
A Quieter Ocean: Experimentally Derived Differences in Attentive Responses of Tursiops truncatus to Anthropogenic Noise Playbacks before and during the COVID-19-Related Anthropause


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Cetaceans and Anthropauses
1.2. Noise and Cetaceans
1.3. Habituation and Sensitization
2. Materials and Methods
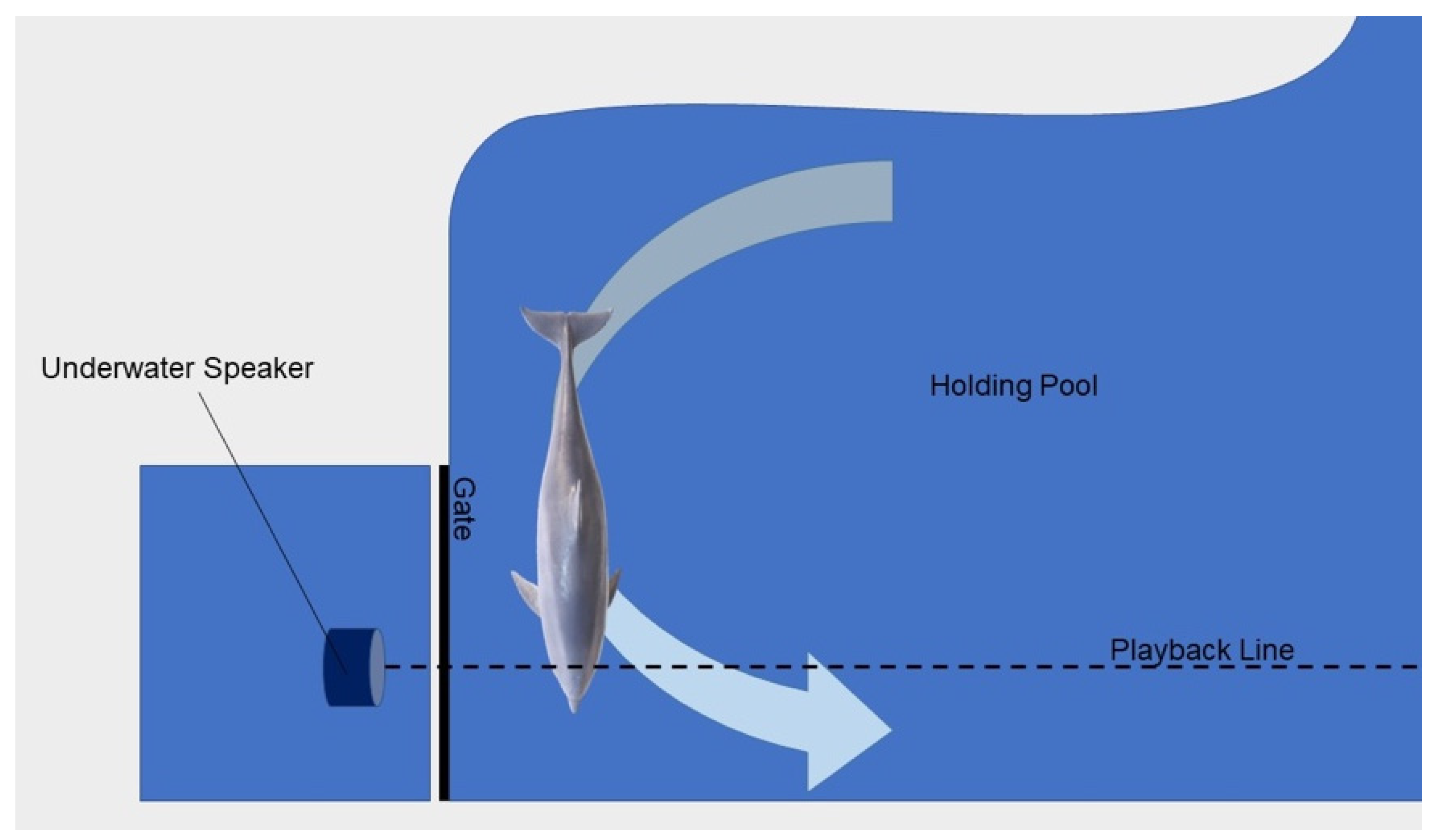
2.1. Setting
2.2. Subjects
2.3. Noise Exposure
2.4. Playback Presentation
2.5. Behavior Scoring
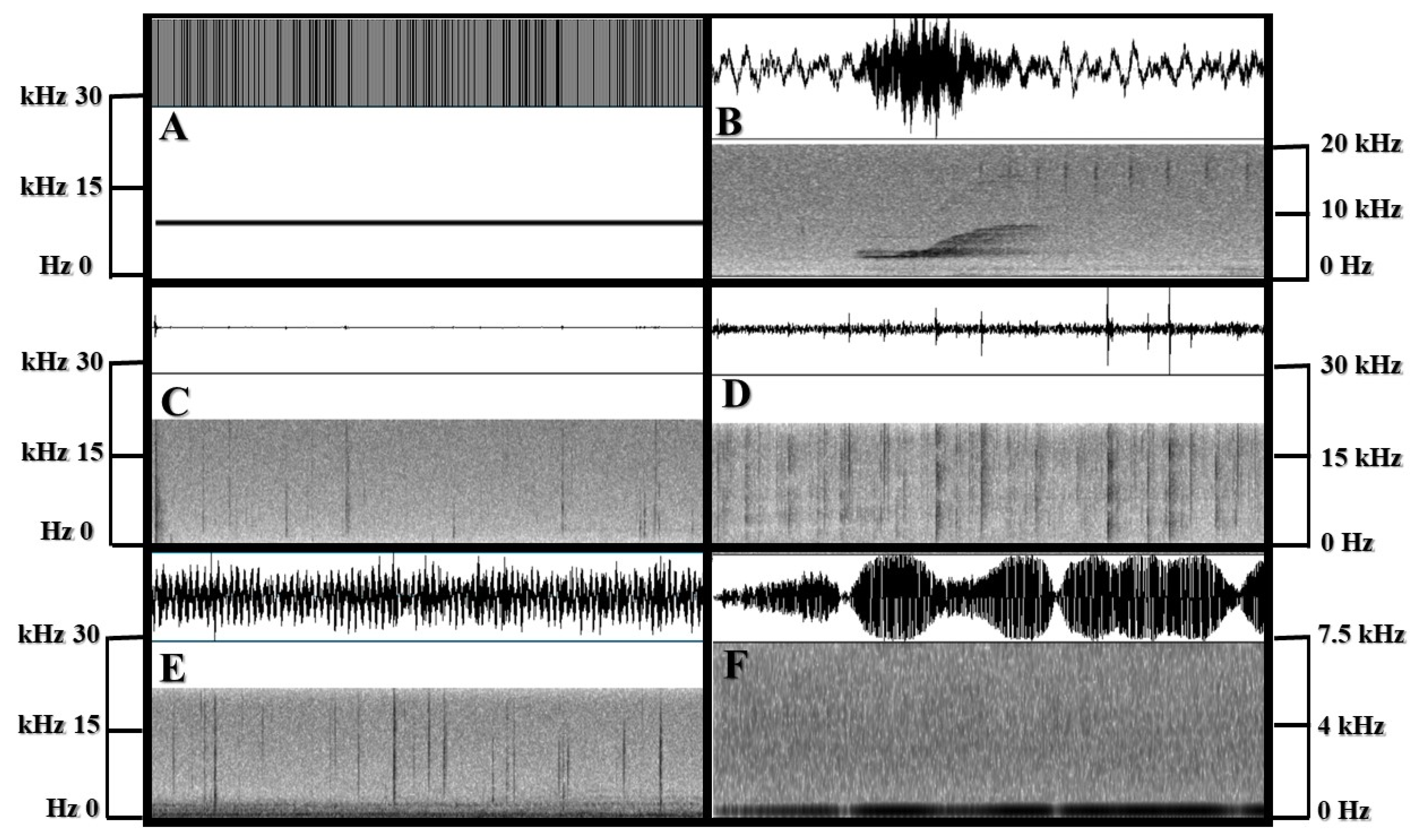
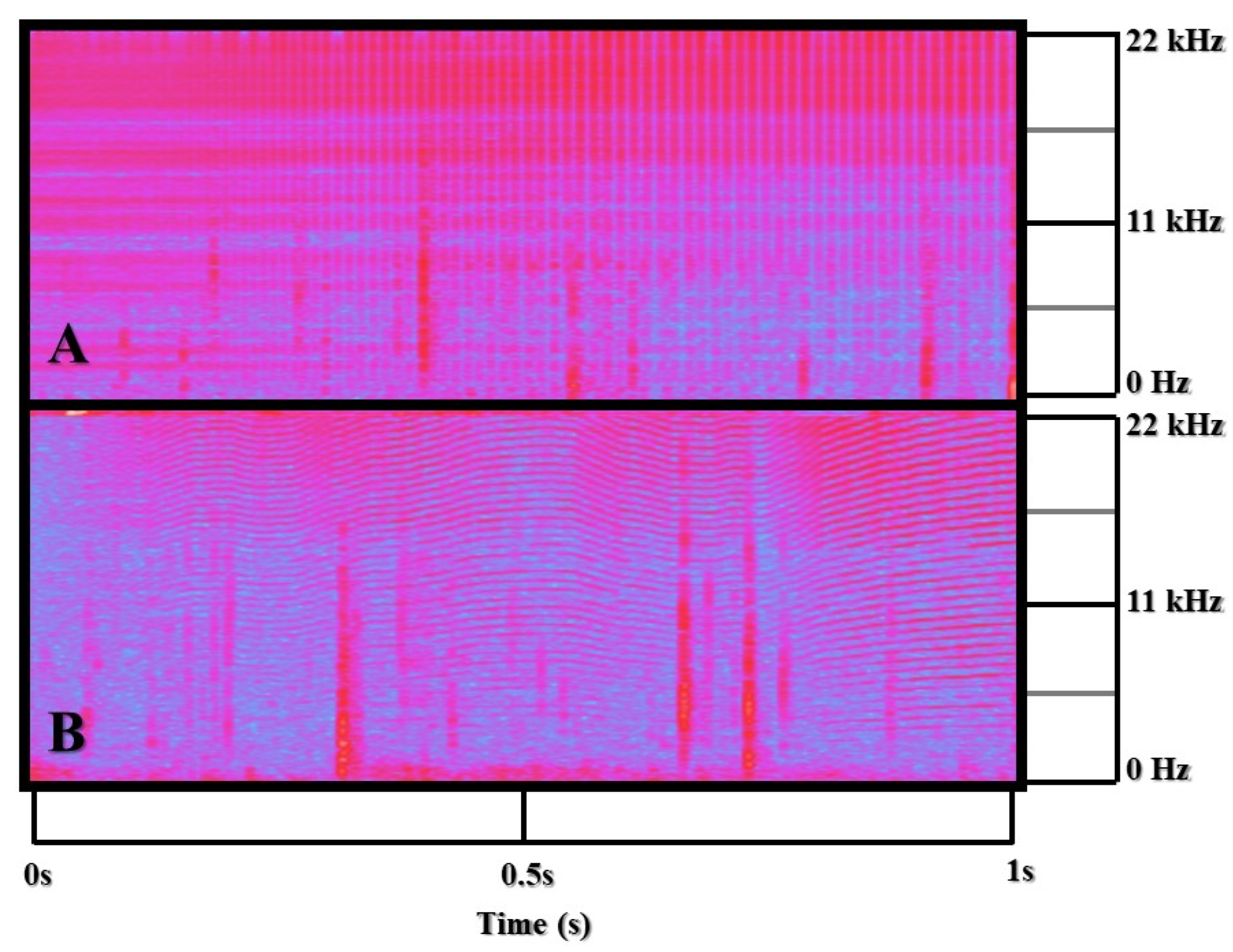
2.6. Acoustic Analysis
2.7. Statistical Analysis
3. Results
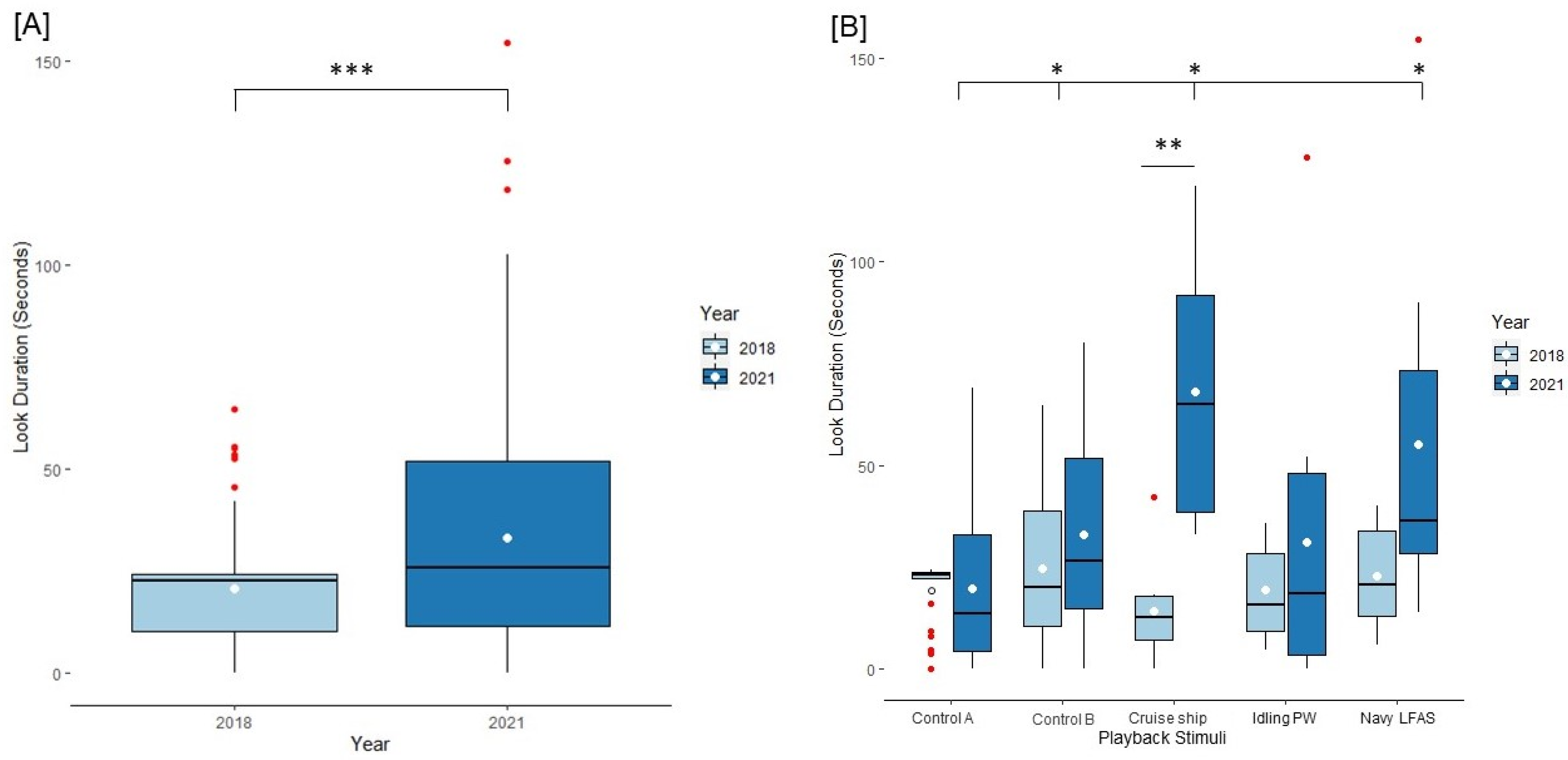
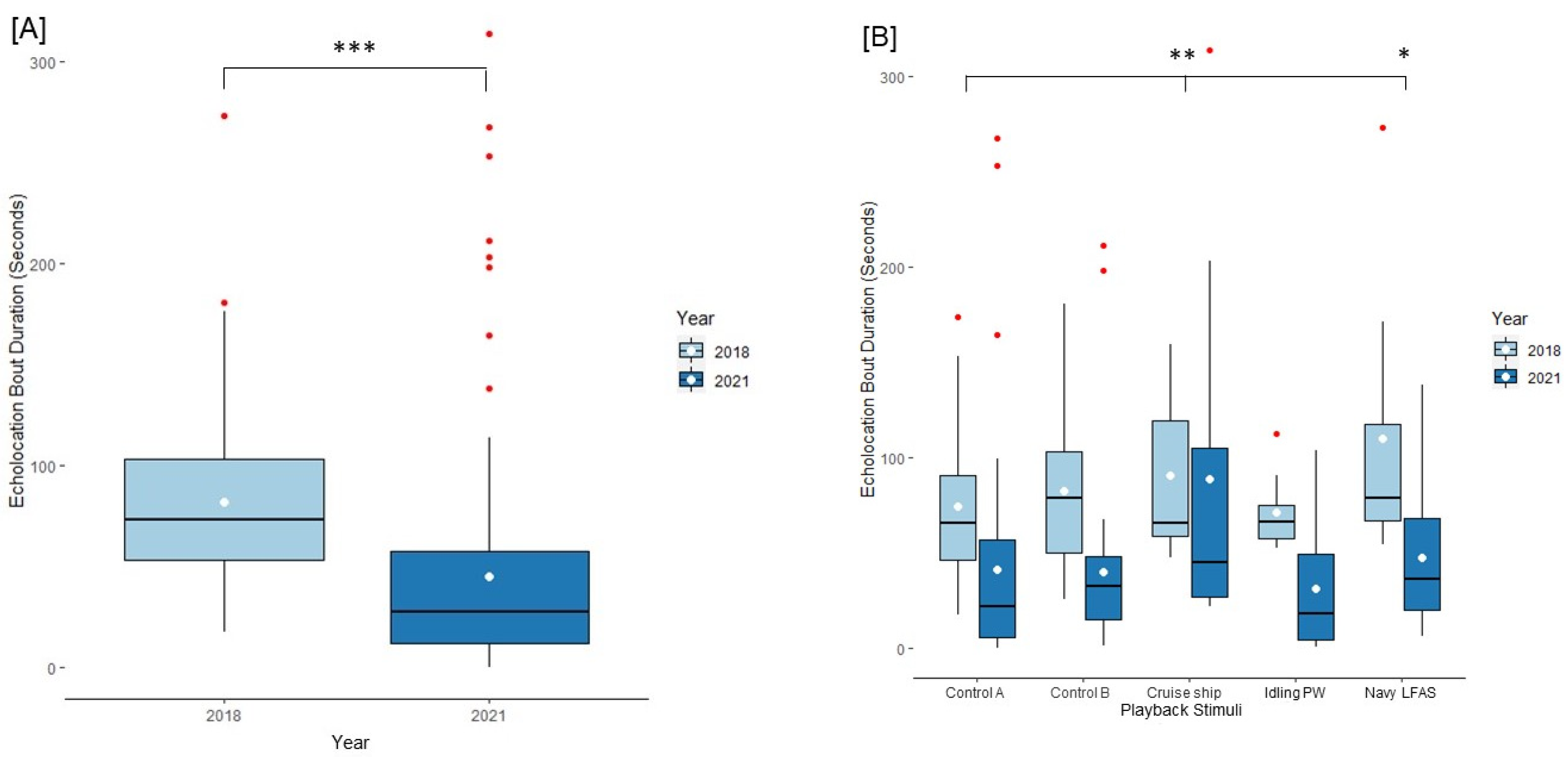
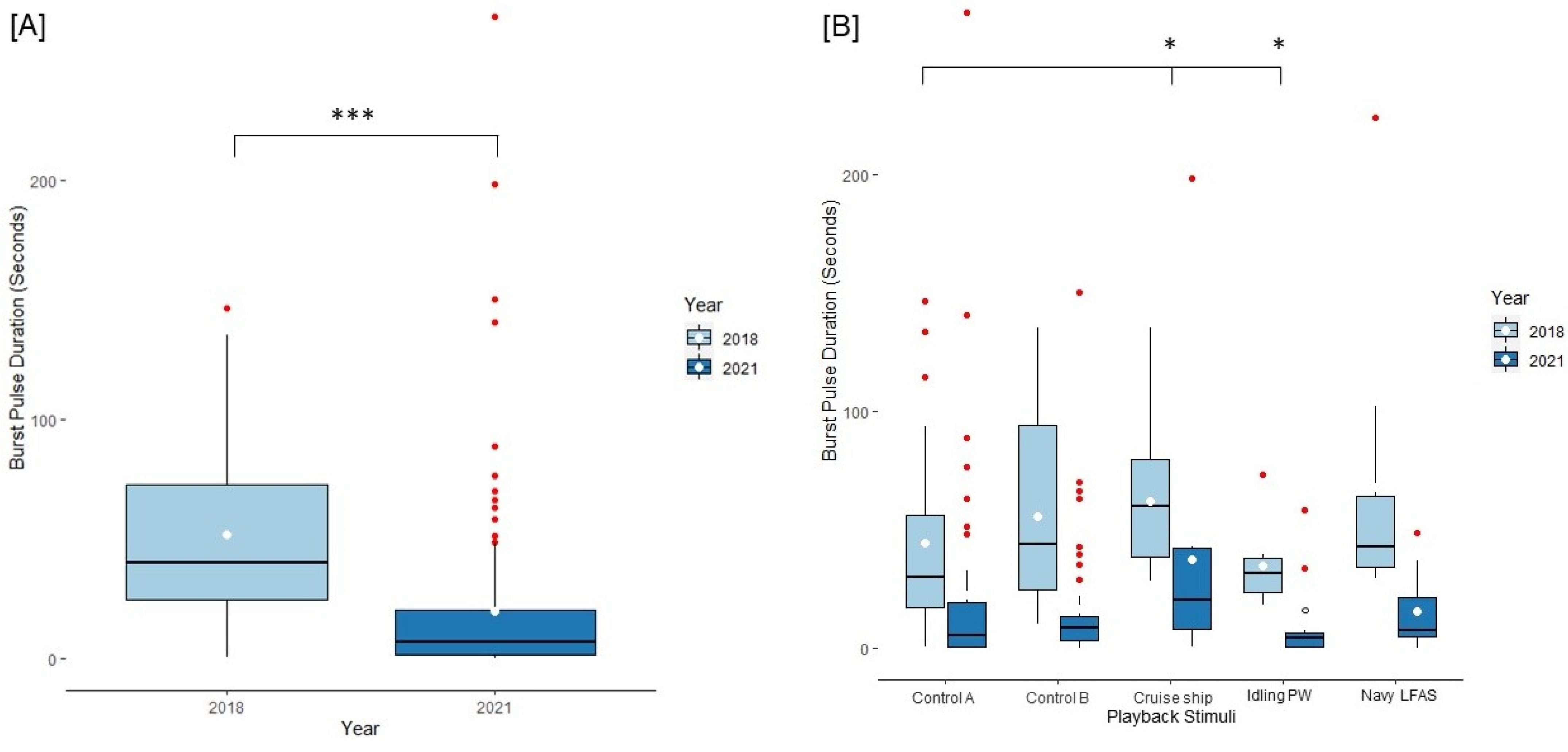
3.1. Behavioral Results
3.2. Acoustic Results
3.3. Results of Dolphins Tested across Both Years
4. Discussion
4.1. COVID-19 Anthropause and Investigative Behaviors
4.2. Anthropause and Acoustic Response
4.3. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duarte, C.M.; Chapuis, L.; Collin, S.P.; Costa, D.P.; Devassy, R.P.; Eguiluz, V.M.; Erbe, C.; Gordon, T.A.C.; Halpern, B.S.; Harding, H.R.; et al. The soundscape of the Anthropocene ocean. Science 2021, 371, eaba4658. [Google Scholar] [CrossRef] [PubMed]
- Perry, C. A review of the impact of anthropogenic noise on cetaceans. In Proceedings of the Scientific Committee at the 50th Meeting of the International Whaling Commission, Muscat, Oman, 16–20 May 1998; p. 3. [Google Scholar]
- Williams, R.; Wright, A.J.; Ashe, E.; Blight, L.K.; Bruintjes, R.; Canessa, R.; Clark, C.W.; Cullis-Suzuki, S.; Dakin, D.T.; Erbe, C.; et al. Impacts of anthropogenic noise on marine life: Publication patterns, new discoveries, and future directions in research and management. Ocean Coast. Manag. 2015, 115, 17–24. [Google Scholar] [CrossRef] [Green Version]
- McGregor, P.K.; Horn, A.G.; Leonard, M.L.; Thomsen, F. Anthropogenic noise and conservation. In Animal Communication and Noise; Springer: Berlin/Heidelberg, Germany, 2013; pp. 409–444. [Google Scholar]
- Weilgart, L.S. The impacts of anthropogenic ocean noise on cetaceans and implications for management. Can. J. Zool. 2007, 85, 1091–1116. [Google Scholar] [CrossRef]
- Tyack, P.L.; Thomas, L. Using dose–response functions to improve calculations of the impact of anthropogenic noise. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 242–253. [Google Scholar] [CrossRef] [Green Version]
- Erbe, C.; Dunlop, R.; Dolman, S. Effects of noise on marine mammals. In Effects of Anthropogenic Noise on Animals; Springer: Berlin/Heidelberg, Germany, 2018; pp. 277–309. [Google Scholar]
- Zirbel, K.; Balint, P.; Parsons, E. Navy sonar, cetaceans and the US Supreme Court: A review of cetacean mitigation and litigation in the US. Mar. Pollut. Bull. 2011, 63, 40–48. [Google Scholar] [CrossRef]
- Stevens, P.E.; Hill, H.M.; Bruck, J.N. Cetacean Acoustic Welfare in Wild and Managed-Care Settings: Gaps and Opportunities. Animals 2021, 11, 3312. [Google Scholar] [CrossRef]
- Southall, B.L.; Nowacek, D.P.; Bowles, A.E.; Senigaglia, V.; Bejder, L.; Tyack, P.L. Marine Mammal Noise Exposure Criteria: Assessing the Severity of Marine Mammal Behavioral Responses to Human Noise. Aquat. Mamm. 2021, 47, 421–464. [Google Scholar] [CrossRef]
- Montagu, G. Description of a species of Delphinus, which appears to be new. Mem. Wernerian Nat. Hist. Soc. 1821, 3, 75–82. [Google Scholar]
- Rutz, C.; Loretto, M.-C.; Bates, A.E.; Davidson, S.C.; Duarte, C.M.; Jetz, W.; Johnson, M.; Kato, A.; Kays, R.; Mueller, T. COVID-19 lockdown allows researchers to quantify the effects of human activity on wildlife. Nat. Ecol. Evol. 2020, 4, 1156–1159. [Google Scholar] [CrossRef]
- Rolland, R.M.; Parks, S.E.; Hunt, K.E.; Castellote, M.; Corkeron, P.J.; Nowacek, D.P.; Wasser, S.K.; Kraus, S.D. Evidence that ship noise increases stress in right whales. Proc. R. Soc. B Biol. Sci. 2012, 279, 2363–2368. [Google Scholar] [CrossRef] [Green Version]
- Linnaeus, C.v. Systema Naturae, 12th ed.; Laurentius Salvius: Stockholm, Sweden, 1758; Volume 1. [Google Scholar]
- Zambrano-Monserrate, M.A.; Ruano, M.A.; Sanchez-Alcalde, L. Indirect effects of COVID-19 on the environment. Sci. Total Environ. 2020, 728, 138813. [Google Scholar] [CrossRef]
- Ihsan, Y.N.; Purba, N.P.; Faizal, I.; Anya, A.; Mulyani, P.G.; Anwar, S.K. Impact of the Pandemic COVID-19 to the Indonesia Seas. Geo J. Tour. Geosites 2022, 40, 30–36. [Google Scholar] [CrossRef]
- Longden, E.G.; Gillespie, D.; Mann, D.; McHugh, K.A.; Rycyk, A.M.; Wells, R.; Tyack, P.L. Comparison of the marine soundscape before and during the COVID-19 pandemic in dolphin habitat in Sarasota Bay, FL. J. Acoust. Soc. Am. 2022, 152, A24. [Google Scholar] [CrossRef]
- Miraglia, N.; Di Brita, A. Behavior of Wildlife Species in Urban Areas to Changing Conditions during COVID-19 Lockdowns: A Review. J. Appl. Anim. Welf. Sci. 2022, 25, 119–125. [Google Scholar] [CrossRef]
- Sharma, P.; Kaur, M.; Narwal, G. Other side of the COVID-19 Pandemic: A review. Pharma Innov. 2020, 9, 366–369. [Google Scholar]
- de Blainville, H.M.D. Prodrome d’une nouvelle distribution sytématique du règne animal. Bull. Des Sci. Par La Société Philomatique De Paris 1816, 8, 105–124. [Google Scholar]
- Pine, M.K.; Wilson, L.; Jeffs, A.G.; McWhinnie, L.; Juanes, F.; Scuderi, A.; Radford, C.A. A Gulf in lockdown: How an enforced ban on recreational vessels increased dolphin and fish communication ranges. Glob. Chang. Biol. 2021, 27, 4839–4848. [Google Scholar] [CrossRef]
- Kragh, I.M.; McHugh, K.; Wells, R.S.; Sayigh, L.S.; Janik, V.M.; Tyack, P.L.; Jensen, F.H. Signal-specific amplitude adjustment to noise in common bottlenose dolphins (Tursiops truncatus). J. Exp. Biol. 2019, 222, jeb216606. [Google Scholar] [CrossRef]
- Murphy, M.M.; Jeyaseelan, S.M.; Howitt, C.; Greaves, N.; Harewood, H.; Quimby, K.R.; Sobers, N.; Landis, R.C.; Rocke, K.; Hambleton, I.R. COVID-19 containment in the Caribbean: The experience of small island developing states. Res. Glob. 2020, 2, 100019. [Google Scholar] [CrossRef]
- Shahid, M.A.H.; Sarker, M.S.A.; Hasan, M.M.; Nasrin, T. COVID-19 Pandemic Lockdown: Be a Reference for Global Environment. IJRASET 2020, 8, 459–466. [Google Scholar] [CrossRef]
- Tyack, P.; Miksis-Olds, J.; Ausubel, J.; Urban, E., Jr. Measuring ambient ocean sound during the COVID-19 pandemic. EOS 2021, 102, 155447. [Google Scholar] [CrossRef]
- Thomson, D.J.; Barclay, D.R. Real-time observations of the impact of COVID-19 on underwater noise. J. Acoust. Soc. Am. 2020, 147, 3390–3396. [Google Scholar] [CrossRef] [PubMed]
- Au, W.W.L.; Hastings, M.C. Auditory Systems of Marine Animals. In Principles of Marine Bioacoustics; Springer: New York, NY, USA, 2008; pp. 227–282. [Google Scholar]
- Popov, V.V.; Supin, A.Y. Analysis of auditory information in the brains of cetaceans. Neurosci. Behav. Physiol. 2007, 37, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Ketten, D. Functional analyses of whale ears: Adaptations for underwater hearing. In Proceedings of the OCEANS’94, Brest, France, 13–16 September 1994; Volume 261, pp. I/264–I/270. [Google Scholar]
- Ketten, D.R. Estimates of blast injury and acoustic trauma zones for marine mammals from underwater explosions. In Sensory Systems of Aquatic Mammals; Kastelein, R.A., Thomas, J.A., Nachtigall, P.E., Eds.; De Spil Publishers: Woerden, The Netherlands, 1995; pp. 391–407. [Google Scholar]
- Ketten, D.R. Structure and function in whale ears. Bioacoustics 1997, 8, 103–135. [Google Scholar] [CrossRef]
- Mooney, T.A.; Nachtigall, P.E.; Castellote, M.; Taylor, K.A.; Pacini, A.F.; Esteban, J.-A. Hearing pathways and directional sensitivity of the beluga whale, Delphinapterus leucas. J. Exp. Mar. Biol. Ecol. 2008, 362, 108–116. [Google Scholar] [CrossRef]
- Nachtigall, P.E.; Supin, A.Y.; Pacini, A.F.; Kastelein, R.A. Four odontocete species change hearing levels when warned of impending loud sound. Integr. Zool. 2018, 13, 160–165. [Google Scholar] [CrossRef]
- Southall, B.L.; Finneran, J.J.; Reichmuth, C.; Nachtigall, P.E.; Ketten, D.R.; Bowles, A.E.; Ellison, W.T.; Nowacek, D.P.; Tyack, P.L. Marine mammal noise exposure criteria: Updated scientific recommendations for residual hearing effects. Aquat. Mamm. 2019, 45, 125–232. [Google Scholar] [CrossRef]
- Dey, M.; Krishnaswamy, J.; Morisaka, T.; Kelkar, N. Interacting effects of vessel noise and shallow river depth elevate metabolic stress in Ganges river dolphins. Sci. Rep. 2019, 9, 15426. [Google Scholar] [CrossRef] [Green Version]
- Farmer, N.A.; Baker, K.; Zeddies, D.G.; Denes, S.L.; Noren, D.P.; Garrison, L.P.; Machernis, A.; Fougères, E.M.; Zykov, M. Population consequences of disturbance by offshore oil and gas activity for endangered sperm whales (Physeter macrocephalus). Biol. Conserv. 2018, 227, 189–204. [Google Scholar] [CrossRef]
- Southall, B.L.; Bowles, A.E.; Ellison, W.T.; Finneran, J.J.; Gentry, R.L.; Greene, C.R., Jr.; Kastak, D.; Ketten, D.R.; Miller, J.H.; Nachtigall, P.E. Structure of the noise exposure criteria. Aquat. Mamm. 2007, 33, 427. [Google Scholar] [CrossRef]
- Mann, D.; Hill-Cook, M.; Manire, C.; Greenhow, D.; Montie, E.; Powell, J.; Wells, R.; Bauer, G.; Cunningham-Smith, P.; Lingenfelser, R. Hearing loss in stranded odontocete dolphins and whales. PLoS ONE 2010, 5, e13824. [Google Scholar] [CrossRef]
- De Clerck, S.; Samarra, F.I.; Svavarsson, J.; Mouy, X.; Wensveen, P. Noise influences the acoustic behavior of killer whales, Orcinus orca, in Iceland. Proc. Meet. Acoust. 2019, 37, 040003. [Google Scholar]
- Luís, A.R.; Couchinho, M.N.; dos Santos, M.E. Changes in the acoustic behavior of resident bottlenose dolphins near operating vessels. Mar. Mammal Sci. 2014, 30, 1417–1426. [Google Scholar] [CrossRef]
- King, S.L.; Connor, R.C.; Krützen, M.; Allen, S.J. Cooperation-based concept formation in male bottlenose dolphins. Nat. Commun. 2021, 12, 2373. [Google Scholar] [CrossRef]
- Castellote, M.; Brotons, J.M.; Chicote, C.; Gazo, M.; Cerdà, M. Long-term acoustic monitoring of bottlenose dolphins, Tursiops truncatus, in marine protected areas in the Spanish Mediterranean Sea. Ocean Coast. Manag. 2015, 113, 54–66. [Google Scholar] [CrossRef]
- Kassamali-Fox, A.; Christiansen, F.; May-Collado, L.J.; Ramos, E.A.; Kaplin, B.A. Tour boats affect the activity patterns of bottlenose dolphins (Tursiops truncatus) in Bocas del Toro, Panama. PeerJ 2020, 8, e8804. [Google Scholar] [CrossRef] [Green Version]
- Papale, E.; Alonge, G.; Grammauta, R.; Ceraulo, M.; Giacoma, C.; Mazzola, S.; Buscaino, G. Year-round acoustic patterns of dolphins and interaction with anthropogenic activities in the Sicily Strait, central Mediterranean Sea. Ocean Coast. Manag. 2020, 197, 105320. [Google Scholar] [CrossRef]
- Southall, B.; Johnson, C.; Scholik, A.; Adam, T.; Harrison, J.; Hollinshead, K. US regulation of the effects of sound on marine life: NOAA’S mandates and use of scientific information. Bioacoustics 2008, 17, 275–278. [Google Scholar] [CrossRef]
- Stevens, P.E.; Bruck, J.N. Sensitization. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–3. [Google Scholar]
- Groves, P.M.; Thompson, R.F. Habituation: A dual-process theory. Psychol. Rev. 1970, 77, 419. [Google Scholar] [CrossRef]
- Martinez, E.; Stockin, K. Blunt trauma observed in a common dolphin delphinus sp. Likely caused by a vessel collision in the Hauraki Gulf, New Zealand. Pac. Conserv. Biol. 2013, 19, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Thompson, R.F.; Berry, S.D.; Rinaldi, P.C.; Berger, T.W. Habituation and the orienting reflex: The dual-process theory revisted. In The Orienting Reflex in Humans; Routledge: London, UK, 2021; pp. 21–60. [Google Scholar]
- Ellenberg, U.; Mattern, T.; Seddon, P.J. Habituation potential of yellow-eyed penguins depends on sex, character and previous experience with humans. Anim. Behav. 2009, 77, 289–296. [Google Scholar] [CrossRef]
- Vincze, E.; Papp, S.; Preiszner, B.; Seress, G.; Bókony, V.; Liker, A. Habituation to human disturbance is faster in urban than rural house sparrows. Behav. Ecol. 2016, 27, 1304–1313. [Google Scholar] [CrossRef]
- Uchida, K.; Blumstein, D.T. Habituation or sensitization? Long-term responses of yellow-bellied marmots to human disturbance. Behav. Ecol. 2021, 32, 668–678. [Google Scholar] [CrossRef]
- Stankowich, T. Ungulate flight responses to human disturbance: A review and meta-analysis. Biol. Conserv. 2008, 141, 2159–2173. [Google Scholar] [CrossRef]
- Blumstein, D.T. Attention, habituation, and antipredator behaviour: Implications for urban birds. Avian Urban Ecol. 2014, 41, 53. [Google Scholar]
- Constantine, R. Increased avoidance of swimmers by wild bottlenose dolphins (Tursiops truncatus) due to long-term exposure to swim-with-dolphin tourism. Mar. Mammal Sci. 2001, 17, 689–702. [Google Scholar] [CrossRef]
- Marcus, E.A.; Nolen, T.G.; Rankin, C.H.; Carew, T.J. Behavioral dissociation of dishabituation, sensitization, and inhibition in Aplysia. Science 1988, 241, 210–213. [Google Scholar] [CrossRef]
- Department of Marine and Ports Services. Government of Bermuda. 2018 Cruise Ship Schedule. 17 May 2018. Available online: http://www.rccbermuda.bm/Documents/Shipping_schedules/2018_Cruise_Ship_Schedule.pdf (accessed on 20 January 2023).
- Department of Marine and Ports Services. Government of Bermuda. 2021 Cruise Ship Schedule. 16 April 2021. Available online: http://rccbermuda.bm/Documents/Shipping_schedules/2021%20Cruise%20Ship%20Schedule.pdf (accessed on 20 January 2023).
- Houser, D.S.; Martin, S.; Crocker, D.E.; Finneran, J.J. Endocrine response to simulated US Navy mid-frequency sonar exposures in the bottlenose dolphin (Tursiops truncatus). J. Acoust. Soc. Am. 2020, 147, 1681–1687. [Google Scholar] [CrossRef]
- Bruck, J.N. Decades-long social memory in bottlenose dolphins. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131726. [Google Scholar] [CrossRef] [Green Version]
- Bruck, J.N.; Walmsley, S.F.; Janik, V.M. Cross-modal perception of identity by sound and taste in bottlenose dolphins. Sci. Adv. 2022, 8, eabm7684. [Google Scholar] [CrossRef]
- Audacity, T. Audacity. The Name Audacity (R) Is a Registered Trademark of Dominic Mazzoni. 2017. Available online: http://audacity.sourceforge.net (accessed on 20 July 2018).
- Fish, J.F.; Turl, C.W. Acoustic Source Levels of Four Species of Small Whales; Naval Undersea Center: Newport, RI, USA, 1976; Volume 547. [Google Scholar]
- Friard, O.; Gamba, M. BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Lammers, M.O.; Au, W.W. Directionality in the whistles of Hawaiian spinner dolphins (Stenella longirostris): A signal feature to cue direction of movement? Mar. Mammal Sci. 2003, 19, 249–264. [Google Scholar] [CrossRef]
- Jones, B.; Zapetis, M.; Samuelson, M.M.; Ridgway, S. Sounds produced by bottlenose dolphins (Tursiops): A review of the defining characteristics and acoustic criteria of the dolphin vocal repertoire. Bioacoustics 2020, 29, 399–440. [Google Scholar] [CrossRef]
- Blomqvist, C.; Amundin, M. High-frequency burst-pulse sounds in agonistic/aggressive interactions in bottlenose dolphins, Tursiops truncatus. In Echolocation in Bats and Dolphins; The University of Chicago Press: Chicago, IL, USA, 2004; pp. 425–431. [Google Scholar]
- Overstrom, N.A. Association between burst-pulse sounds and aggressive behavior in captive Atlantic bottlenosed dolphins (Tursiops truncatus). Zoo Biol. 1983, 2, 93–103. [Google Scholar] [CrossRef]
- Chambers, J.M. Software for Data Analysis: Programming with R; Springer: Berlin/Heidelberg, Germany, 2008; Volume 2. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Delignette-Muller, M.L.; Dutang, C. fitdistrplus: An R package for fitting distributions. J. Stat. Softw. 2015, 64, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Rankin, C.H.; Abrams, T.; Barry, R.J.; Bhatnagar, S.; Clayton, D.F.; Colombo, J.; Coppola, G.; Geyer, M.A.; Glanzman, D.L.; Marsland, S.; et al. Habituation revisited: An updated and revised description of the behavioral characteristics of habituation. Neurobiol. Learn. Mem. 2009, 92, 135–138. [Google Scholar] [CrossRef] [Green Version]
- Ronje, E.; Whitehead, H.; Barry, K.; Piwetz, S.; Struve, J.; Lecours, V.; Garrison, L.; Wells, R.S.; Mullin, K.D. Abundance and occurrence of common bottlenose dolphins (Tursiops truncatus) in three estuaries of the Northwestern Gulf of Mexico. Gulf Caribb. Res. 2020, 31, 18–34. [Google Scholar] [CrossRef]
- Van Beneden, P.J. Notice sur un nouveau dauphin de la Nouvelle-Zélande. Bull. Roy. Acad. Belg. 1881, 3, 877–882. [Google Scholar]
- Carome, W.; Slooten, E.; Rayment, W.; Webster, T.; Wickman, L.; Brough, T.; Dawson, S.M. A long-term shift in the summer distribution of Hector’s dolphins is correlated with an increase in cruise ship tourism. Aquat. Conserv. Mar. Freshw. Ecosyst. 2022, 32, 1660–1674. [Google Scholar] [CrossRef]
- Mooney, T.A.; Nachtigall, P.E.; Vlachos, S. Sonar-induced temporary hearing loss in dolphins. Biol. Lett. 2009, 5, 565–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filadelfo, R.; Mintz, J.; Michlovich, E.; D’Amico, A.; Tyack, P.L.; Ketten, D.R. Correlating military sonar use with beaked whale mass strandings: What do the historical data show? Aquat. Mamm. 2009, 35, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Buckstaff, K.C. Effects of watercraft noise on the acoustic behavior of bottlenose dolphins, Tursiops truncatus, in Sarasota Bay, Florida. Mar. Mammal Sci. 2004, 20, 709–725. [Google Scholar] [CrossRef]
- Caldwell, M.C.; Caldwell, D.K. Intraspecific transfer of information via the pulsed sound in captive odontocete cetaceans. In Animal Sonar Systems: Biology and Bionics; Laboratoire de Physiologie Acoustique: Jouy-de-Josas, France, 1967; pp. 879–936. [Google Scholar]
- Cuvier, G.; Valenciennes, A. Histoire naturelle des poisons. Tome dix-septième. Suite du livre dix-huitième. Cyprinoïdes 1844, 17, 487–519. [Google Scholar]
- Mills, S.C.; Beldade, R.; Henry, L.; Laverty, D.; Nedelec, S.L.; Simpson, S.D.; Radford, A.N. Hormonal and behavioural effects of motorboat noise on wild coral reef fish. Environ. Pollut. 2020, 262, 114250. [Google Scholar] [CrossRef]
- Önsal, Ç.; Yelimlieş, A.; Akçay, Ç. Aggression and multi-modal signaling in noise in a common urban songbird. Behav. Ecol. Sociobiol. 2022, 76, 102. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stevens, P.E.; Allen, V.; Bruck, J.N. A Quieter Ocean: Experimentally Derived Differences in Attentive Responses of Tursiops truncatus to Anthropogenic Noise Playbacks before and during the COVID-19-Related Anthropause. Animals 2023, 13, 1269. https://doi.org/10.3390/ani13071269
Stevens PE, Allen V, Bruck JN. A Quieter Ocean: Experimentally Derived Differences in Attentive Responses of Tursiops truncatus to Anthropogenic Noise Playbacks before and during the COVID-19-Related Anthropause. Animals. 2023; 13(7):1269. https://doi.org/10.3390/ani13071269
Chicago/Turabian StyleStevens, Paige E., Veda Allen, and Jason N. Bruck. 2023. "A Quieter Ocean: Experimentally Derived Differences in Attentive Responses of Tursiops truncatus to Anthropogenic Noise Playbacks before and during the COVID-19-Related Anthropause" Animals 13, no. 7: 1269. https://doi.org/10.3390/ani13071269