Google Scholar] [CrossRef]
- Couto, N.; Belas, A.; Couto, I.; Perreten, V.; Pomba, C. Genetic relatedness, antimicrobial and biocide susceptibility comparative analysis of methicillin-resistant and -susceptible Staphylococcus pseudintermedius from Portugal. Microb. Drug Resist. 2013, 20, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, R. Resistência a Antimicrobianos em Staphylococcus aureus Isolados de Animais de Companhia. Master’s Thesis, Instituto de Higiene e Medicina Tropical, Lisbon, Portugal, 2019. [Google Scholar]
- Costa, S.S.; Rosa, M.; Rodrigues, A.C.; Holtreman, F.; Viveiros, M.; Pomba, C.; Couto, I. Emergence of non-wild-type populations towards biocides and antibiotic resistance in Staphylococcus epidermidis colonizing veterinary staff and students. ePoster Mini-Oral Session. In Proceedings of the 28th European Congress on Clinical Microbiology and Infectious Diseases (ECCMID), Madrid, Spain, 21–24 April 2018. [Google Scholar]
- Bergot, M.; Martins-Simões, P.; Kilian, H.; Châtre, P.; Worthing, K.; Norris, J.; Madec, J.; Laurent, F.; Haenni, M. Evolution of the population structure of Staphylococcus pseudintermedius in France. Front. Microbiol. 2021, 9, 3055. [Google Scholar] [CrossRef] [PubMed]
- Rynhoud, H.; Forde, M.; Beatson, S.; Abraham, S.; Meler, E.; Magalhães, R.; Gibson, J. Molecular epidemiology of clinical and colonizing methicillin-resistant Staphylococcus isolates in companion animals. Front. Vet. Sci. 2021, 8, 620491. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.; Oliveira, A.; Manageiro, V.; Caniça, M.; Contente, D.; Capita, R.; Alonso-Calleja, C.; Carvalho, I.; Capelo, J.L.; Igrejas, G.; et al. Clonal diversity and antimicrobial resistance of methicillin-resistant Staphylococcus pseudintermedius isolated from canine pyoderma. Microorganisms 2021, 9, 482. [Google Scholar] [CrossRef] [PubMed]
- Tambat, R.; Jangra, M.; Mahey, N.; Chandal, N.; Kaur, M.; Chaudhary, S.; Verma, D.K.; Thakur, K.G.; Raje, M.; Jachak, S.; et al. Microbe-derived indole metabolite demonstrates potent multidrug efflux pump inhibition in Staphylococcus aureus. Front. Microbiol. 2019, 10, 2153. [Google Scholar] [CrossRef] [PubMed]
- Baiomy, A.; Shaker, G.; Abbas, H. Sensitizing multi drug resistant Staphylococcus aureus isolated from surgical site infections to antimicrobials by efflux pumps inhibitors. Afr. Health Sci. 2020, 20, 1632–1645. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, B.V.; Babu, T.M.C.; Reddy, N.V.; Rajendra, W. Homology modeling, molecular dynamics, and virtual screening of NorA efflux pump inhibitors of Staphylococcus aureus. Drug Des. Devel. Ther. 2016, 10, 3237–3252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zárate, S.G.; Morales, P.; Świderek, K.; Bolanos-Garcia, V.M.; Bastida, A. Molecular modeling approach to identify novel inhibitors of the major facilitator superfamily of efflux pump transporters. Antibiotics 2019, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buffet-Bataillon, S.; Tattevin, P.; Maillard, J.Y.; Bonnaure-Mallet, M.; Jolivet Gougeon, A. Efflux pump induction by quaternary ammonium compounds and fluoroquinolone resistance in bacteria. Future Microbiol. 2016, 11, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Lekshmi, M.; Ammini, P.; Adjei, J.; Sanford, L. Modulation of antimicrobial efflux pumps of the major facilitator superfamily in Staphylococcus aureus. AIMS Microbiol. 2018, 4, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Beco, L.; Guaguère, E.; Lorente Méndez, C.; Noli, C.; Nuttall, T.; Vroom, M. Suggested guidelines for using systemic antimicrobials in bacterial skin infections: Part 2-antimicrobial choice, treatment regimens and compliance. Vet. Rec. 2013, 172, 156–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Medicines Agency (EMA). Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2021; European Medicines Agency (EMA): Amesterdam, The Netherlands, 2022.
- Oliveira, A.; Devesa, J.S.P.; Hill, P.B.; Silva, V.; Poeta, P. Treatment of selected canine dermatological conditions in Portugal—A research survey. J. Vet. Res. 2018, 62, 563–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Poveda, B.; Moreno, M. Antimicrobial prescriptions for dogs in the Capital of Spain. Front. Vet. Sci. 2018, 5, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loiacono, M.; Martino, P.A.; Albonico, F.; Dell’Orco, F.; Ferretti, M.; Zanzani, S.; Mortarino, M. High-resolution melting analysis of gyrA codon 84 and grlA codon 80 mutations conferring resistance to fluoroquinolones in Staphylococcus pseudintermedius isolates from canine clinical samples. J. Vet. Diagn. Investig. 2017, 29, 711–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadlec, K.; Schwarz, S. Antimicrobial resistance of Staphylococcus pseudintermedius. Vet. Dermatol. 2012, 23, 276-e55. [Google Scholar] [CrossRef]
- Onuma, K.; Tanabe, T.; Sato, H. Antimicrobial resistance of Staphylococcus pseudintermedius isolates from healthy dogs and dogs affected with pyoderma in Japan. Vet. Dermatol. 2012, 23, 17-e5. [Google Scholar] [CrossRef]
- Awji, E.G.; Tassew, D.D.; Lee, J.S.; Lee, S.J.; Choi, M.J.; Reza, M.A.; Rhee, M.H.; Kim, T.H.; Park, S.C. Comparative mutant prevention concentration and mechanism of resistance to veterinary fluoroquinolones in Staphylococcus pseudintermedius. Vet. Dermatol. 2012, 23, 376-e69. [Google Scholar] [CrossRef]
- Moore, L.E.; Ledder, R.G.; Gilbert, P.; McBain, A.J. In vitro study of the effect of cationic biocides on bacterial population dynamics and susceptibility. Appl. Environ. Microbiol. 2008, 74, 4825–4834. [Google Scholar] [CrossRef] [Green Version]
- Martín-Rodríguez, A.J.; Babarro, J.M.F.; Lahoz, F.; Sansón, M.; Martín, V.S.; Norte, M.; Fernandéz, J.J. From broad-spectrum biocides to quorum sensing disruptors and mussel repellents: Antifouling profile of alkyl triphenylphosphonium salts. PLoS ONE 2015, 10, e0123652. [Google Scholar] [CrossRef] [Green Version]
- McDonnell, G. Antisepsis, Disinfection, and Sterilization: Types, Action, and Resistance, 2nd ed.; ASM Press: Washigton, DC, USA, 2017; pp. 167–183. [Google Scholar]
- Lei, Q.; Lai, X.; Zhang, Y.; Li, Z.; Li, R.; Zhang, W.; Ao, N.; Zhang, H. PEGylated Bis-Quaternary Triphenyl-Phosphonium Tosylate allows for balanced antibacterial activity and cytotoxicity. Appl. Bio Mater. 2020, 3, 6400–6407. [Google Scholar] [CrossRef]
- Kanazawa, A.; Ikeda, T.; Endo, T. A novel approach to mode of action of cationic biocides: Morphological effect on antibacterial activity. J. Appl. Bacteriol. 1995, 78, 55–60. [Google Scholar] [CrossRef]
- Kaatz, G.; Seo, S.; O’Brien, L.; Wahiduzzaman, M.; Foster, T.J. Evidence for the existence of a multidrug efflux transporter distinct from NorA in Staphylococcus aureus. Antimicrob. Agents. Chemother. 2000, 44, 1404–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, A.; Sellon, D.; Gay, J.; Lofgren, E.; Moore, D.; Jones, L.; Davis, M.A. Prevalence of methicillin-resistant Staphylococcus pseudintermedius on hand-contact and animal-contact surfaces in companion animal community hospitals. Can. Vet. J. La Rev. Vét. Can. 2020, 61, 613–620. [Google Scholar]
- Murphy, C.; Reid-Smith, R.; Boerlin, P.; Weese, J.S.; Prescott, J.; Janecko, N.; Hassard, L.; McEwen, S.A. Escherichia coli and selected veterinary and zoonotic pathogens isolated from environmental sites in companion animal veterinary hospitals in southern Ontario. Can. Vet. J. 2010, 51, 963–972. [Google Scholar]
- Walker, M.A.; Singh, A.; Gibson, T.W.; Rousseau, J.; Weese, J.S. Presence of Qac genes in clinical isolates of methicillin-resistant and methicillin-susceptible Staphylococcus pseudintermedius and their impact on chlorhexidine digluconate susceptibility. Vet. Surg. 2020, 49, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Vali, L.; Davies, S.E.; Lai, L.L.G.; Dave, J.; Amyes, S.G.B. Frequency of biocide resistance genes, antibiotic resistance and the effect of chlorhexidine exposure on clinical methicillin-resistant Staphylococcus aureus isolates. J. Antimicrob. Chemother. 2008, 61, 524–532. [Google Scholar] [CrossRef]
- Valentine, B.K.; Dew, W.; Yu, A.; Weese, J.S. In vitro evaluation of topical biocide and antimicrobial susceptibility of Staphylococcus pseudintermedius from dogs. Vet. Dermatol. 2012, 23, 493-e95. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.M.; Loeffler, A.; Bond, R. Susceptibility in vitro of canine methicillin-resistant and -susceptible staphylococcal isolates to fusidic acid, chlorhexidine and miconazole: Opportunities for topical therapy of canine superficial pyoderma. J. Antimicrob. Chemother. 2015, 70, 2048–2052. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COWT 99% | SD (log2) | WT Population | NWT Population | |||
|---|---|---|---|---|---|---|
| X ≤ COWT | No. Strains (%) | X > COWT | No. Strains (%) | |||
| EtBr | 4 mg/L | 0.56 | ≤4 mg/L | 155 (100%) | >4 mg | 0 (0%) |
| TPP | 16 mg/L | 0.52 | ≤16 mg/L | 137 (88.4%) | >16 mg/L | 18 (11.6%) |
| Category | Strain | Characteristics 1,2 | Mutations in QRDR 1 | MIC (mg/L) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MRSP MSSP | MDR | FQ | ST-agr | GrlA | GyrA | TPP | +TZ | +VER | EtBr | +TZ | +VER | CIP | +TZ | +VER | ||
| Type strain | DSM21284T | MSSP | MDR | FQS | ST63-agrIV 3 | - | - | 8 | 4 | 2 | 1 | <0.06 | <0.06 | 0.125 | 0.125 | 0.125 |
| TPPNWT | BIOS-V40 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 16 | 4 | 2 | 0.5 | 0.125 | 64 | 32 | 64 |
| BIOS-V52 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 8 | 4 | 2 | 0.5 | 0.125 | 64 | 32 | 64 | |
| BIOS-V53 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 8 | 4 | 1 | 0.5 | <0.06 | 64 | 64 | 32 | |
| BIOS-V64 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 8 | 4 | 1 | 0.5 | <0.06 | 64 | 64 | 64 | |
| BIOS-V83 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 64 | 16 | 4 | 1 | 0.5 | <0.06 | 64 | 32 | 64 | |
| BIOS-V99 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 8 | 4 | 1 | 0.25 | <0.06 | 64 | 32 | 64 | |
| BIOS-V104 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 4 | 1 | 1 | 0.125 | <0.06 | 32 | 32 | 32 | |
| BIOS-V108 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 4 | 4 | 2 | 0.5 | <0.06 | 32 | 32 | 32 | |
| BIOS-V125 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 8 | 4 | 2 | 1 | <0.06 | 64 | 32 | 64 | |
| BIOS-V131 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 8 | 4 | 2 | 0.125 | <0.06 | 64 | 64 | 64 | |
| BIOS-V143 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 64 | 8 | 8 | 2 | 0.5 | 0.125 | 64 | 32 | 32 | |
| BIOS-V144 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 8 | 8 | 2 | 0.25 | 0.125 | 64 | 32 | 32 | |
| BIOS-V146 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 64 | 8 | 4 | 1 | 0.5 | 0.125 | 64 | 32 | 64 | |
| BIOS-V207 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 8 | 4 | 2 | 0.125 | 0.125 | 64 | 64 | 64 | |
| BIOS-V223 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 8 | 4 | 2 | 0.5 | <0.06 | 64 | 32 | 64 | |
| BIOS-V234 | MSSP | No | FQS | ST2194-agrIII | - | - | 64 | 16 | 8 | 2 | 0.5 | 0.125 | 0.25 | 0.125 | 0.125 | |
| BIOS-V262 | MRSP | MDR | FQR | ST118-agrII | S80I | S84L | 128 | 16 | 8 | 4 | 0.5 | 0.125 | 64 | 32 | 64 | |
| BIOS-V299 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 32 | 8 | 4 | 1 | 0.5 | 0.125 | 32 | --- | --- | |
| TPPWT | BIOS-V7 | MRSP | MDR | FQS | ST157-agrIV | - | - | 4 | --- | --- | 1 | 0.06 | 0.03 | 1 | 1 | 1 |
| BIOS-V10 | MSSP | No | FQS | ST2095-agrIV | - | - | 8 | 4 | 4 | 1 | 0.25 | 0.125 | 0.06 | 0.06 | 0.06 | |
| BIOS-V11 | MSSP | No | FQS | ST2054-agrIII | - | - | 4 | 4 | --- | 1 | 0.125 | 0.03 | 0.06 | 0.06 | 0.06 | |
| BIOS-V15 | MSSP | No | FQS | ---/agrIII | - | - | 8 | 4 | 2 | 2 | 0.125 | <0.06 | 0.125 | 0.125 | 0.125 | |
| BIOS-V25 | MRSP | MDR | FQS | ST157-agrIV | - | - | 4 | --- | --- | 2 | 0.125 | 0.5 | 1 | 0.5 | 0.5 | |
| BIOS-V26 | MRSP | No | FQS | ST157-agrIV | - | - | 8 | 8 | 4 | 0.5 | 0.06 | 0.03 | 0.25 | 0.125 | 0.125 | |
| BIOS-V29 | MRSP | No | FQS | ST2055-agrI | - | - | 8 | 4 | 4 | 1 | 0.03 | 0.03 | 0.125 | 0.125 | 0.125 | |
| BIOS-V37 | MSSP | No | FQS | ---/agrIV | - | - | 8 | 1 | 1 | 2 | <0.06 | <0.06 | 0.25 | 0.125 | 0.125 | |
| BIOS-V39 | MSSP | MDR | FQS | ST2096-agrIII | S80I | - | 8 | 4 | 4 | 2 | 0.125 | 0.03 | 1 | 0.25 | 0.5 | |
| BIOS-V48 | MSSP | No | FQS | ---/agrIII | - | - | 4 | --- | --- | 0.5 | 0.125 | 0.0015 | 0.25 | 0.06 | 0.06 | |
| TPPWT | BIOS-V65 | MSSP | No | FQS | ---/agrIV | - | - | 8 | 4 | 4 | 2 | 0.125 | 0.03 | 0.25 | 0.25 | 0.25 |
| BIOS-V68 | MSSP | MDR | FQS | ST2097-agrIV | --- | --- | 8 | 1 | 1 | 1 | <0.06 | <0.06 | 0.5 | 0.5 | 0.5 | |
| BIOS-V79 | MSSP | MDR | FQS | ST2098-agrI | --- | --- | 8 | 8 | --- | 1 | 0.125 | 0.03 | 0.125 | 0.125 | 0.125 | |
| BIOS-V84 | MSSP | MDR | FQR | ST2099-agrIV | D84G | S84L | 8 | --- | --- | 1 | 0.06 | 0.06 | 8 | 8 | 8 | |
| BIOS-V90 | MSSP | No | FQS | ST2100-agrIII | --- | --- | 8 | 8 | 4 | 1 | 0.125 | 0.06 | 0.25 | 0.125 | 0.25 | |
| BIOS-V96 | MSSP | MDR | FQS | ST2101-agrIV | S80R | - | 4 | 4 | 2 | 1 | 0.06 | 0.06 | 1 | 0.5 | 0.5 | |
| BIOS-V97 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 16 | 4 | 4 | 1 | 0.25 | 0.125 | 64 | 32 | 32 | |
| BIOS-V101 | MSSP | MDR | FQR | ST2102-agrII | S80R | D83N | 8 | 4 | 4 | 1 | 0.06 | 0.06 | 2 | 2 | 2 | |
| BIOS-V103 | MSSP | No | FQS | ---/agrII | S80R | - | 8 | 8 | 8 | 1 | 0.5 | 0.5 | 1 | 0.5 | 0.5 | |
| BIOS-V105 | MRSP | MDR | FQR | ST2056-agrII | S80I | S84L | 8 | 4 | 4 | 1 | 0.125 | 0.06 | 64 | 32 | 32 | |
| BIOS-V106 | MSSP | No | FQS | ---/agrIII | - | - | 4 | --- | --- | 1 | 0.25 | 0.06 | 0.25 | 0.25 | 0.25 | |
| BIOS-V116 | MSSP | MDR | FQS | ---/agrIV | S80R | - | 4 | --- | --- | 1 | --- | --- | 1 | 0.5 | 0.5 | |
| BIOS-V119 | MRSP | MDR | FQS | ST2057-agrIII | - | - | 4 | 4 | 4 | 0.5 | 0.06 | 0.015 | 0.25 | 0.25 | 0.25 | |
| BIOS-V120 | MSSP | No | FQS | ST2103-agrIII | --- | --- | 4 | 2 | 1 | 1 | <0.06 | 0.03 | 0.06 | 0.06 | 0.06 | |
| BIOS-V122 | MRSP | MDR | FQS | ST2104-agrIV | S80I | - | 8 | 8 | 4 | 1 | 0.125 | 0.06 | 2 | 0.5 | 1 | |
| BIOS-V127 | MRSP | MDR | FQS | ST157-agrIV | S80I | - | 16 | 16 | 8 | 2 | 0.25 | 0.03 | 1 | 1 | 1 | |
| BIOS-V136 | MSSP | MDR | FQS | ST2058-agrIII | - | - | 4 | 2 | 1 | 1 | 0.125 | 0.03 | 0.25 | 0.125 | 0.25 | |
| BIOS-V164 | MSSP | MDR | FQS | ST1350-agrIV | - | - | 16 | 8 | 8 | 1 | 0.125 | 0.06 | 0.125 | 0.06 | 0.06 | |
| BIOS-V175 | MSSP | MDR | FQS | ST555-agrIV | --- | --- | 4 | 2 | --- | 1 | 0.06 | 0.03 | 0.06 | 0.06 | 0.06 | |
| BIOS-V176 | MSSP | No | FQS | ---/agrII | S80I | - | 8 | --- | --- | 2 | 0.5 | 0.125 | 1 | 0.5 | 0.5 | |
| BIOS-V179 | MRSP | MDR | FQR | ST2059-agrIII | D84N | S84L | 8 | 8 | 8 | 1 | 0.06 | 0.125 | 4 | 2 | 4 | |
| BIOS-V190 | MSSP | MDR | FQS | ST2106-agrI | --- | --- | 8 | 8 | 4 | 0.5 | 0.03 | 0.03 | 0.06 | 0.06 | 0.06 | |
| BIOS-V211 | MSSP | No | FQS | ST2107-agrIV | --- | --- | 8 | --- | --- | 1 | 0.25 | 0.06 | 0.25 | 0.125 | 0.25 | |
| BIOS-V212 | MSSP | No | FQS | ST1183-agrIV | S80I | - | 8 | 8 | 4 | 2 | 0.25 | 0.06 | 0.125 | 0.06 | 0.06 | |
| BIOS-V213 | MRSP | MDR | FQR | ST422-agrIII | S80I | S84L | 8 | --- | --- | 1 | 0.125 | 0.125 | 64 | 32 | 64 | |
| BIOS-V217 | MRSP | MDR | FQS | ST2060-agrIV | - | - | 8 | 4 | 4 | 1 | 0.06 | 0.03 | 0.25 | 0.125 | 0.125 | |
| BIOS-V218 | MSSP | MDR | FQS | ST241-agrIII | - | - | 8 | 8 | 8 | 1 | 0.25 | 0.125 | 0.25 | 0.06 | 0.25 | |
| BIOS-V224 | MRSP | MDR | FQR | ST45-agrII | S80I | S84L | 8 | 4 | 2 | 1 | 0.125 | 0.125 | 64 | 64 | 64 | |
| BIOS-V225 | MSSP | No | FQS | ST455-agrIII | - | - | 8 | 4 | 4 | 1 | 0.5 | 0.125 | 0.125 | 0.125 | 0.125 | |
| BIOS-V227 | MRSP | MDR | FQR | ST551-agrIII | S80I | S84L | 8 | 4 | 4 | 1 | 0.125 | 0.06 | 32 | 32 | 32 | |
| BIOS-V228 | MRSP | MDR | FQR | ST45-agrII | S80I | S84L | 8 | --- | --- | 1 | 0.125 | 0.125 | 64 | 32 | 32 | |
| BIOS-V230 | MSSP | No | FQS | ---/agrIV | --- | --- | 8 | 4 | 4 | 1 | 0.06 | 0.03 | 0.06 | 0.06 | 0.06 | |
| BIOS-V231 | MSSP | No | FQS | ---/agrIV | - | - | 8 | 8 | 4 | 2 | 0.25 | 0.125 | 0.125 | 0.06 | 0.125 | |
| BIOS-V240 | MRSP | MDR | FQR | ST2061-agrIII | S80I | S84L | 8 | 8 | 8 | 4 | 1 | 0.125 | 64 | 32 | 64 | |
| BIOS-V242 | MSSP | MDR | FQS | ST2108-agrII | S80I | - | 8 | 4 | 4 | 2 | 0.06 | 0.03 | 1 | 1 | 0.5 | |
| BIOS-V264 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 16 | 4 | 2 | 4 | 1 | 0.25 | 64 | 32 | 64 | |
| TPPWT | BIOS-V268 | MSSP | MDR | FQS | ---/agrII | - | - | 4 | 1 | 2 | 1 | <0.06 | <0.06 | 0.25 | 0.125 | 0.125 |
| BIOS-V270 | MRSP | MDR | FQR | ST25-agrIII | S80I | S84L | 8 | 4 | 2 | 1 | 0.25 | 0.125 | 32 | 16 | 32 | |
| BIOS-V276 | MRSP | MDR | FQS | ST497-agrIII | D84G | - | 16 | 4 | 1 | 2 | 0.125 | <0.06 | 1 | 0.5 | 0.5 | |
| BIOS-V280 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 16 | 8 | 4 | 4 | 0.25 | <0.06 | 64 | 32 | 64 | |
| BIOS-V292 | MRSP | MDR | FQR | ST45-agrII | S80I | S84L | 8 | 4 | 4 | 1 | 0.125 | 0.125 | 32 | 32 | 32 | |
| BIOS-V302 | MRSP | MDR | FQR | ST71-agrIII | S80I | S84L | 16 | 8 | 2 | 4 | 0.5 | 0.125 | 64 | 32 | 64 | |
| Category | Strain | Relative Final Fluorescence (RFF) | |||
|---|---|---|---|---|---|
| −Glucose +TZ | −Glucose +VER | +Glucose +TZ | +Glucose +VER | ||
| Type strain | DSM21284T | −0.24 ± 0.00 | 0.54 ± 0.03 | 0.61 ± 0.11 | 1.05 ± 0.11 |
| TPPWT | BIOS-V37 | −0.37 ± 0.02 | −0.09 ± 0.14 | 0.65 ± 0.16 | 1.18 ± 0.14 |
| TPPNWT Subgroup 2 | BIOS-V104 | 0.00 ± 0.08 | 1.12 ± 0.41 | 0.93 ± 0.16 | 1.81 ± 0.17 |
| BIOS-V143 | −0.13 ± 0.05 | 0.12 ± 0.05 | 0.82 ± 0.40 | 1.36 ± 0.68 | |
| BIOS-V234 | −0.38 ± 0.11 | −0.23 ± 0.04 | 0.89 ± 0.14 | 1.59 ± 0.03 | |
| TPPNWT Subgroup 3 | BIOS-V99 | −0.18 ± 0.13 | 0.63 ± 0.20 | 0.30 ± 0.09 | 0.76 ± 0.33 |
| BIOS-V262 | −0.40 ± 0.11 | 0.77 ± 0.66 | 1.80 ± 0.30 | 2.07 ± 0.04 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leal, M.; Morais, C.; Ramos, B.; Pomba, C.; Abrantes, P.; Costa, S.S.; Couto, I. Exploring Efflux as a Mechanism of Reduced Susceptibility towards Biocides and Fluoroquinolones in Staphylococcus pseudintermedius. Animals 2023, 13, 1270. https://doi.org/10.3390/ani13071270
Leal M, Morais C, Ramos B, Pomba C, Abrantes P, Costa SS, Couto I. Exploring Efflux as a Mechanism of Reduced Susceptibility towards Biocides and Fluoroquinolones in Staphylococcus pseudintermedius. Animals. 2023; 13(7):1270. https://doi.org/10.3390/ani13071270
Chicago/Turabian StyleLeal, Marta, Catarina Morais, Bárbara Ramos, Constança Pomba, Patrícia Abrantes, Sofia Santos Costa, and Isabel Couto. 2023. "Exploring Efflux as a Mechanism of Reduced Susceptibility towards Biocides and Fluoroquinolones in Staphylococcus pseudintermedius" Animals 13, no. 7: 1270. https://doi.org/10.3390/ani13071270