
Nutraceutical Effect of Resveratrol on the Mammary Gland: Focusing on the NF-κb /Nrf2 Signaling Pathways
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Plan
Chemicals
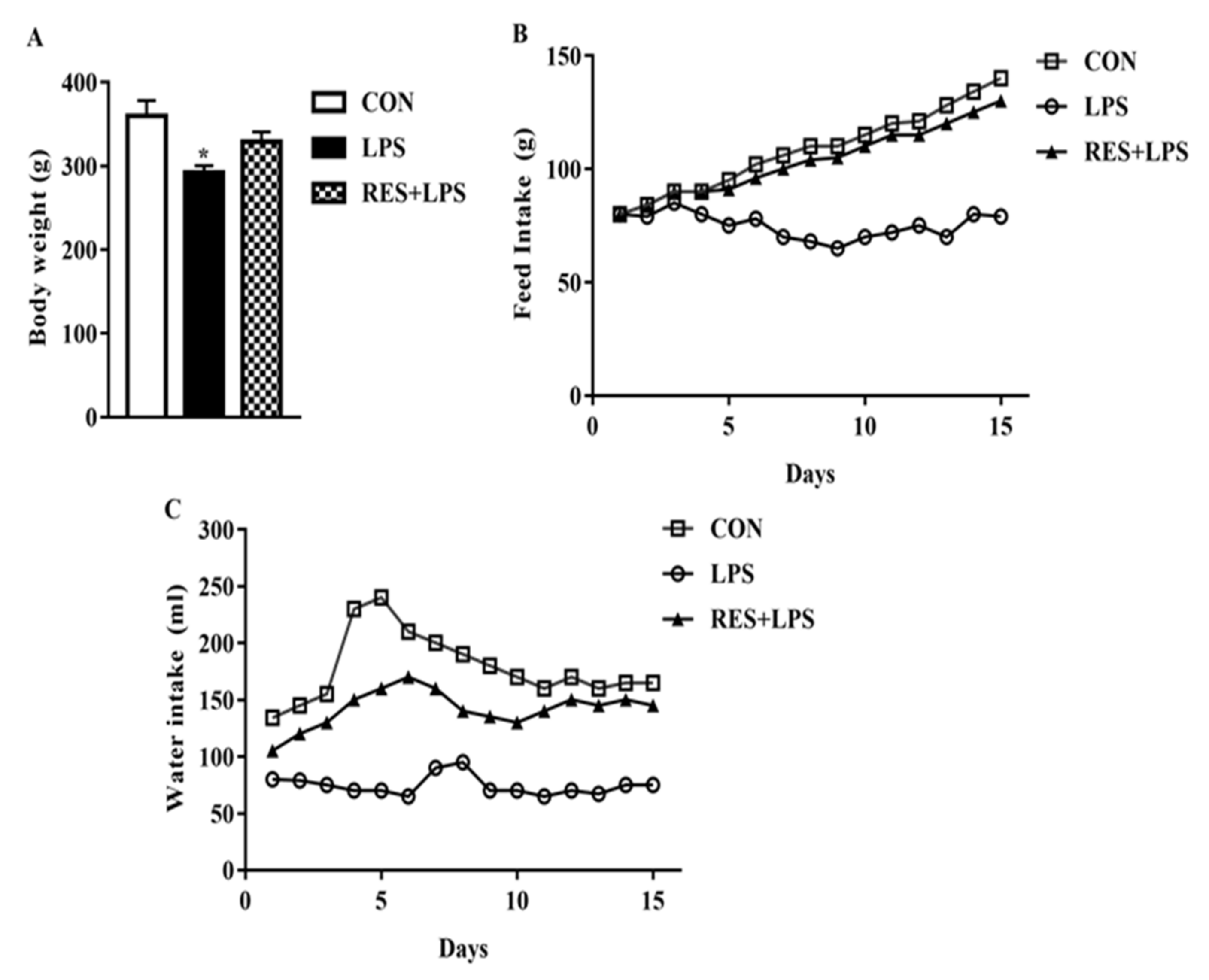
2.3. Determination of Body Weight, Feed and Water Intake in Each Group of Mice
2.4. Biochemical Analysis of Oxidative and Antioxidative Biomarkersin Mammary Tissue
2.4.1. Glutathione Peroxidase (GPX)
2.4.2. Superoxide Dismutase (SOD)
2.4.3. Catalase (CAT)
2.4.4. Malondialdehyde (MDA)
2.4.5. Determination of Total Antioxidants (T-AOC)
2.5. Radioimmunoassay
2.6. Real-Time (qRT-PCR)
Total RNA Isolation and Quantitative Real-Time PCR
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
3.1. Comparison of Anthropometric Parameters
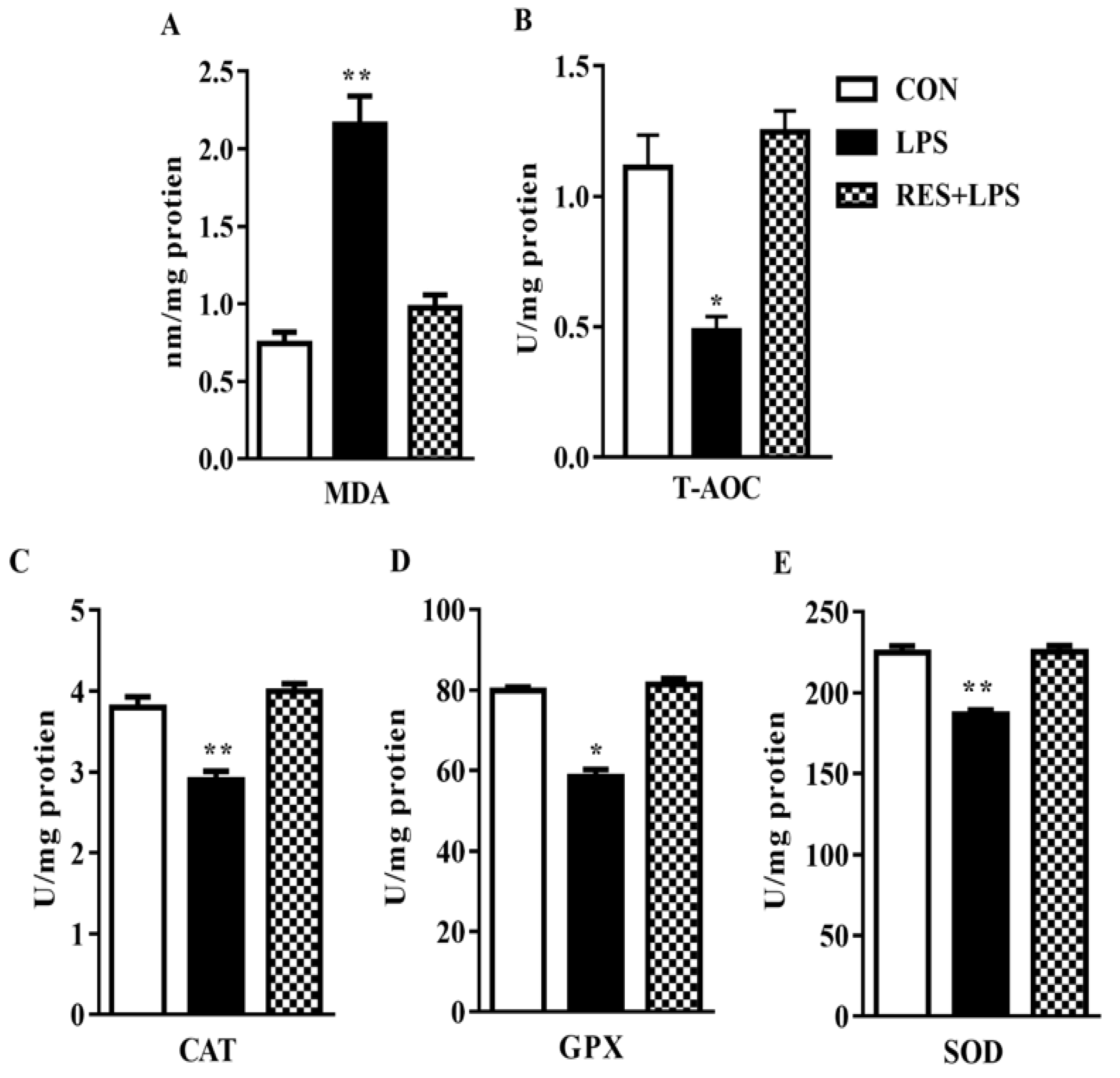
3.2. MDA Production and T-AOC Capacity in the Plasma Samples of Female Mice
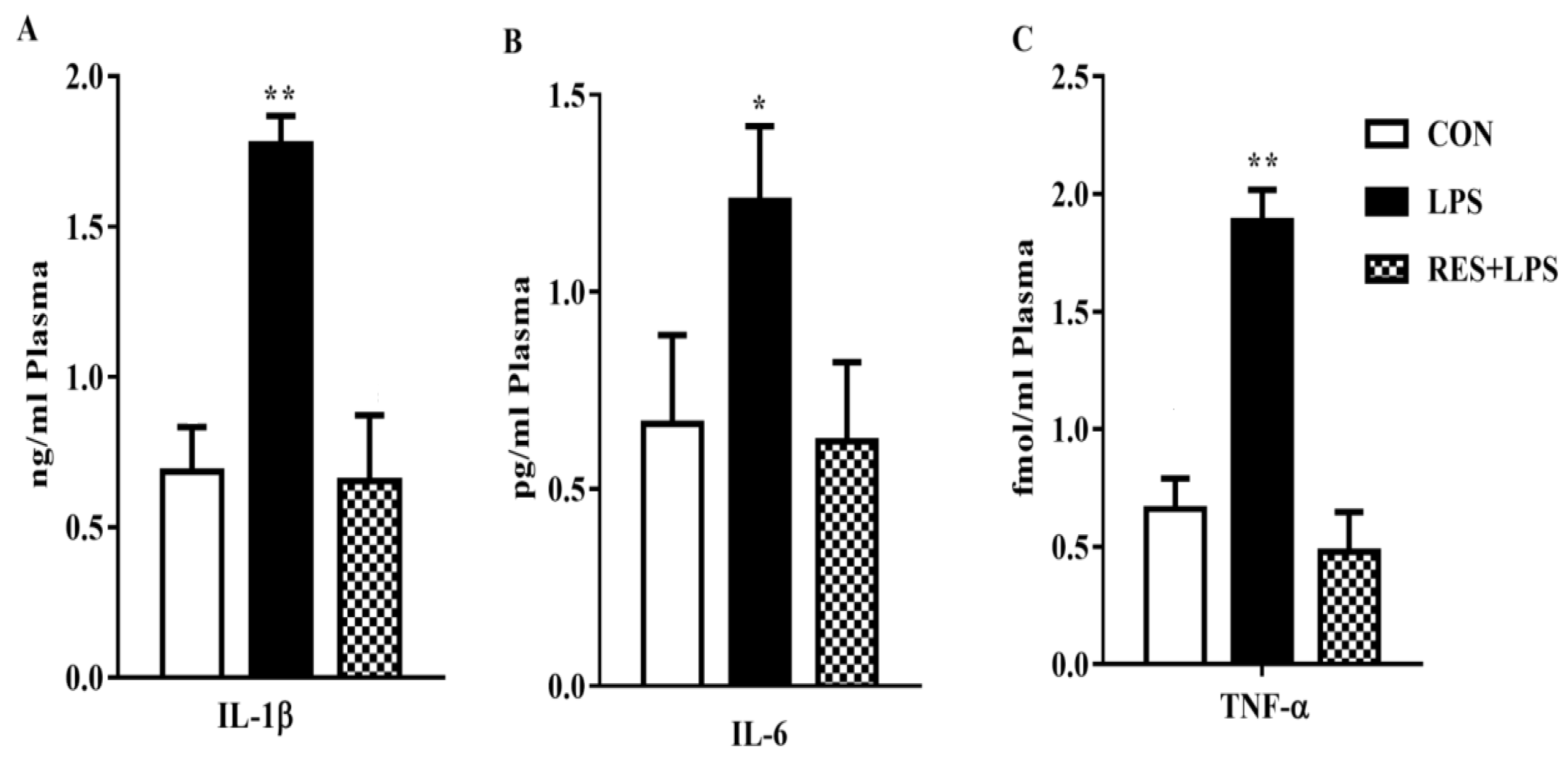
3.3. Proinflammatory Cytokines Concentration in the Plasma Samples
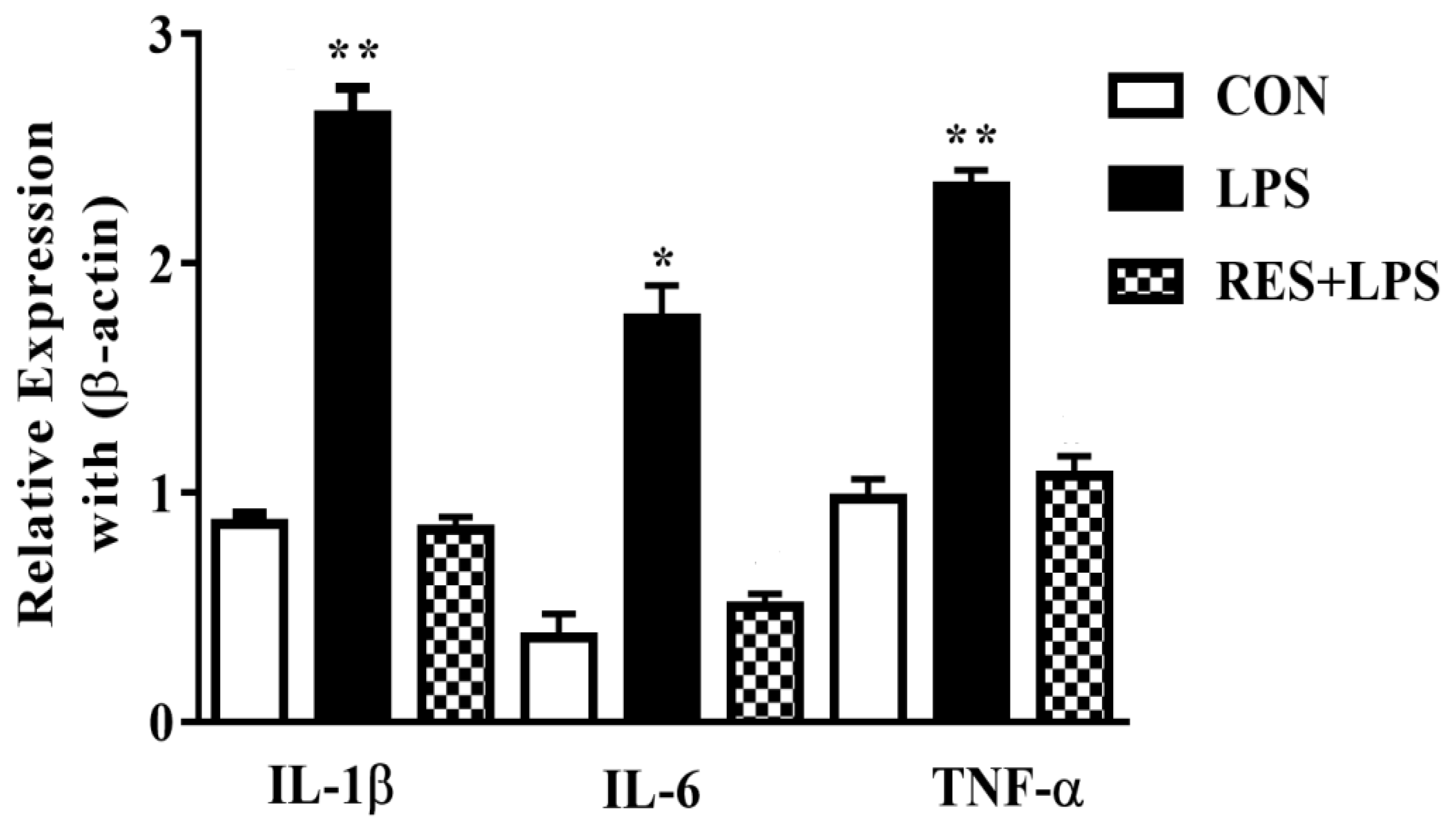
3.4. The mRNA Levels of Proinflammatory Cytokines in Mammary Tissue of Female Mice
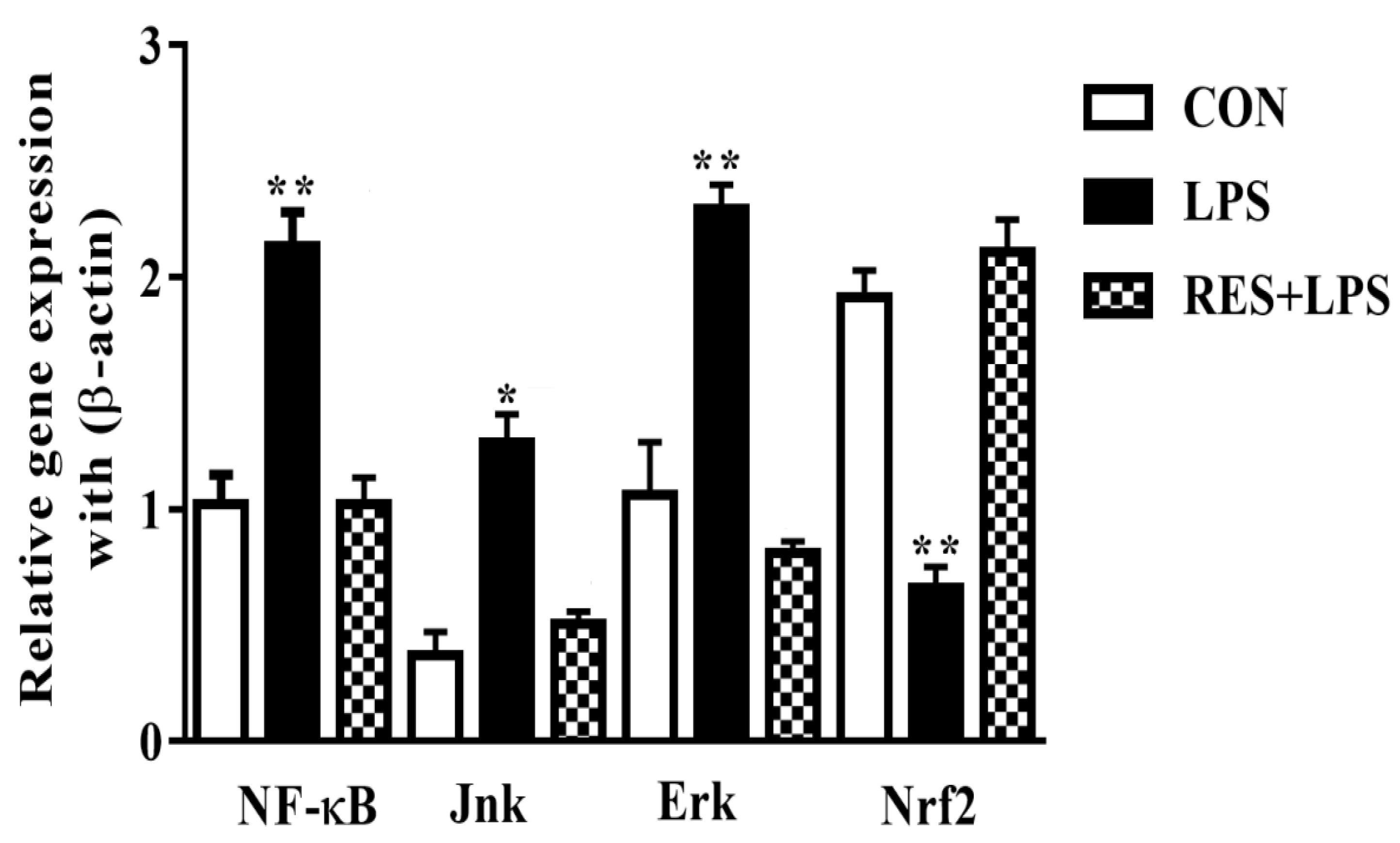
3.5. The mRNAs Expression of NF-κB, JNK, ERK and Nrf2 in Mammary Tissue of Female Mice
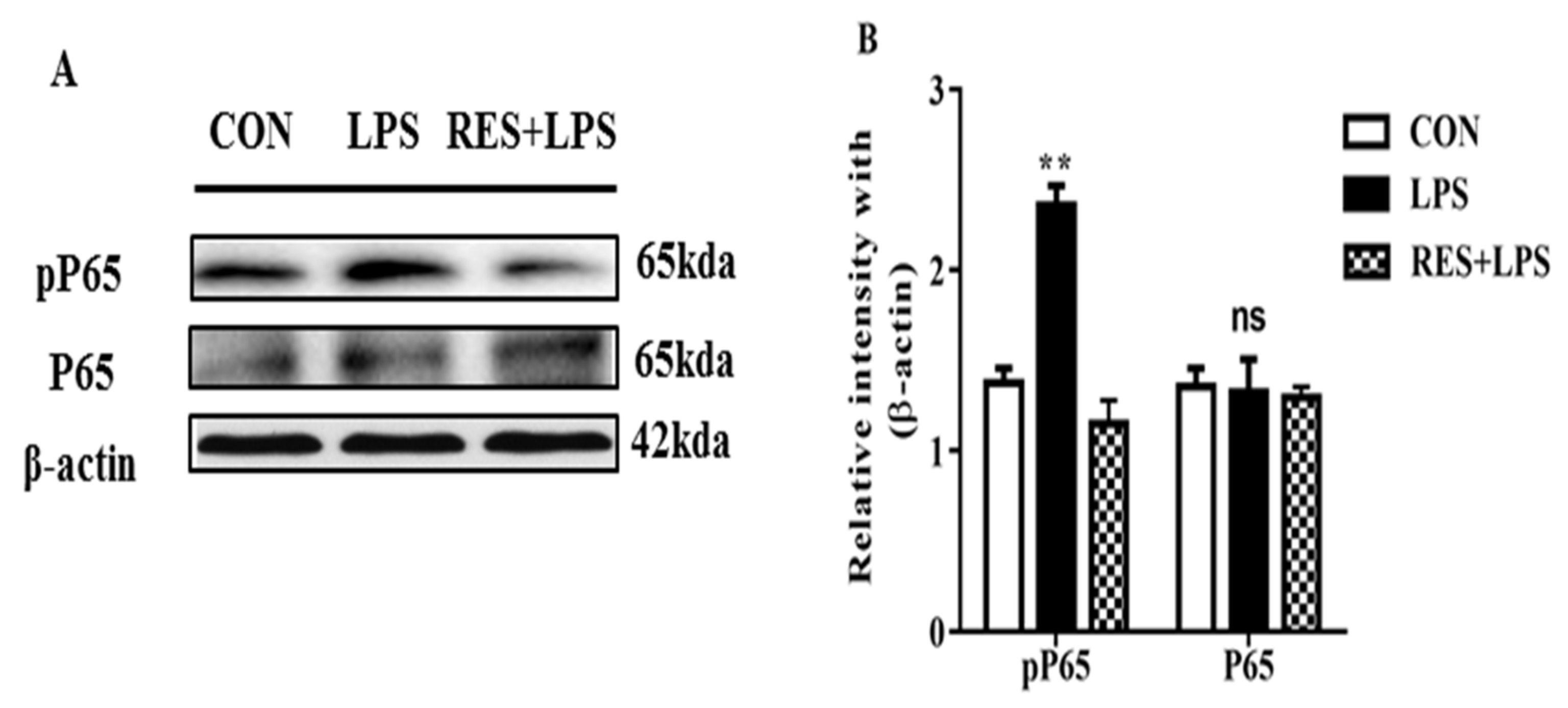
3.6. The Protein Expression of NF-κB (P65 and PP65) in Mammary Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lüderitz, O.; Galanos, C.; Rietschel, E. Lipopolysaccharides of gram-negative bacteria. In Current Topics in Membranes and Transport; Elsevier: Amsterdam, The Netherlands, 1982; pp. 79–151. [Google Scholar]
- Dai, H.; Liu, X.; Yan, J.; Aabdin, Z.U.; Bilal, M.S.; Shen, X. Sodium Butyrate Ameliorates High-Concentrate Diet-Induced Inflammation in the Rumen Epithelium of Dairy Goats. Can. J. Agric. Food Chem. 2017, 65, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Meredith, T.C.; Kahne, D. On the essentiality of lipopolysaccharide to Gram-negative bacteria. Curr. Opin. Microbiol. 2013, 16, 779–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youn, G.S.; Lee, K.W.; Choi, S.Y.; Park, J. Overexpression of HDAC6 induces pro-inflammatory responses by regulating ROS-MAPK-NF-κB/AP-1 signaling pathways in macrophages. Free Radic. Biol. Med. 2016, 97, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.S.; Abaker, J.A.; Aabdin, Z.U.; Xu, T.; Dai, H.; Zhang, K.; Liu, X.; Shen, X. Lipopolysaccharide derived from the digestive tract triggers an inflammatory response in the uterus of mid-lactating dairy cows during SARA. BMC Vet. Res. 2016, 12, 284. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.C.; Chang, G.; Ma, N.; Wang, Y.; Roy, S.; Liu, J.; Aabdin, Z.U.; Shen, X. Sodium butyrate suppresses NOD1-mediated inflammatory molecules expressed in bovine hepatocytes during iE-DAP and LPS treatment. J. Cell Physiol. 2019, 234, 19602–19620. [Google Scholar] [CrossRef]
- Cronin, J.G.; Turner, M.; Goetze, L.; Bryant, C.E.; Sheldon, I.M. Toll-like receptor 4 and MYD88-dependent signaling mechanisms of the innate immune system are essential for the response to lipopolysaccharide by epithelial and stromal cells of the bovine endometrium. Biol. Reprod. 2012, 86, 51. [Google Scholar] [CrossRef]
- Handy, D.E.; Loscalzo, J. Redox regulation of mitochondrial function. Antioxid. Redox Signal. 2012, 16, 1323–1367. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.L. Oxidative Damage and Mechanism in Dairy Goats with Mastitis; Northeast Agricultural University: Harbin, China, 2007. [Google Scholar]
- Yin, B.-S.; Li, J.J.; Hu, S.H.; Cui, Y.Z.; Song, T. Study on Relationship between Free Radical Oxidative Damage and Cow Mastitis. Prog. Vet. Med. 2011, 10, 14. [Google Scholar]
- Grant, C.M. Metabolic reconfiguration is a regulated response to oxidative stress. J. Biol. 2008, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Drackley, J.K. Biology of dairy cows during the transition period: The final frontier? J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef]
- Swaisgood, H.E. Enzymes indigenous to bovine milk. In Handbook of Milk Composition; Academic Press: New York, NY, USA, 1995; pp. 472–476. [Google Scholar]
- Abd Ellah, M. Role of free radicals and antioxidants in mastitis. J. Adv. Vet. Res. 2013, 3, 1–7. [Google Scholar]
- Braun, S.; Hanselmann, C.; Gassmann, M.G.; Keller, U.A.D.; Berclaz, C.B.; Chen, K.; Ken, Y.W.; Werner, S. Nrf2 transcription factor, a novel target of keratinocyte growth factor action which regulates gene expression and inflammation in the healing skin wound. Mol. Cell. Biol. 2002, 22, 5492–5505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.-L.; Dodd, G.; Thomas, S.; Zhang, X.; Wasserman, M.A.; Rovin, B.H.; Kunsch, C. Activation of Nrf2/ARE pathway protects endothelial cells from oxidant injury and inhibits inflammatory gene expression. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1862–H1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Hasegawa, S.; Takagi, T.; Wang, F.U.; et al. The relationship between Helicobacter pylori infection and promoter polymorphism of the Nrf2 gene in chronic gastritis. Int. J. Mol. Med. 2007, 19, 143–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, N.; Abaker, J.A.; Bilal, M.S.; Dai, H.; Shen, X. Sodium butyrate improves antioxidant stability in sub-acute ruminal acidosis in dairy goats. BMC Vet. Res. 2018, 14, 275. [Google Scholar] [CrossRef]
- Kobashigawa, J.A.; Katznelson, S.; Laks, H.; Johnson, J.A.; Yeatman, L.; Wang, X.M.; Chia, D.; Terasaki, P.I.; Sabad, A.; Cogert, G.E. Effect of pravastatin on outcomes after cardiac transplantation. N. Engl. J. Med. 1995, 333, 621–627. [Google Scholar] [CrossRef]
- Cai, W.; Zhang, L.; Song, Y.; Zhang, B.; Cui, X.; Hu, G.; Fang, J. 3,4,4′-Trihydroxy-trans-stilbene, an analogue of resveratrol, is a potent antioxidant and cytotoxic agent. Free Radic. Res. 2011, 45, 1379–1387. [Google Scholar] [CrossRef]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lluch, G.L.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Bereswill, S.; Muñoz, M.; Fischer, A.; Plickert, R.; Haag, L.M.; Otto, B.; Kühl, A.A.; Loddenkemper, C.; Göbel, U.B.; Heimesaat, M.M. Anti-inflammatory effects of resveratrol, curcumin and simvastatin in acute small intestinal inflammation. PLoS ONE 2010, 5, e15099. [Google Scholar] [CrossRef] [Green Version]
- Aabdin, U.Z.; Bilal, M.S.; Dai, H.; Abaker, J.A.; Liu, X.; Benazir, S.; Yan, J.; Shen, X. NOD1/NF-kappaB signaling pathway inhibited by sodium butyrate in the mammary gland of lactating goats during sub-acute ruminal acidosis. Microb. Pathog. 2018, 122, 58–62. [Google Scholar] [CrossRef]
- Larrosa, M.; Yañéz-Gascón, M.J.; Selma, M.V.; González-Sarrías, A.; Toti, S.; Cerón, J.J.; Tomás-Barberán, F.; Dolara, P.; Espín, J.C. Effect of a low dose of dietary resveratrol on colon microbiota, inflammation and tissue damage in a DSS-induced colitis rat model. J. Agric. Food Chem. 2009, 57, 2211–2220. [Google Scholar] [CrossRef]
- Ma, Z.H.; Ma, Q.Y.; Wang, L.C.; Sha, H.C.; Wu, S.L.; Zhang, M. Effect of resveratrol on peritoneal macrophages in rats with severe acute pancreatitis. Inflamm. Res. 2005, 54, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Elmali, N.; Baysal, O.; Harma, A.; Esenkaya, I.; Mizrak, B. Effects of resveratrol in inflammatory arthritis. Inflammation 2007, 30, 1–6. [Google Scholar] [CrossRef]
- Jarlhelt, I.; Genster, N.; Kirketerp-Møller, N.; Skjoedt, M.O.; Garred, P. The ficolin response to LPS challenge in mice. Mol. Immunol. 2019, 108, 121–127. [Google Scholar] [CrossRef]
- Mustafa, S.; Wei, Q.; Ennab, W.; Lv, Z.; Nazar, K.; Siyal, F.A.; Rodeni, S.; Kavita, N.M.X.; Shi, F. Resveratrol ameliorates testicular histopathology of mice exposed to restraint stress. Animals 2019, 9, 743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehfooz, A.; Wei, Q.; Zheng, K.; Fadlalla, M.B.; Maltasic, G.; Shi, F. Protective roles of Rutin against restraint stress on spermatogenesis in testes of adult mice. Tissue Cell 2018, 50, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, L.J.; Zhu, F.X.; Zhu, J.Y.; Chen, X.D.; Zou, L.; Saito, M. In vitro and in vivo studies on the antioxidant activities of the aqueous extracts of Douchi (a traditional Chinese salt-fermented soybean food). Food Chem. 2008, 107, 1421–1428. [Google Scholar] [CrossRef]
- Chen, H.; Ma, N.; Song, X.; Wei, G.; Zhang, H.; Liu, J.; Shen, X.; Zhuge, X.; Chang, G. Protective Effects of N-Acetylcysteine on Lipopolysaccharide-Induced Respiratory Inflammation and Oxidative Stress. Antioxidants 2022, 11, 879. [Google Scholar] [CrossRef]
- Huang, J.; Liu, J.; Chang, G.; Wang, Y.; Ma, N.; Roy, A.C.; Shen, X. Glutamine Supplementation Attenuates the Inflammation Caused by LPS-Induced Acute Lung Injury in Mice by Regulating the TLR4/MAPK Signaling Pathway. Inflammation 2021, 44, 2180–2192. [Google Scholar] [CrossRef]
- Lu, H.; Lei, X.; Zhang, Q. Moderate activation of IKK2-NF-kB in unstressed adult mouse liver induces cytoprotective genes and lipogenesis without apparent signs of inflammation or fibrosis. BMC Gastroenterol. 2015, 15, 94. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kwon, D.J.; Ju, S.M.; Youn, G.S.; Choi, S.Y.; Park, J. Suppression of iNOS and COX-2 expression by flavokawain A via blockade of NF-κB and AP-1 activation in RAW 264.7 macrophages. Food Chem. Toxicol. 2013, 58, 479–486. [Google Scholar] [CrossRef]
- Turk, R.; Piras, C.; Kovačić, M.; Samardžija, M.; Ahmed, H.; De-Canio, M.; Urbani, A.; Meštrić, Z.F.; Soggiu, A.; Bonizzi, L.; et al. Proteomics of inflammatory and oxidative stress response in cows with subclinical and clinical mastitis. J. Proteom. 2012, 75, 4412–4428. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Deng, S.; Yu, K.; Jiang, W.; Li, Y.; Wang, S.; Deng, Z.; Yao, Y.; Zhang, B.; Liu, G.; Liu, Y.; et al. Over-expression of Toll-like receptor 2 up-regulates heme oxygenase-1 expression and decreases oxidative injury in dairy goats. J. Anim. Sci. Biotechnol. 2017, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Min, B.; Ahn, D. Mechanism of lipid peroxidation in meat and meat products—A review. Food Sci. Biotechnol. 2005, 14, 152–163. [Google Scholar]
- Bouwstra, R.J.; Nielen, M.; Stegeman, J.A.; Dobbelaar, P.; Newbold, J.R.; Jansen, E.H.; Van-Werven, T. Vitamin E supplementation during the dry period in dairy cattle. Part I: Adverse effect on incidence of mastitis postpartum in a double-blind randomized field trial. J. Dairy Sci. 2010, 93, 5684–5695. [Google Scholar] [CrossRef] [Green Version]
- Abaker, J.; Xu, T.; Jin, D.; Chang, G.; Zhang, K.; Shen, X. Lipopolysaccharide derived from the digestive tract provokes oxidative stress in the liver of dairy cows fed a high-grain diet. J. Dairy Sci. 2017, 100, 666–678. [Google Scholar] [CrossRef] [Green Version]
- Sohn, M.-J.; Hur, G.M.; Byun, H.S.; Kim, W.G. Cyclo (dehydrohistidyl-l-tryptophyl) inhibits nitric oxide production by preventing the dimerization of inducible nitric oxide synthase. Biochem. Pharmacol. 2008, 75, 923–930. [Google Scholar] [CrossRef]
- Zhang, K.; Chang, G.; Xu, T.; Xu, L.; Guo, J.; Jin, D.; Shen, X. Lipopolysaccharide derived from the digestive tract activates inflammatory gene expression and inhibits casein synthesis in the mammary glands of lactating dairy cows. Oncotarget 2016, 7, 9652–9665. [Google Scholar] [CrossRef] [Green Version]
- Dong, G.; Liu, S.; Wu, Y.; Lei, C.; Zhou, J.; Zhang, S. Diet-induced bacterial immunogens in the gastrointestinal tract of dairy cows: Impacts on immunity and metabolism. Acta Vet. Scand. 2011, 53, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhao, L.; Wei, Z.; Zhang, X.; Wang, Y.; Li, F.; Fu, Y.; Liu, B. Inhibition of histone deacetylase reduces lipopolysaccharide-induced-inflammation in primary mammary epithelial cells by regulating ROS-NF-κB signaling pathways. Int. Immunopharmacol. 2018, 56, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Gwinn, M.R.; Vallyathan, V. Respiratory burst: Role in signal transduction in alveolar macrophages. J. Toxicol. Environ. Health B Crit. Rev. 2006, 9, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Brady, S.T. Post-translational modifications of tubulin: Pathways to functional diversity of microtubules. Trends Cell Biol. 2015, 25, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Knaapen, A.M.; Seiler, F.; Schilderman, P.A.; Nehls, P.; Bruch, J.; Schins, R.P.; Borm, P.J. Neutrophils cause oxidative DNA damage in alveolar epithelial cells. Free. Radic. Biol. Med. 1999, 27, 234–240. [Google Scholar] [CrossRef]
- Notebaert, S.; Demon, D.; Vanden-Berghe, T.; Vandenabeele, P.; Meyer, E. Inflammatory mediators in Escherichia coli-induced mastitis in mice. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 551–565. [Google Scholar] [CrossRef]
- MacMicking, J.; Xie, Q.-W.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.L. The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Gessner, D.K.; Fiesel, A.; Most, E.; Dinges, J.; Wen, G.; Ringseis, R.; Eder, K. Supplementation of a grape seed and grape marc meal extract decreases activities of the oxidative stress-responsive transcription factors NF-κB and Nrf2 in the duodenal mucosa of pigs. Acta Vet. Scand. 2013, 55, 18. [Google Scholar] [CrossRef] [Green Version]
- Ki, Y.W.; Park, J.H.; Lee, J.E.; Shin, I.C.; Koh, H.C. JNK and p38 MAPK regulate oxidative stress and the inflammatory response in chlorpyrifos-induced apoptosis. Toxicol. Lett. 2013, 218, 235–245. [Google Scholar] [CrossRef]
- He, X.; Wei, Z.; Zhou, E.; Chen, L.; Kou, J.; Wang, J.; Yang, Z. Baicalein attenuates inflammatory responses by suppressing TLR4 mediated NF-κB and MAPK signaling pathways in LPS-induced mastitis in mice. Int. Immunopharmacol. 2015, 28, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhou, E.; Wei, Z.; Liang, D.; Wang, W.; Wang, T.; Guo, M.; Zhang, N.; Yang, Z. Glycyrrhizin inhibits the inflammatory response in mouse mammary epithelial cells and a mouse mastitis model. FEBS J. 2014, 281, 2543–2557. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malik, M.U.U.H.; Hashmi, N.; Khan, M.; Aabdin, Z.u.; Sami, R.; Aljahani, A.H.; Al-Eisa, R.A.; Moawadh, M.S.; Algehainy, N.A. Nutraceutical Effect of Resveratrol on the Mammary Gland: Focusing on the NF-κb /Nrf2 Signaling Pathways. Animals 2023, 13, 1266. https://doi.org/10.3390/ani13071266
Malik MUUH, Hashmi N, Khan M, Aabdin Zu, Sami R, Aljahani AH, Al-Eisa RA, Moawadh MS, Algehainy NA. Nutraceutical Effect of Resveratrol on the Mammary Gland: Focusing on the NF-κb /Nrf2 Signaling Pathways. Animals. 2023; 13(7):1266. https://doi.org/10.3390/ani13071266
Chicago/Turabian StyleMalik, Muhammad Umair Ul Hassan, Nighat Hashmi, Marium Khan, Zain ul Aabdin, Rokayya Sami, Amani H. Aljahani, Rasha A. Al-Eisa, Mamdoh S. Moawadh, and Naseh A. Algehainy. 2023. "Nutraceutical Effect of Resveratrol on the Mammary Gland: Focusing on the NF-κb /Nrf2 Signaling Pathways" Animals 13, no. 7: 1266. https://doi.org/10.3390/ani13071266