1. Introduction
Body color is one of the most striking phenotypic characteristics of fish and plays important roles in predation, camouflage, immunity, and courtship [
1,
2]. The formation of body color is determined by combinations and distributions of different types of pigment cells [
3]. Body color can be divided into pigment color and structural color according to the mechanisms underlying color development [
4]. The combination of these two pigment patterns ultimately determines the color of fish [
5]. Six chromatophores have been identified in teleosts, including melanophores, erythrophores, xanthophores, iridophores, leucophores, and cyanophores [
6]. The distribution of different chromatophores leads to color differences among sites on the fish body. For example, melanophore, xanthophore, and iridophore pigment cells have been found in zebrafish [
7]. The types of pigment cells differ between the dark and light areas of this species [
8].
Fish color patterns are closely related to genetic, physiological, environmental, and dietary factors, of which genetic factors are the most critical determinants [
9]. Melanin, particularly the eumelanin, is the most widely distributed pigment in various animals, and a series of genes associated with eumelanin have been identified, such as
microphthalmia-associated transcription factor (
mitf),
melanocortin receptor 1 (
mc1r),
agouti signaling protein (
asip),
tyrosinase (
tyr),
tyrosinase-related protein 1 (
tyrp1), and
tyrosinase-related protein 2 (
tyrp2) [
10,
11]. Carotenoids often contribute to red and yellow in the skin or muscle tissues of fish. Borel et al. found that both
scavenger receptor class B member 1 (
scarb1) and
cluster determinant 36 (
cd36) are involved in carotenoid uptake and transport [
12]. In
Cyprinus carpio with
scarb1 knockout, regional red skin fades into white and the astaxanthin content is decreased [
13]. The gene
tetratricopeptide repeat domain protein 39B (
ttc39b), associated with lipid metabolism in mammals, is highly expressed in the red skin of cichlid fish [
14]. The gene
colony-stimulating factor 1 (
csf1) is associated with the maturation and development of yellow pigment cells and may be involved in the formation of yellow skin in fish [
15,
16]. The gene
apolipoprotein D (
apod) is associated with carotenoid deposition and is up-regulated in the yellow mutant rainbow trout [
17]. The
beta-carotene 15,15′-monooxygenase-1 (
bcmo1) and
beta-carotene 9’,10’-oxygenase (
bco2) genes are involved in carotenoid catabolism in mammals and fishes [
18]. Furthermore,
cytochrome P450,
family 2,
subfamily J,
polypeptide 19 (
cyp2j19) may be involved in regulating the conversion of yellow carotenoids to red ketocarotenoids in red feathers of birds [
19,
20]. Recently, Huang et al. found that
cytochrome P450,
family 2,
subfamily AE,
polypeptide 2 (
cyp2ae2) also plays an important role in ketocarotenoid accumulation in
Danio albolineatus erythrophores [
21]. However, few studies of these genes have focused on fish. In particular, genes related to the formation of red body color in fish are not well studied.
The formation of various body colors in fish is dependent on melanin, carotenoids, and pteridine. Carotenoids cannot be synthesized in fish and need to be obtained from dietary sources [
22]. Therefore, the addition of carotenoids to feed is an important strategy to promote fish coloration. Carotenoids commonly used in aquaculture include synthetic β-carotene, astaxanthin, canthaxanthin, zeaxanthin, and lutein [
23]. Astaxanthin, a keto-carotenoid, promotes coloration [
24] and improves antioxidant activity [
25] and immunity [
26]. Astaxanthin tissue deposition in fish with pink/red coloration in integument is greater than that of other carotenoids [
27]. In
Amphiprion ocellaris, feeding astaxanthin results in a more intense red color than that obtained by feeding lutein, as well as a significant increase in the skin carotenoid content [
24]. Synthetic astaxanthin and natural astaxanthin are available on the market. Synthetic astaxanthin is obtained by chemical synthesis using chemical raw materials and is a racemic mixture with an optical isomer ratio of 1:2:1 (3S,3′S:3R,3′S:3R,3′R) [
25,
28]. Natural astaxanthin is obtained from microalgae, such as
Bracteacoccus aggregatus BM5/15, yeasts
Phaffia rhodozyma, or crustaceans, such as shrimp and crabs [
29,
30,
31]. The microalgae
Haematococcus lacustris (formerly
Haematococcus pluvialis) was reported to have abundant astaxanthin [
32]. Astaxanthin of
H. lacustris exists mainly in esterified form, with a levorotatory structure (3S,3’S) in the spin isomer. The stability and antioxidant activity of astaxanthin extracted from this species were higher than those of synthetic astaxanthin [
25,
33]. Therefore,
H. lacustris is one of the best sources of natural astaxanthin. It is worth noting that the uptake and utilization efficiency of esterified/free astaxanthin by fish may vary according to species. For instance, relatively little tissue deposition of natural astaxanthin has been observed in salmon [
34]. In contrast, natural astaxanthin showed better effects on color than those of synthetic astaxanthin in
Cyprinus carpio,
Carassius auratus, and
Pseudochromis fridmani, although both improved fish color [
35,
36].
The leopard coralgrouper (
Plectropomus leopardus) is a coral reef fish with a bright red body color, luster, and blue spots. Its skin redness is one of the most important economic traits determining its market value [
37,
38]. Astaxanthin has been shown to be more effective in coloring
P. leopardus than other carotenoid sources, such as spirulina [
39,
40]. However, little is known about the effects of different sources of astaxanthin on coloration in
P. leopardus. In the present study, we compared the effects of natural astaxanthin and synthetic astaxanthin on the growth and pigmentation of
P. leopardus. Meanwhile, we used transcriptomic and metabolomic approaches to investigate the candidate genes and metabolites that may be involved in the formation of body color, providing a theoretical basis for the mechanism of coloration in
P. leopardus.
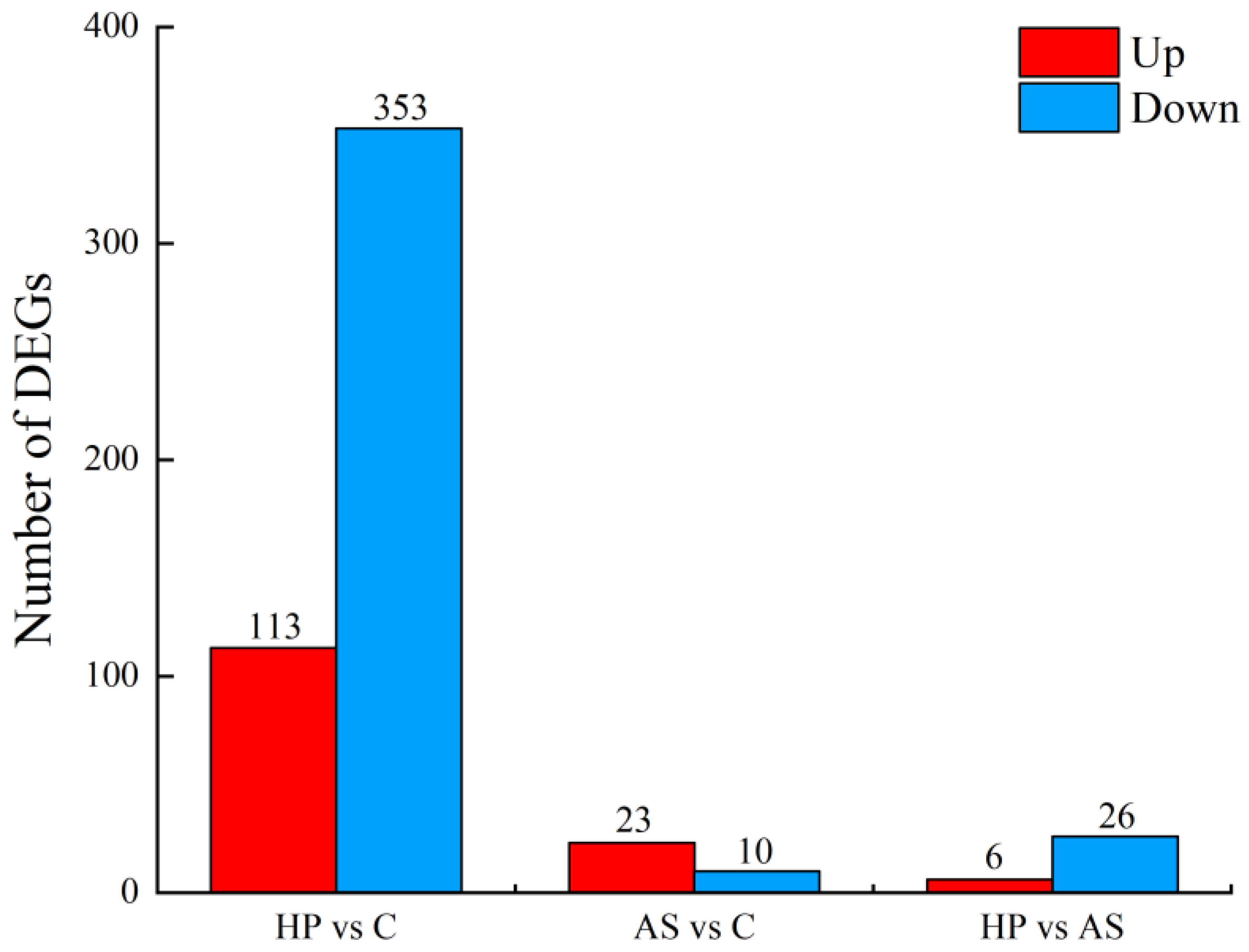
4. Discussion
The role of carotenoids in promoting growth in fish is controversial. Zhu et al. [
26] found no significant increase in the growth performance of
P. leopardus after treatment with different concentrations of astaxanthin. Micah et al. [
46] found that astaxanthin had no significant effect on blood parrotfish growth. At week 14, WGR and SGR of fish in the HP group were significantly higher than those in the C and AS groups. This suggests that natural astaxanthin exerted a beneficial effect on the growth of
P. leopardus, which is similar to results for other species, such as
Salmo salar [
47],
Pagrus pagrus [
48], and
Pseudosciaena crocea [
49]. DEGs between the HP and C group were involved in energy-metabolism-related pathways, such as sulfur metabolism and oxidative phosphorylation. In particular,
cytochrome c oxidase subunit 7A2 (
cox7a2),
NADH:ubiquinone oxidoreductase subunit B3 (
ndufb3), and
selenium-binding protein 1 (
selenbp1) were down-regulated; lipid metabolism-related pathways were also enriched. These results suggest that
P. leopardus may have a higher absorption and utilization efficiency for natural astaxanthin. Previous studies have found that astaxanthin can enhance the intermediate metabolism to promote the absorption and utilization of nutrients, thus improving the growth performance of rainbow trout [
50]. However, in our study, synthetic astaxanthin did not have a significant effect on the growth performance of
P. leopardus. Additionally, natural astaxanthin did not show a growth-promoting effect on the species in the seventh week. The growth of fish may also be related to the development stage, breeding environment, and breeding time. It is important to note that other ingredients in the supplement may also provide a supporting role in growth, even though we have maintained the consistency of the astaxanthin, protein, and fat content, respectively. Hence, the mechanism by which astaxanthin may affect the growth performance of fish needs to be further investigated.
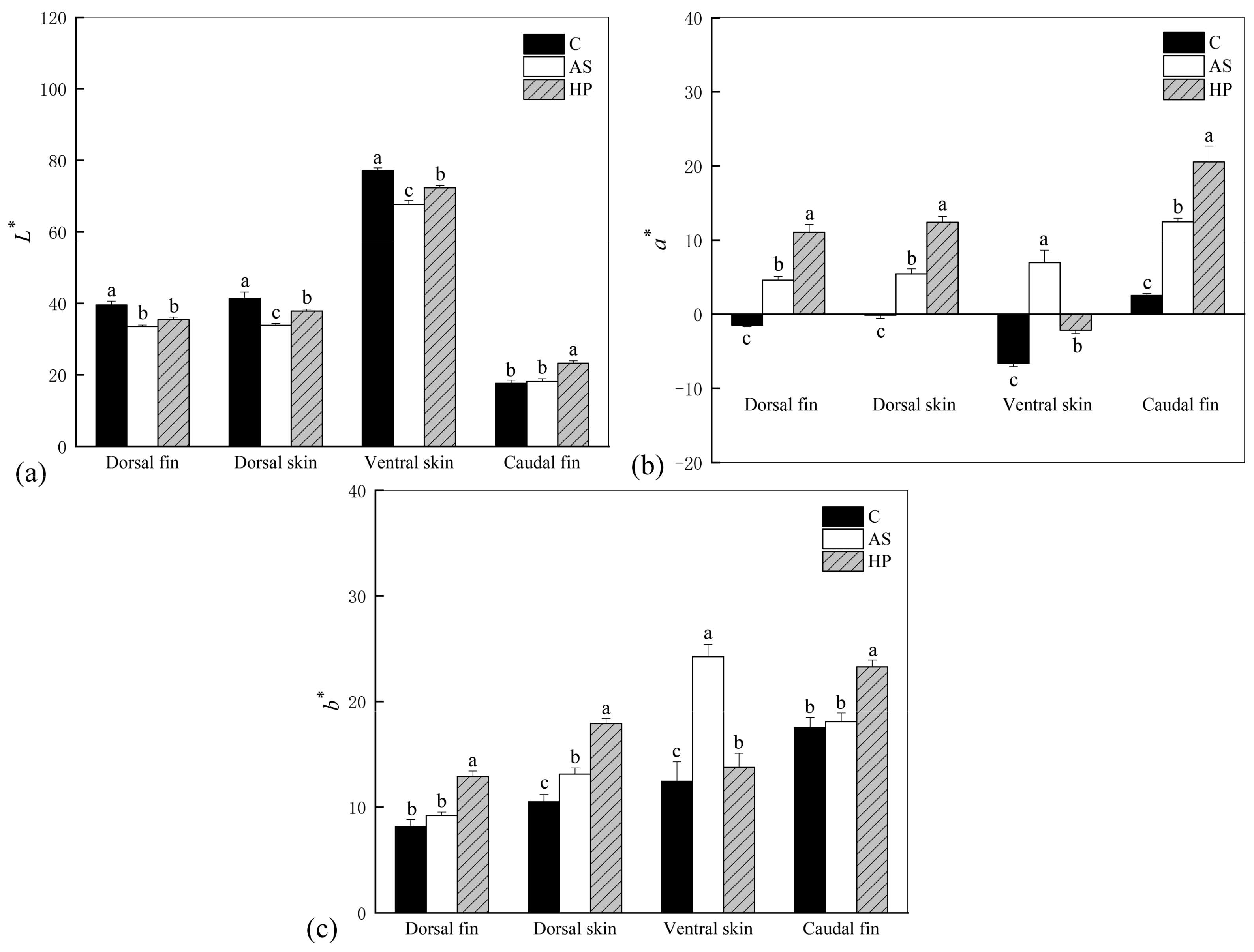
We found that astaxanthin decreased skin brightness
L*. The addition of astaxanthin to the diet effectively increased the redness
a* and yellowness
b* in
P. leopardus. The decrease in fish brightness may be related to astaxanthin. Some studies have shown that astaxanthin supplementation increased redness and yellowness but decreased brightness in blood parrotfish and
Colisa lalia [
46,
51]. This suggests that astaxanthin deposition may promote fish coloration but also lead to a reduction in brightness, resulting in a dark red appearance of the fish. Moreover, the beneficial effect of natural astaxanthin on
P. leopardus color was significantly greater than that of synthetic astaxanthin, and
a* and
b* values in the HP fish were significantly higher than those in the AS group. This may be related to the efficiency of astaxanthin digestion as well as absorption and tissue deposition. Tissue deposition in
P. leopardus may be more effective for natural astaxanthin. We also found that the body color of fish fed synthetic astaxanthin was weaker than that of the fish fed natural astaxanthin. In addition to stereochemistry, natural and synthetic astaxanthin also differ in esterification. Natural astaxanthin is mainly present in the esterified form, with a small percentage in the free state. Synthetic astaxanthin, however, is present in the non-esterified free form [
25]. Angell et al. [
52] found that the esterified astaxanthin was better absorbed than the free pigment, resulting in better coloration compared to that of the free form of synthetic astaxanthin. In
Exopalaemon carinicauda, natural astaxanthin from
H. lacustris showed better efficiency in terms of pigmentation [
53], consistent with the results of the present study. Furthermore, Maoka et al. found higher levels of total carotenoids in the red skin of
P. leopardus than in brown and black fish. The red skin of this species contained 52.7% and 28.5% of the total carotenoids in astaxanthin diesters and astaxanthin monoesters, respectively, while only 0.8% of astaxanthin diesters were detected in the black skin [
54]. This suggests that red color development
P. leopardus skin cannot be achieved without esterified astaxanthin. Li et al. found a consistency between the intensity of skin attaining a red color in blood parrots and the concentration of astaxanthin in the diet. The rate of fish attaining a red color accelerated with increasing astaxanthin in the diet. Additionally, the coloring effect of synthetic astaxanthin was better than that of natural astaxanthin [
55]. However, there was no consistency in the amount of astaxanthin added in their study, which resulted in diets with much higher levels of synthetic astaxanthin than natural astaxanthin. As they mentioned, a similar coloring effect can be obtained with natural astaxanthin if the content of natural astaxanthin is consistent with that of synthetic astaxanthin. In the experiments, more
H. lacustris powder was added to the HP diet in order to obtain a uniform astaxanthin content. However, it is unclear whether other ingredients in the supplement also contribute to fish coloration, so further research is needed.
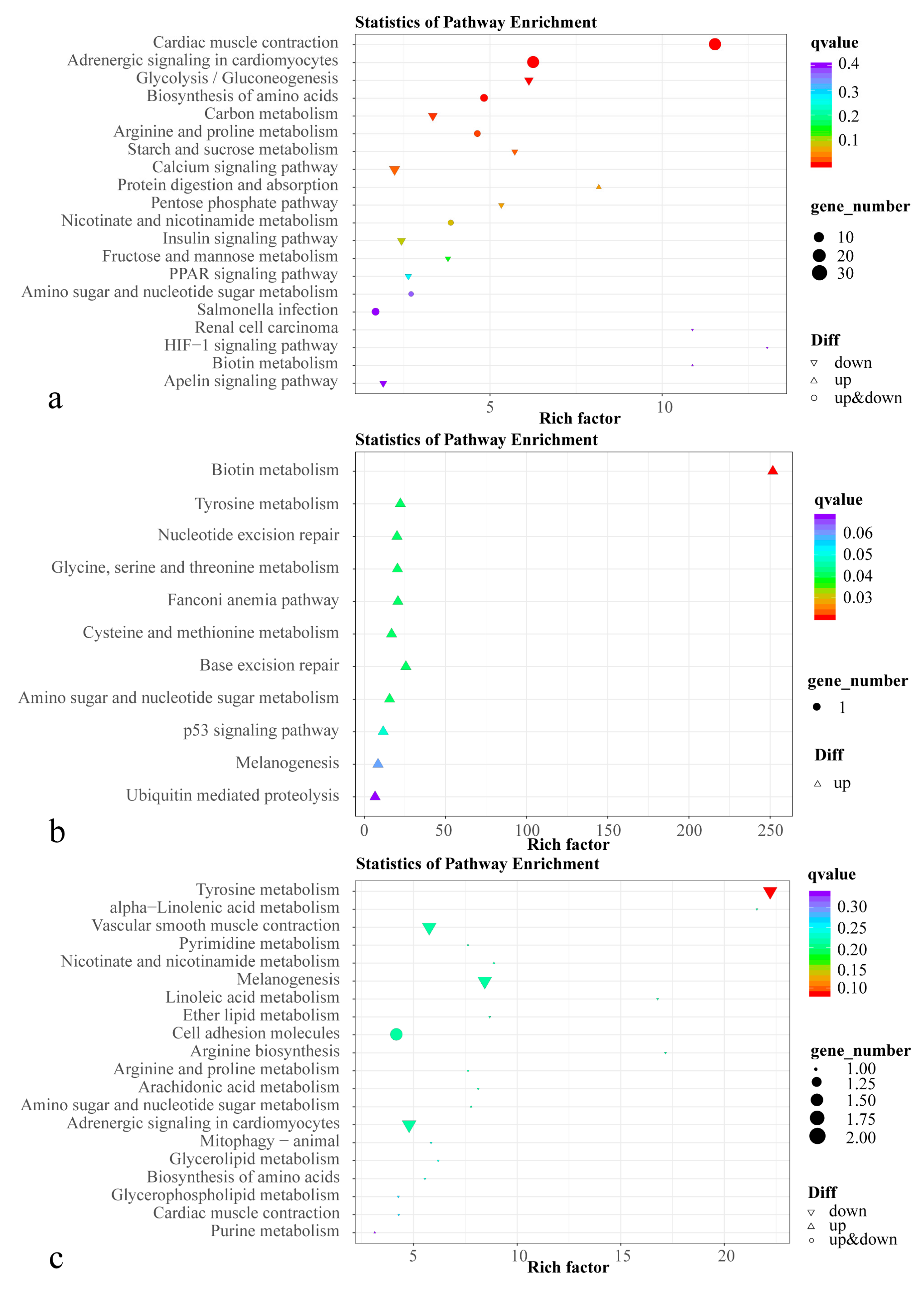
Red, orange, and yellow tissue colors in most animals are determined by carotenoids and pteridine pigments. Unlike melanin and pteridine pigments, carotenoids cannot be synthesized by fish de novo and need to be obtained in the diet [
56,
57]. As a fat-soluble substance, the carotenoid transport process is closely associated with the transport of other lipids. Binding to specific lipoproteins or apolipoproteins is necessary to transport carotenoids to specific tissues for deposition [
58,
59]. In our study, the PPAR signaling pathway, related to lipid metabolism, was significantly enriched; various genes in this pathway, including
acyl-CoA synthetase long-chain family member 1a (
acsl1a),
carnitine palmitoyl transferase 1b (
cpt1b),
fabp2, and
malic enzyme 1 (
me1), were significantly down-regulated. ACSLs (long-chain fatty acyl-CoA synthetases) are essential enzymes for the catalytic production of esteryl coenzyme A from fatty acid activation in mammals. It plays an important role in the synthesis of cholesterol, triglycerides, and phospholipids, and in fatty acid metabolism [
60]. The overexpression of this gene promotes the transport of fatty acids and the deposition of triglycerides [
61,
62,
63]. Gene
cpt1b reduces fat deposition by regulating the entry of long-chain fatty acids into mitochondria for β-oxidation [
64]. Fatty-acid-binding proteins play a key role in fatty acid uptake, intracellular transport, and metabolism [
65]. Chicken
fabp2 expression promotes fat deposition [
66]. Samulin et al. [
67] found that the co-expression of
fabp2 and
fabp1 may also have important effects on the regulation of fat deposition in pigs. me1 provides reductase for fatty acid biosynthesis coenzyme II (NADPH), which makes a positive contribution to lipid synthesis [
68]. In the present study,
acsl1a,
cpt1b,
fabp2, and
me1 were significantly down-regulated in HP vs. C, which may lead to the inhibition of fatty acid catabolic activity in fish in the HP group. Furthermore, we found that the expression of
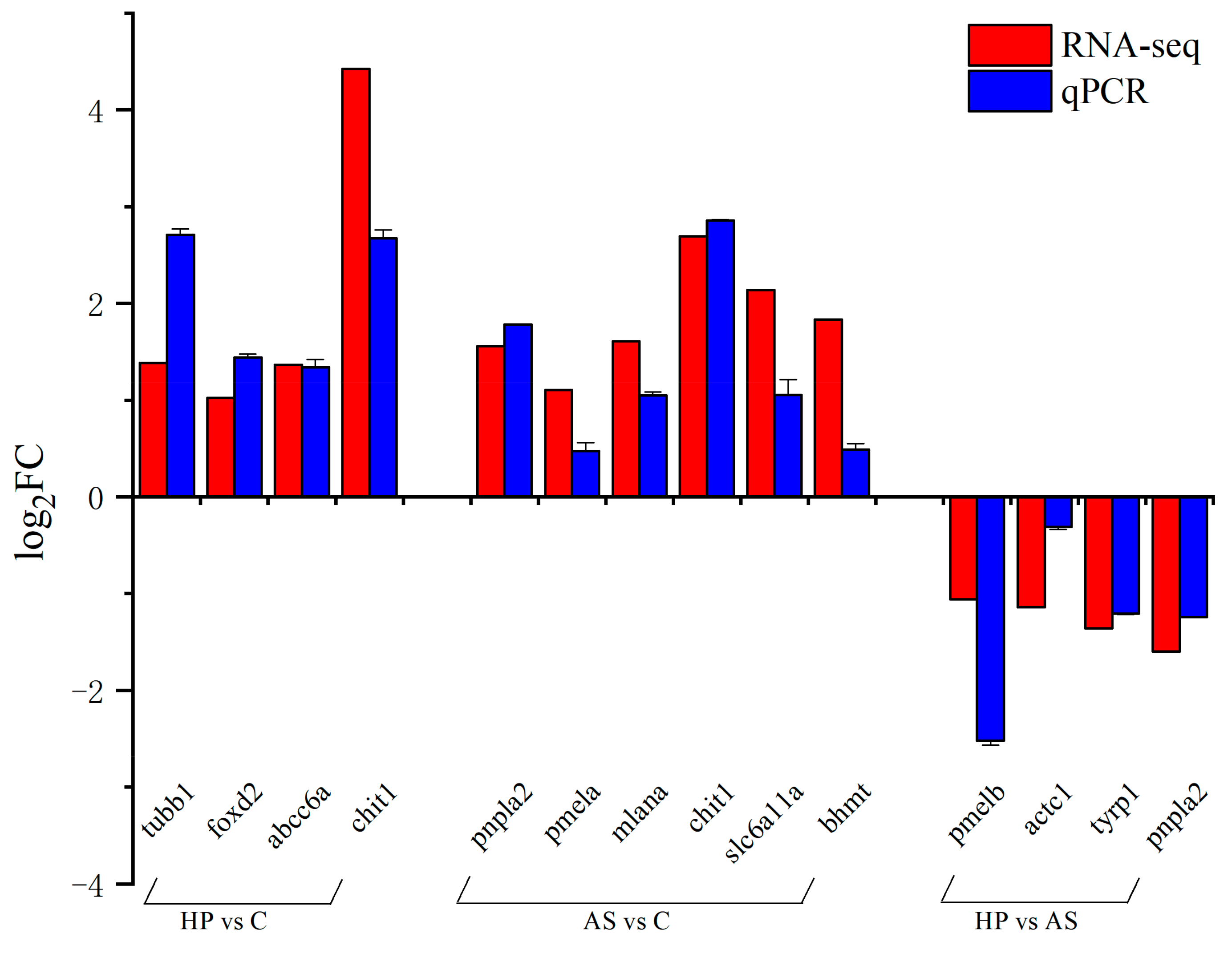
pnpla2, which is associated with lipid storage and metabolism, was down-regulated in HP vs. AS. The gene
pnpla2 encodes adipose triglyceride lipase (ATGL), which catalyzes the hydrolysis of triglyceride to diacylglycerol and free fatty acids. Studies of East African cichlids and mutant rainbow trout have reported that
pnpla2 expression is up-regulated in yellow skin. This suggests that
pnpla2 may facilitate carotenoid storage and promote yellow pigment deposition [
17,
69]. Additionally, the gene
rdh12, related to retinol metabolism using carotenoids as substrates, was down-regulated in HP vs. C, which may explain the low carotenoid substrate conversion in the fish skin.
Arachidonic acid in feed is also associated with pigmentation in fish. Diets with excessive arachidonic acid affect pigmentation in fish and can lead to albinism [
70]. In the arachidonic acid metabolic pathway of HP vs. C, we found that
Cytochrome P450 2K1 (
cyp2k1) expression was down-regulated while
phospholipase A2 group X (
pla2g10) expression was up-regulated.
Phospholipase A2 group III (
pla2g3) gene expression was down-regulated in HP vs. AS. The cytochrome CYP2 family is involved in the oxidation of fatty acids, such as arachidonic acid and lauric acid [
71]. The gene
cyp2k1 has significant fatty acid hydroxylase activity [
72].
cyp2k1 gene down-regulation in HP might inhibit fatty acid oxidation. The gene
pla2g10 catalyzes the hydrolysis of glycerophospholipids and the production of arachidonic acid [
73]. The up-regulation of
pla2g10 in the HP group may correspond to an increase in the arachidonic acid content. The gene
pla2g3 is involved in the hydrolysis of arachidonic acid on the sperm membrane in mice [
74]. We found that
pla2g3 expression was down-regulated in the HP group compared to AS, suggesting that diets supplemented with natural astaxanthin may cause the inhibition of arachidonic acid hydrolysis. Arachidonic acid is converted to prostaglandin- and thromboxane-like metabolites via cyclooxygenase. The final conversion in the liver generates the metabolite 11-dehydro-thromboxane B2. Moreover, 5(S)-HETE and leukotriene are lipoxygenase metabolites resulting from arachidonic acid metabolism [
74,
75]. Increased levels of metabolites 11-dehydro-thromboxane B2 and 5(S)-HETE implies that arachidonic acid depletion is enhanced in the HP group. We speculate that arachidonic acid may be required for color enhancement in
P. leopardus. The depletion of arachidonic acid thereby contributes to increased skin redness, similar to the results of Zhu et al. [
76].
In the present study, we found that KEGG signaling pathways associated with melanin synthesis, such as tyrosine metabolism and melanogenesis, were significantly enriched in HP compared to AS, of which
tyrp1 and
tyrp1b were significantly down-regulated. Tyrosine metabolism is associated with the synthesis of melanin, especially eumelanin. The tyrosinase protein family, mainly composed of
tyr,
tyrp1, and
tyrp2, includes key enzymes involved in the regulation of melanin synthesis. It plays an important role in melanogenesis in the skin and hair of animals. The production of albino phenotypes in vertebrates is caused by mutations or deletions in the
tyrp1 gene [
77]. Lin et al. found that the expression level of
tyrp1 was higher in black-spotted skin than in non-spotted skin in
Scatophagus argus [
78]. The gene
pmel/
silv is located downstream of the tyrosine gene family in the melanin synthesis pathway, which plays an important role in the formation of melanosomes [
79]. The knockdown of the
pmela or
pmelb gene in Nile tilapia results in a significant reduction in the number and size of melanocytes in the skin [
80]. The expression level of
creb3 was more highly expressed in HP than in C. Studies have shown that the activation of creb induces the expression of
mitf, a downstream gene. The expression of mitf can also promote the expression of the downstream genes
tyr,
tyrp1, and
tyrp2 and finally promote the generation of melanin [
81]. In the present study, the expression levels of
tyrp1 and
pmel were significantly lower in HP than in AS, suggesting that melanogenesis may be suppressed, to some extent, and that the skin was significantly redder in HP. The
tyrp1 and
pmel levels were higher in AS than in C. This suggests that the addition of synthetic astaxanthin contributed to the production of melanin in the fish. However, the redness
a* was also significantly higher in AS than in C. Therefore, further studies are needed to determine whether the inhibition of melanin synthesis or the reduction in the melanin content in the skin contributed to the redder phenotype of
P. leopardus.
In the present study, we found that the ABC transporter pathway was enriched in all three comparisons, suggesting that ABC transporters play an important role in the uptake and transport of carotenoids. Increased levels of ABC-transporter-protein-associated inosine and uridine in the HP group, compared to the control group. Inosine is involved in ATP production as a raw material for energy synthesis. It is degraded by glial cells to uric acid, which acts as a physiological antioxidant to reduce oxidative damage [
82,
83]. Similar to the metabolomic data, a transcriptome analysis revealed the significant up-regulation of
abcc6a expression in the HP group. We hypothesized that ABC transporters are involved in pigment substrates that contribute to red pigment deposition in fish. We also detected significant enrichment for glutathione metabolism and thyroid hormone synthesis pathways in both AS and HP, with the increased levels of the associated metabolite oxidized glutathione. As an endogenous antioxidant, glutathione is able to scavenge excess reactive oxygen species produced during oxidative stress. Glutathione inhibits tyrosinase activity and slows down the oxidation of L-tyrosine to form melanin, thus reducing the melanin content in the organism [
84]. This is consistent with the down-regulation of melanin-synthesis-related genes (
pmel and
tyrp1) in the HP group.
In the present study, levels of
γ-glutamyl hydrolase (
ggh) and the metabolites sepiapterin and 4a-hydroxytetrahydrobiopterin in the folate biosynthesis pathway were higher in HP than in C. The gene
ggh converts 5 methyl and 10-formyl-tetrahydrofolate forms of folic acid to the monoglutamic form in the intestine, allowing its transport through biofilms [
85]. It is noteworthy that the differentially expressed genes and metabolites in all three comparisons were enriched for folate biosynthesis pathway. The content of the metabolite sepiapterin in this pathway is increased. Sepiapterin is a yellow pteridine-like pigment involved in insect body color formation. Studies of zebrafish embryonic pteridine pigments have revealed that sepiapterin is catalyzed by sepiapterin reductase (SPR) to produce tetrahydrobiopterin (BH
4) [
86]. BH
4 can be oxidized to 4a-hydroxytetrahydrobiopterin, which can also contribute to 4a-carbinolamine dehydratase and dihydropteridine reductase reduction under the action of BH
4 [
87,
88]. This suggests that sepiapterin and 4a-hydroxytetrahydrobiopterin may be involved in the formation and deposition of red and yellow pigments in
P. leopardus. However, no DEGs related to the pteridine pathway were found in the transcriptome analysis. Therefore, the functions of these metabolites in fish still need to be further investigated.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}