1. Introduction
Welfare monitoring constitutes an indispensable part of farm animal management. Good welfare provides many benefits, including improved performance (e.g., [
1,
2,
3]) and survivability [
4] as well as fulfilling both ethical and legal obligations [
5]. The assessment of farm animal welfare has often been based on a combination of behavioural and physiological data. Behavioural parameters offer an animal-focused assessment of welfare which is particularly in keeping with one of the Five Freedoms, namely, the freedom to express normal behaviours [
6]. The behavioural measures of welfare in a commercial context are varied. For example, studies looking at the provision of alternative diets in broiler breeders used feeding motivation as a prevalent behavioural indicator of positive welfare and a healthy level of satiety [
7,
8,
9]. Other research has looked at the time budgets of behaviour in farm animals in relation to their wild-living relatives, such as in pigs or in domestic fowl [
10,
11]. Other uses of behaviour as a means to assess welfare in poultry include studies on feather pecking (e.g., [
12,
13,
14]), perching behaviour (e.g., [
15,
16,
17]) and preening (e.g., [
18,
19]).
Additionally, resting behaviour has also been used as a measure of welfare, though, often, this is as part of a broader measure of activity budgets wherein it is considered among the complete range and frequency of other common behaviours [
20,
21]. While such measures of the activity budget (and, by proxy, resting) can be useful indicators in the assessment of welfare in livestock or zoo animals, such as in the identification of stereotypic behaviour (see [
22]), there tends to be a greater focus on observable daytime activity, with little or no consideration for nighttime sleeping behaviour. While this might be due to limitations in observing animals at night, there, nevertheless, remains a considerable knowledge gap not only in our understanding of sleep behaviour as a whole but also in its greater applications in the behavioural assessment of animal welfare.
Generally, sleep can be characterised either behaviourally or electrophysiologically (by brain activity). The broader behavioural definition of sleep is outlined by five key criteria: (1) a stereotypic or species–specific sleep posture, (2) behavioural quiescence, (3) an elevated arousal threshold, (4) rapid reversibility from this state with sufficient stimulus and (5) the presence of rebound sleep following a period of sufficient deprivation [
23,
24]. When looking at sleep from an electrophysiological perspective, sleep (in mammals) is divided into two main phases: rapid eye movement (REM) sleep, also referred to as paradoxical sleep, and slow-wave sleep (SWS), also referred to as non-rapid eye movement (NREM) sleep. REM sleep is characterized by low-amplitude high-frequency brain waves, while SWS is characterized by high-amplitude low-frequency waves [
25,
26,
27]. In addition to the electrophysiological characteristics, SWS is further identified by behavioural quiescence and a reduction in muscle tone [
28]. Likewise, REM sleep is further classified by rapid eye movements, complete muscle atonia, twitching of the facial muscles and extremities and irregular heart and breathing rates [
23,
29].
Unfortunately, despite chickens being used in one of the first electroencephalogram (EEG) sleep studies [
30], the vast majority of work on sleep behaviour has been done on humans and rats (e.g., [
31,
32,
33,
34,
35,
36]), resulting in a comparatively poorer understanding of sleep in other groups of animals, including birds [
30]. Despite this, sleep in birds is surprisingly similar to that in mammals. Birds also engage in both SWS and REM sleep, although they have significantly less REM sleep compared to mammals [
37,
38]. Furthermore, the muscle atonia seen in mammals is largely restricted in birds to the neck and, occasionally, the wings [
39,
40,
41]. Birds are further unique (alongside cetaceans) in their ability to sleep uni-hemispherically, e.g., keeping one hemisphere active and the contralateral eye open at a time while the other hemisphere and eye are sleeping and closed, respectively [
29,
39,
42,
43,
44,
45,
46,
47,
48]. They can also resist sleep pressure/deprivation and forego the effects of extreme deprivation entirely, if only under specific circumstances. However, under normal circumstances, birds succumb and respond to sleep deprivation in much the same way as mammals [
49]. Specifically, wild migratory birds, such as sparrows (
Zonotrichia leucophrys gambelii) and warblers (
Sylvia Borin), have been observed to fly great distances during the night and to spend little time engaging in brief bouts of napping during the day, with seemingly no detrimental effects as a result of this deprivation. However, this resilience is only present during the migratory season [
50,
51,
52,
53]. Studies on both wild male pectoral sandpipers (
Calidris melanotos) and great frigate birds (
Fregata minor) have found that the need for sleep can be highly plastic at least for brief periods of time, such as during mating or migration [
54,
55,
56].
However, our understanding of avian sleep and the exact degree of this plasticity is unknown in domestic chickens (
Gallus gallus domesticus) and, in particular, commercially kept chickens. This is partly due to greater focus being placed on production metrics and improving performance in commercial animals rather than purely behavioural studies, such as studies looking at the behavioural time budgets of broilers in varying light conditions (e.g., [
57]), studies looking at improving reproductive success (e.g., [
58]) and/or studies looking to improve feed conversion efficiency (e.g., [
59]). The studies that have looked at sleep in chickens have either been limited to purely behavioural observations, which—while useful—are limited in describing the full breadth of sleep, or are, in the face of ever-advancing technology, out of date [
30,
60,
61,
62]. The only studies employing EEGs in order to specifically observe the sleep behaviour of poultry were those by Ookawa and Gotoh [
30], Ookawa [
60] and van Luijtelaar et al. [
63]. These seminal studies were fundamental in establishing the use of EEGs to study sleep. Their overall findings showed that chickens engage mostly in SWS, which is often interrupted by REM sleep [
30]. REM sleep is described as only occurring at night and lasting between 5 and 30 sec per bout [
30]. The few remaining papers used behavioural observations to describe the timing and duration of resting and sleeping behaviours (purely based on posture) [
64] or to describe perching behaviour and perching motivation [
65,
66]. Moreover, work by Boerema et al. [
67], using behavioural observations, found that laying hens deprived of sleep exhibited a decrease in monocular (i.e., uni-hemispheric) sleep in favour of binocular sleep. While a fair number of studies have looked at sleep behaviour in juvenile birds (e.g., [
42,
68,
69,
70]), the great discrepancy in sleep requirements and behaviour between juvenile and adult birds renders comparisons inappropriate.
It is, therefore, evident that there remains a gap when it comes to our understanding of sleep in poultry. Consequently, the aim of this study was to combine both behavioural and physiological (via EEG) observations to first establish a baseline for sleep in laying hens and to observe the short-term effects of disturbances on laying hen sleep behaviour, with the potential of using sleep as a welfare indicator in the future.
2. Materials and Methods
2.1. Animals
This study was approved by SRUC’s Animal Experiments Committee prior to beginning. A group of 19 H&N Brown Nick (brown-feathered) laying hens were acquired from a commercial flock (JSR Services Ltd.; Blairgowrie, UK) at 59 weeks of age. Hens consisted of experimental (n = 12) and companion (n = 7) birds. Three experimental birds could not be used post-surgery due to a failure in logging equipment and so are not referred to further. The remaining experimental birds made up experimental batches 1–3 (3 hens per batch). A further three hens were collected from the same commercial flock at 65 weeks of age to make up batch 4. Upon arrival, all birds were weighed, were wing-tagged with Ketchum wing tags (Putham, UK) and were placed in the holding room.
2.2. Housing
Birds were housed in two different rooms: one for general holding and one for experimental treatments. Both rooms were similar in size, had concrete floors, had LED lighting (one light above each pen) and were automatically heated and ventilated to maintain the room at 21 °C. Lights were on daily from 05:00 to 19:00 GMT (14L:10D) to reflect a standard commercial egg production environment. Pens in both rooms were made of wooden frames with an area of 2 m2 each and wire mesh siding (5 × 5 cm) that was 1 m high. A layer of fine wire mesh (1 × 1 cm) was added up to a height of 0.5 m with the pen walls above 1 m and made of plastic mesh (5 × 12 cm). Pen floors were filled with wood shavings as litter. Birds were provided with ad libitum access to layer pellets (16% crude protein and 11.5 MJ energy; ForFarmers UK Ltd., Rougham, UK) and water, approximately 1 kg of alfalfa hay used as enrichment (topped up as required), and a perch 60 cm long × 40 cm high was used.
2.2.1. Holding Room
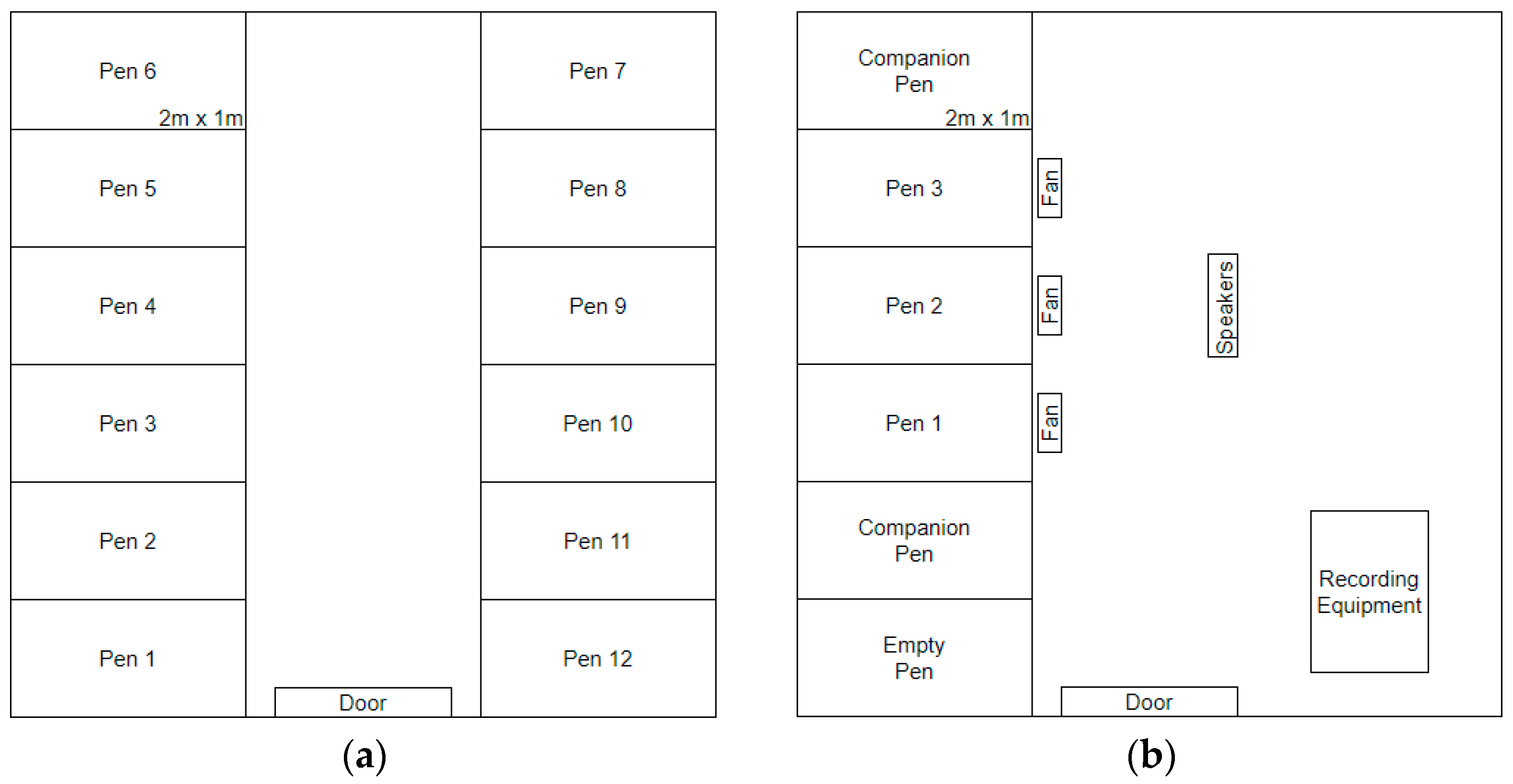
The holding room (
Figure 1a) consisted of 12 pens (6 pens on either side of the room) that were 2 m deep × 1 m wide and made with wire mesh and wood frames. Here, floor pen litter was approximately 5 cm deep, and hens also had access to nest boxes (1 nest box per pen). The first 19 hens were housed in 7 pens of 2 birds each and 5 pens of a single bird each. Birds were housed individually or in pairs based on their behaviour and signs of feather pecking to maximize welfare and comfort, and they always had visual and auditory contact with neighbouring birds. Bird weight and condition was used to determine which birds were to be used as experimental animals, with the lightest birds and those with the worst feather condition being used as companion animals. As the total number of birds in the holding room decreased, paired birds identified as experimental hens were separated into individual pens. Birds in batch 4 were added after birds 1–3 had been moved to the experimental room.
2.2.2. Experimental Room
Hens were housed in this room after EEG surgery to record their sleep behaviour. The experimental room (
Figure 1b) consisted of five pens (all on one side of the room) of identical construction to those used in the holding room. The flooring of each pen was filled with approximately 2.5 cm of wood shavings to limit sitting/sleeping in the litter and to encourage a greater amount of perching. In these pens, hens had no nest boxes to allow cameras to record all of their behaviours, and perches were 30 cm long. One companion bird was always in each of the end pens, with the three focal experimental birds occupying the three central pens (one hen/pen). Companion birds were used to ensure that all experimental birds had a conspecific on either side with whom to remain in auditory and visual contact. Companion birds were replaced after the end of batches 1 and 3 to limit their exposure to experimental stressors (
Table 1).
2.3. EEG Implants
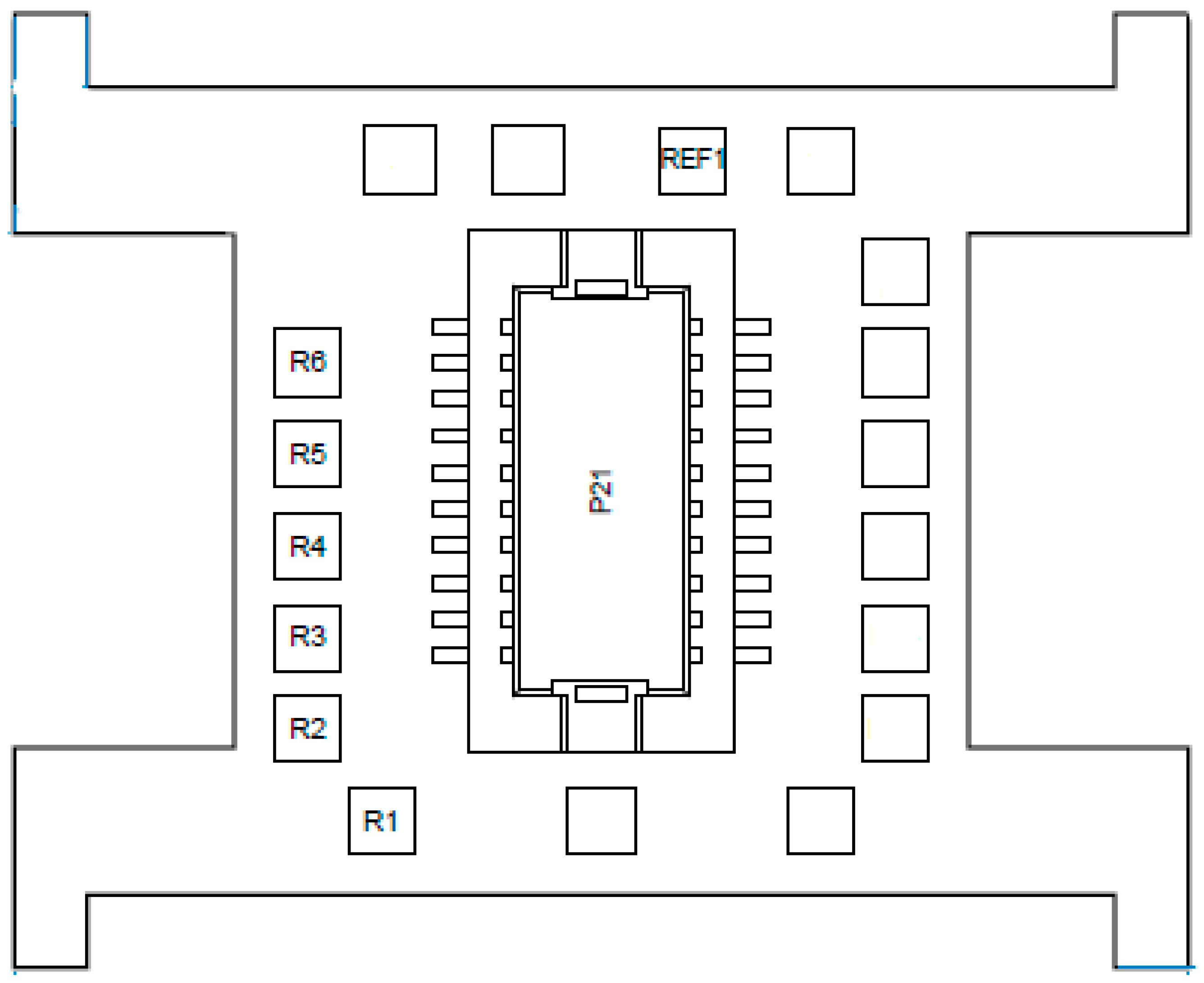
EEG implants were used to record brain activity during sleep. Implants consisted of an electronic interface board (EIB) microchip (ViewPoint Behavior Technology; Lyon, France). To transmit electrical brain activity to the chip, 0.25 mm diameter insulated wires were soldered to the EIB on one end and were soldered to a socket adaptor on the other. Six wires of 2.5 cm each were used for EEG probes, and one wire of 3.5 cm was used for the reference/baseline electrode. The number of wires used corresponded with the number of EEG channels to be used (
Figure 2). Approximately 0.5 cm of each wire was exposed and wrapped around a 3 A, 100 V, IC socket adaptor (0.40 mm diameter and 2.5 mm length) (Bürklin Elektronik; Oberhaching, Germany). This adaptor was used as the electrode for measuring brain wave activity. The wire and IC socket adaptor were soldered together using soldering flux (Farnell; Paisley, UK); a 25 W, 230 V soldering iron (Farnell; Paisley, UK) and 1 mm solder (Maplin Professional Services; Edinburgh, UK). The opposite end of the wire was stripped by approximately 0.3 cm; the wire was then fed through the bottom of the correct hole (see
Figure 3) and was unravelled. An EIB pin was positioned over the hole, and special pliers were used to press the pin into the hole to secure the wire (
Figure 3). Frayed wire ends were removed with a scalpel blade to ensure accurate signal conductance and were covered in a thin layer of dental acrylic (Kemdent; Swindon, UK) to prevent detachment of wires during surgical implantation. Probe ends were cleaned with acetone to disinfect them prior to implantation.
2.4. Surgical Preparation
All operating surfaces were made aseptic using Milton sterilising fluid (2% sodium hypochlorite in water; Milton International; Nantes, France); all surgical tools were washed and sterilised in sterilising fluid (from batch 1) and then via autoclave (from batch 3 onwards) prior to surgical procedures. Three birds (constituting one batch) were pre-selected for surgery 2–3 days prior to the surgery date. One bird (randomly selected to be first) was fasted overnight (approximately 12 h) before surgery. The remaining two birds were fasted on the surgical day starting at approximately 07:30.
2.5. Surgical Procedure
Birds were brought into the operating room one at a time, were weighed, and were placed inside a cage measuring 49 cm × 55 cm × 78 cm. Ketamine (Ketamidor, Chanelle; UK) at 5 mg/kg and butorphanol (Torbugesic, Covetrus; Portland, ME, USA) at 0.4 mg/kg were given intra-muscularly to act as a sedative and analgesic, respectively, prior to provision of anaesthesia. Anaesthesia was induced using 4–5% sevoflurane (Zoetis; Louvain-la-Neuve, Belgium) per 1 L O2 and was maintained at between 2 and 2.5%. Vital signs (i.e., pulse, body temperature and oxygen saturation) were continuously monitored during surgery; additionally, fluid levels were maintained with regular injections of saline solution via a brachial vein indwelling catheter. After confirming complete anaesthesia using comb and toe pinches, birds were placed in a stereotaxic frame, with the head secured using ear bars. Birds were placed on top of a fluid-absorbing pad to collect excreta as well as a self-heating pad and covered in a fleece blanket to maintain body temperature during surgery.
Pet clippers (Langba; Ningbo, China) were used to remove feathers on the head within an approximately 4.5 cm × 4.5 cm area; any larger remaining feathers were removed with forceps. The area was cleaned with hydrogen peroxide prior to incision. A single incision was made using a Number 10 scalpel blade (Swann–Morton; Sheffield, UK), running from approximately 0.2 cm posterior to the comb to approximately 0.5 cm anterior to the neck musculature. From here, four haemostat clamps (Surtex Instruments Ltd.; London, UK) were attached to the four corners of the incision site to hold the skin open and to expose the cranium. Dry cotton swabs were used to remove the connective tissue on top of the cranium as well as to absorb any blood. Hydrogen peroxide was used to further clean the skull and to highlight any remaining material. The cranium was then dried.
The implant methodology used by [
56] was employed. A point 1 cm anterior to the fusion lines of the cranial bones was demarcated as the point of origin using a skin marker (P
3; Bristol, UK) and was used to delimit the length of the hyperpallial region. From here, dots were placed at 0.2 cm on either side of the origin to indicate the placement of the first two EEG probes. A second point was placed 0.7 cm anterior to the origin from where dots were placed at 0.3 cm on either side. A third point 0.3 cm anterior to the origin was demarcated, and a final two dots were placed at 0.4 cm on either side. This resulted in a total of 6 EEG probes with as wide a spread across the hyperpallium as possible. A final smaller ‘X’ was placed approximately 0.5 cm towards the bottom-right corner from the origin (in the porous bone) above the cerebellum to identify where the reference probe should be placed.
The demarcated dots were then drilled to expose the dura beneath. The tip of a scalpel blade was used to score all seven of the drill sites to prevent the drill bit from slipping during drilling. A 0.5 mm drill bit fitted to a manual pin vise drill (XLYYLWB; China) was used to drill the seven holes as well as to drill several anchor holes within the porous bone of the posterior part of the skull to act as additional surface area for the dental acrylic to adhere to. The drilling of the probe holes went only as far as the dura, which could be identified by a slight change in resistance and confirmed by a difference in hue from that of the cranial bones. From here, a low-viscosity drop of dental acrylic was placed in each probe hole immediately prior to probe insertion to ensure probes did not fall out of their designated holes. Upon securing all seven probes, a higher-viscosity dental acrylic was used to create a foundation across the probe ends protruding from the probe holes and across the anchor holes. The EIB microchip was then manipulated to be at a sufficient height (approximately 1 cm vertically from the skull) to allow for the probe wires to be tucked underneath, to prevent skin irritation, and to later allow for the EEG device to sit flush against the implant. From here, a thick layer of dental acrylic was steadily built up to encase the wiring underneath and to secure the EIB microchip to the skull.
Prior to completely encasing the bottom and corners of the microchip, two small omega-shaped metal hooks bent downwards at 90 degrees (handmade from large paperclips) were placed lateral to the implant and were secured with dental acrylic. Prior to suturing, 2% lidocaine (Hameln Pharmaceuticals; Gloucester, UK) was applied topically to the skin to minimize any immediate post-operative pain. 1 suture was placed anterior to the implant, and 1–2 sutures were placed posteriorly. After completion of surgery, birds were taken off anaesthesia and were monitored for signs of wakefulness (e.g., reflex test response, movement and independent breathing). Once muscle tone was present and once birds could stand, they were placed inside a second cage (identical to that used pre-surgery) with food, water, wood shavings as litter and a heat source if the bird was slow to wake. Once birds were alert and standing, they were brought to the experimental room to recover.
2.6. Post-Surgical Recovery and Monitoring
Birds were monitored during post-operative recovery for six days as well as being habituated to the experimental pens. General behavioural observations (e.g., body posture, activity levels and egg laying) and assessment of health were used as an overall guide. Birds were weighed daily to monitor their health and to determine the correct dosage for post-operative medications. The medications given were as follows: meloxicam (Meloxidyl, Covetrus; Portland, ME, USA) (1 mg/kg) was given subcutaneously twice per day as an analgesic for a total of 4 days; enrofloxacin (Baytril, Covetrus; Portland, ME, USA) (10 mg/kg) was given subcutaneously once per day as an antibiotic for a total of 6 days and dexamethasone (Dexafast, Covetrus; Portland, ME, USA) (0.5 mg/kg) was given intra-muscularly once per day to reduce the likelihood of neurological complications for a total of 2 days. Post-operative medications were provided in the morning at approximately 09:00 with second dosing (meloxicam only) provided at approximately 16:00.
2.7. EEG Devices
EEGs were monitored and recorded using the ONIEROS device (ViewPoint; Lyon, France). The device weighed 56 g (i.e., no more than 3.4% of body weight of the lightest bird used) and measured 3 cm × 2 cm × 1.5 cm. Each device was clicked into the implant and was further secured with a rubber band using the omega-shaped hooks. Devices were charged daily. Devices were removed at 08:30 every morning during routine husbandry and charged for 1 h (maximum charging time required) after which they were re-attached. This allowed for 23 h of continuous EEG recording for each bird.
2.8. Data Recording
EEG data were collected using EphyLab software (ViewPoint; Lyon, France). Each individual EEG channel, a reference channel, and three channels for accelerometery (x, y and z axes) were collectively recorded in real time and were automatically parcelled into four-hour-long data files.
Each experimental pen was outfitted with three infrared cameras: two H.264 CCTV IMX323 night vision IR mini cameras (Ailipu Technology Co., Ltd.; Shenzhen, China) on either side of the perch and a third high-resolution varifocal camera (Twilight CCTV; Merseyside, UK) pointing downwards from the top of the pen to capture the entire enclosure in frame (
Figure 4). All nine cameras were connected to a computer with Geovision (Taipei, Taiwan) recording software installed which recorded and saved all video files in ‘.avi’ format. Nighttime recording of both EEGs and behaviour via video started at lights out (19:00) and ran until 08:30 the following day when devices were removed for charging. Daytime recording resumed at 09:30 and continued until 19:00. EEG and behavioural recordings were synchronized across devices.
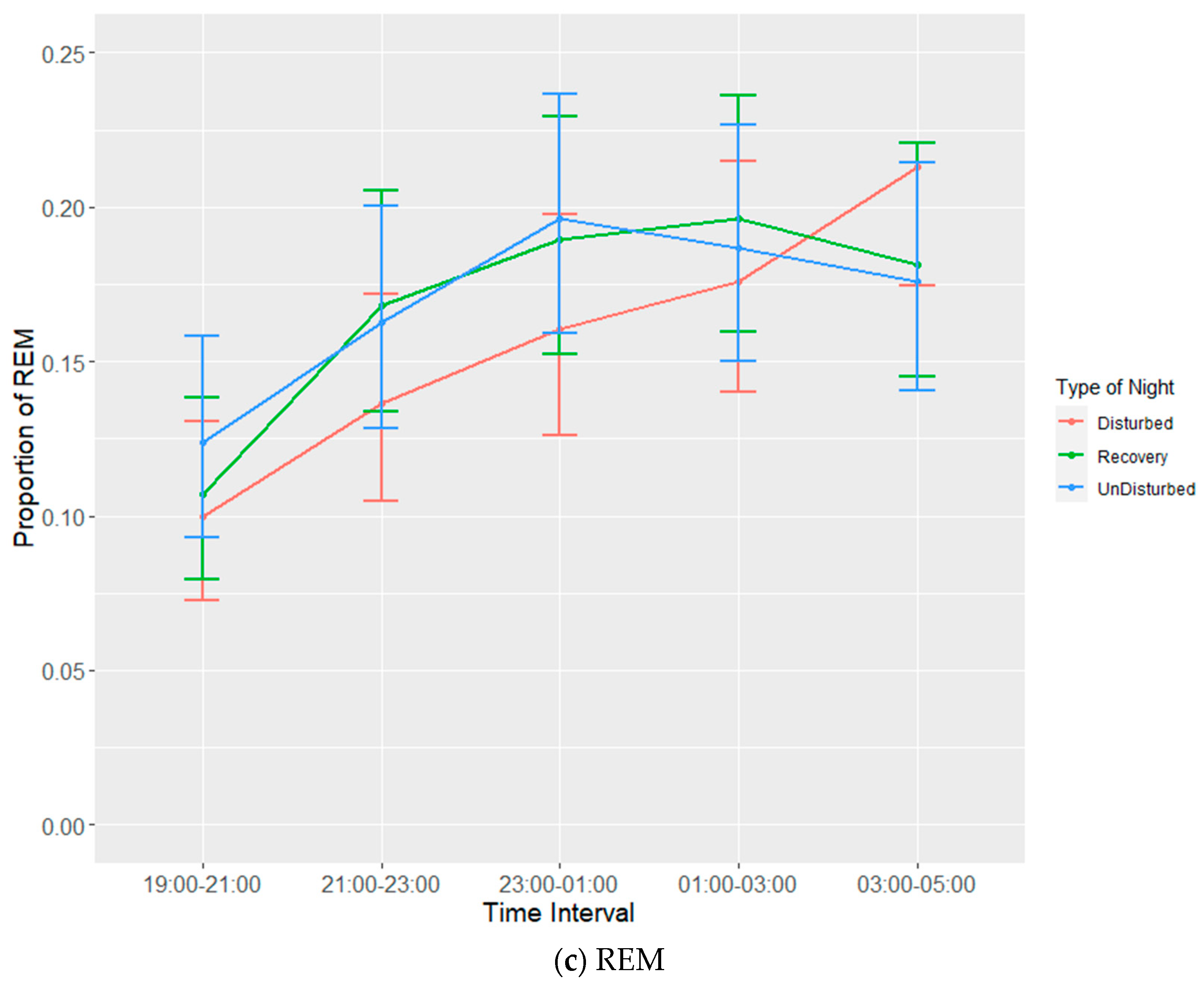
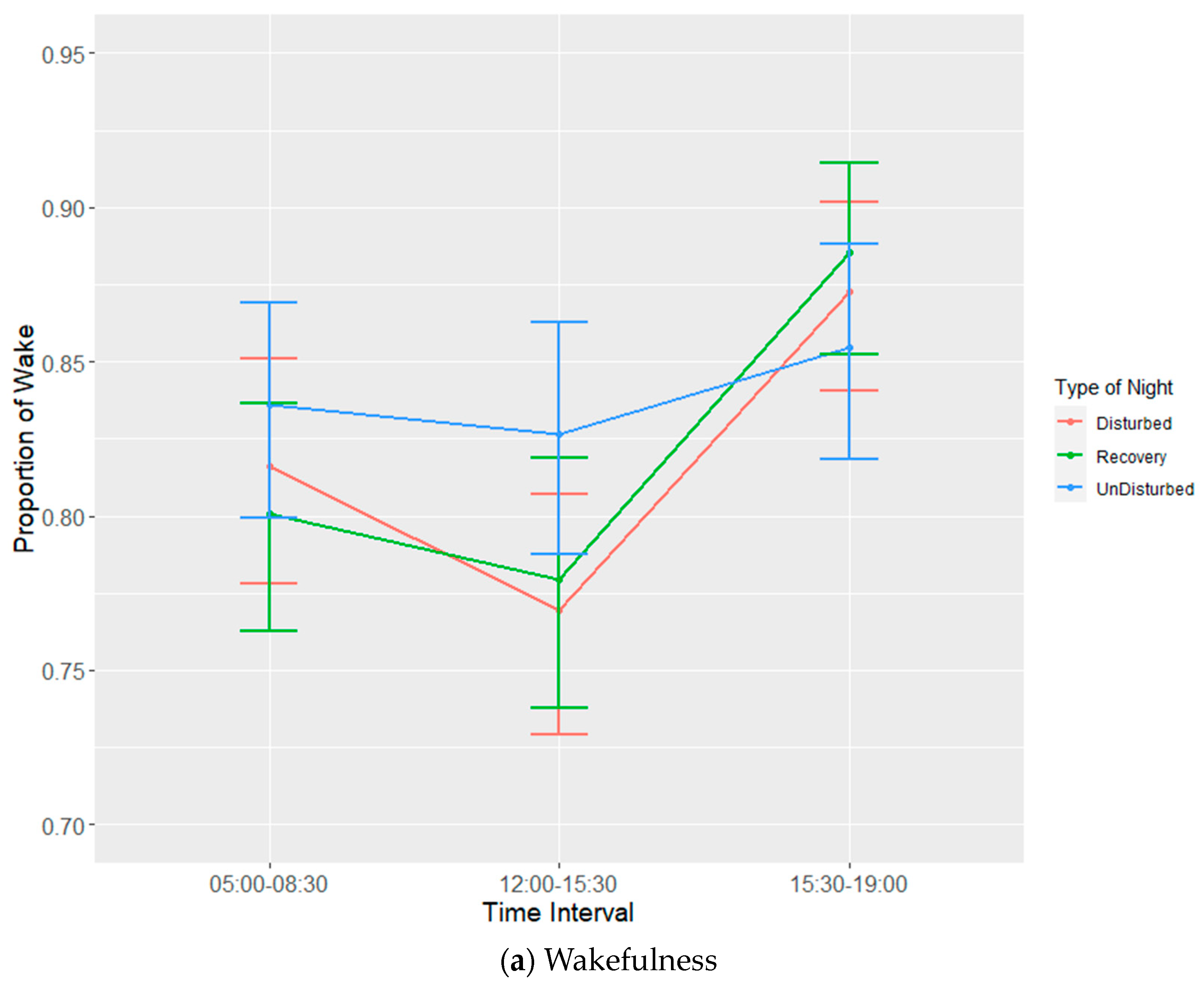
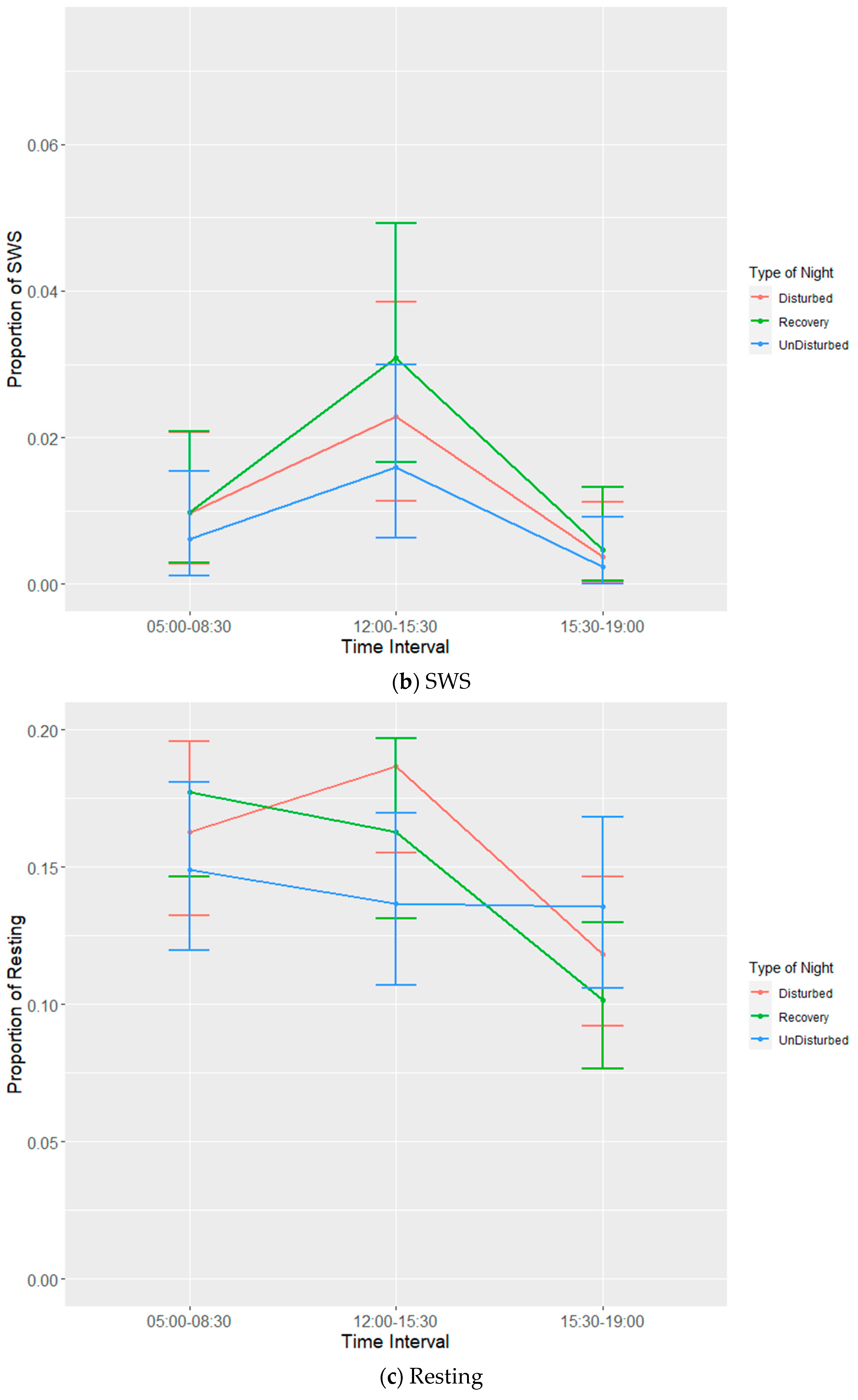
Each experimental batch of hens was recorded for six 24 h periods. Recording periods consisted of undisturbed nights, disturbed nights and recovery nights (after a disturbed night) to observe both baseline and disruption effects on bird sleep behaviour. The allocation of disruptions per day per batch are given in
Table 2. It was not possible to completely randomize the given nights, as a recovery night always had to follow a disturbed night. EEG devices would be attached to birds prior to lights out on the first night and then set to begin recording (in tandem with the three infrared cameras) at 19:00. Birds were manually placed on perches to ensure complete recording of sleep behaviours during the dark cycle. The total duration of the dark period was 10 h during which there was a 6-h span of sleep disruptors that were ordered to ensure balance (
Table 3). Disruptors were set to turn on every half hour between 21:00 and 02:30, remain on for a duration of five minutes and then turn off again. The three disruptors used were wind from a small fan, noise generated by an mp3 player playing a traffic audio file on repeat and speakers set at a volume of 90 dB and light generated by the normal operating lights set at a maximum of 20 lux. Following a disturbed night, a recovery night would involve the use of the EEG devices and cameras without any of the disruptors. Undisturbed nights also used EEG devices and cameras without any of the disruptors and were used as a means of establishing a baseline for sleep behaviour. Devices were removed on the morning of the seventh day following recording.
2.9. Data Collection and Processing
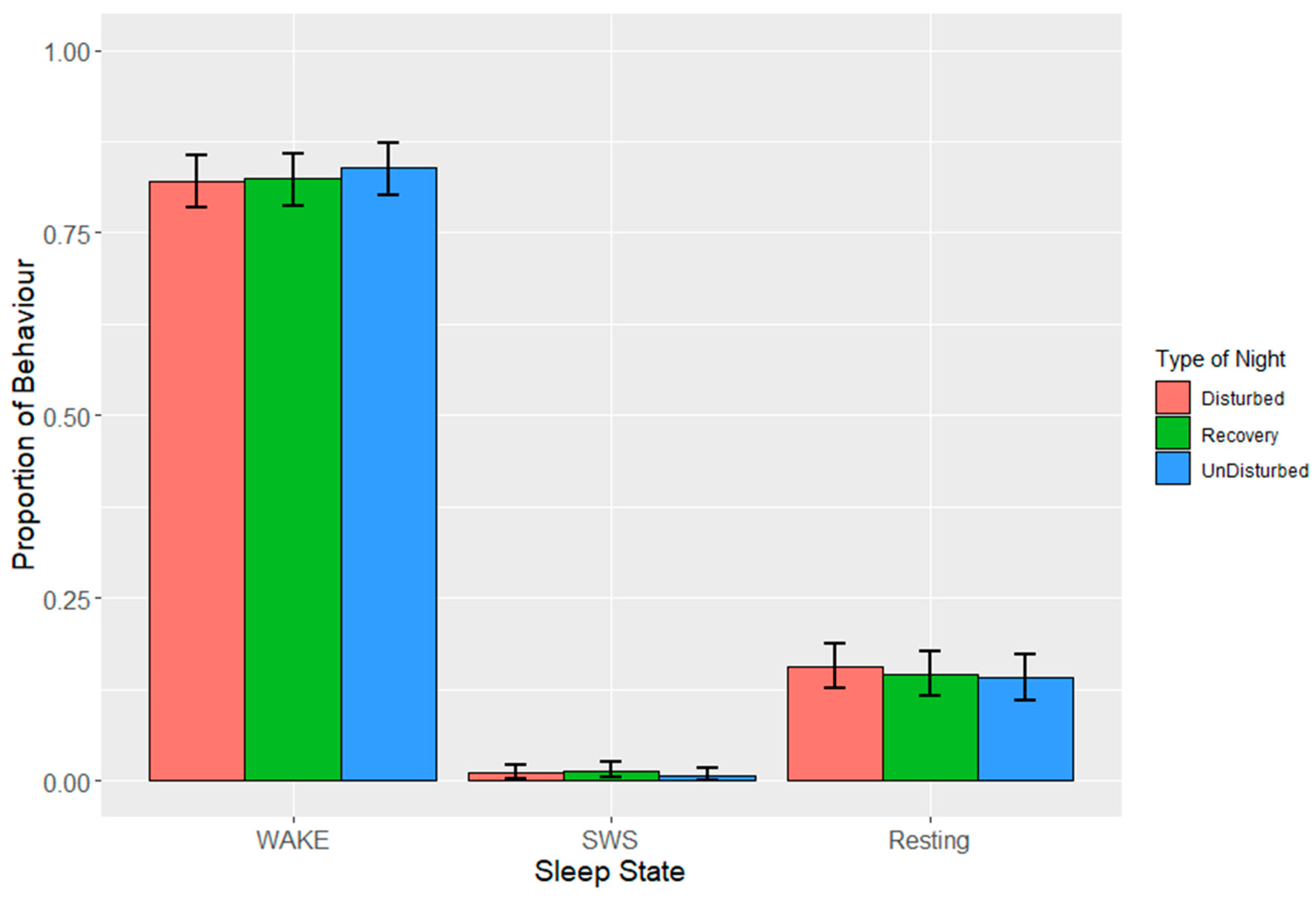
EEG data were processed using Slip Analysis (ViewPoint; Lyon, France). Individual EEG files (noted by date and time) were uploaded to the software and were merged with the corresponding video files. Slow-wave sleep (SWS), rapid eye movement sleep (REM), wakefulness, and artefacts (electronic noise) were scored at the start of every minute using an epoch length of four seconds based on the amplitude of the EEG waves. SWS is identified by the presence of high-amplitude low-frequency waves, while both wakefulness and REM sleep are identified via low-amplitude high-frequency waves. Videos of hen behaviour were used as confirmation for intermediate (or uni-hemispheric) states or states that were more difficult to define based solely on EEG data such as between wakefulness and REM sleep (
Table 4). The presence of one open eye was a clear indicator for instances of intermediate state, where wave amplitude alone was not sufficient. Additional behavioural indicators were used to discern REM sleep from wakefulness, closed eyes and drooping head being the primary indicators.
2.10. Statistical Analysis
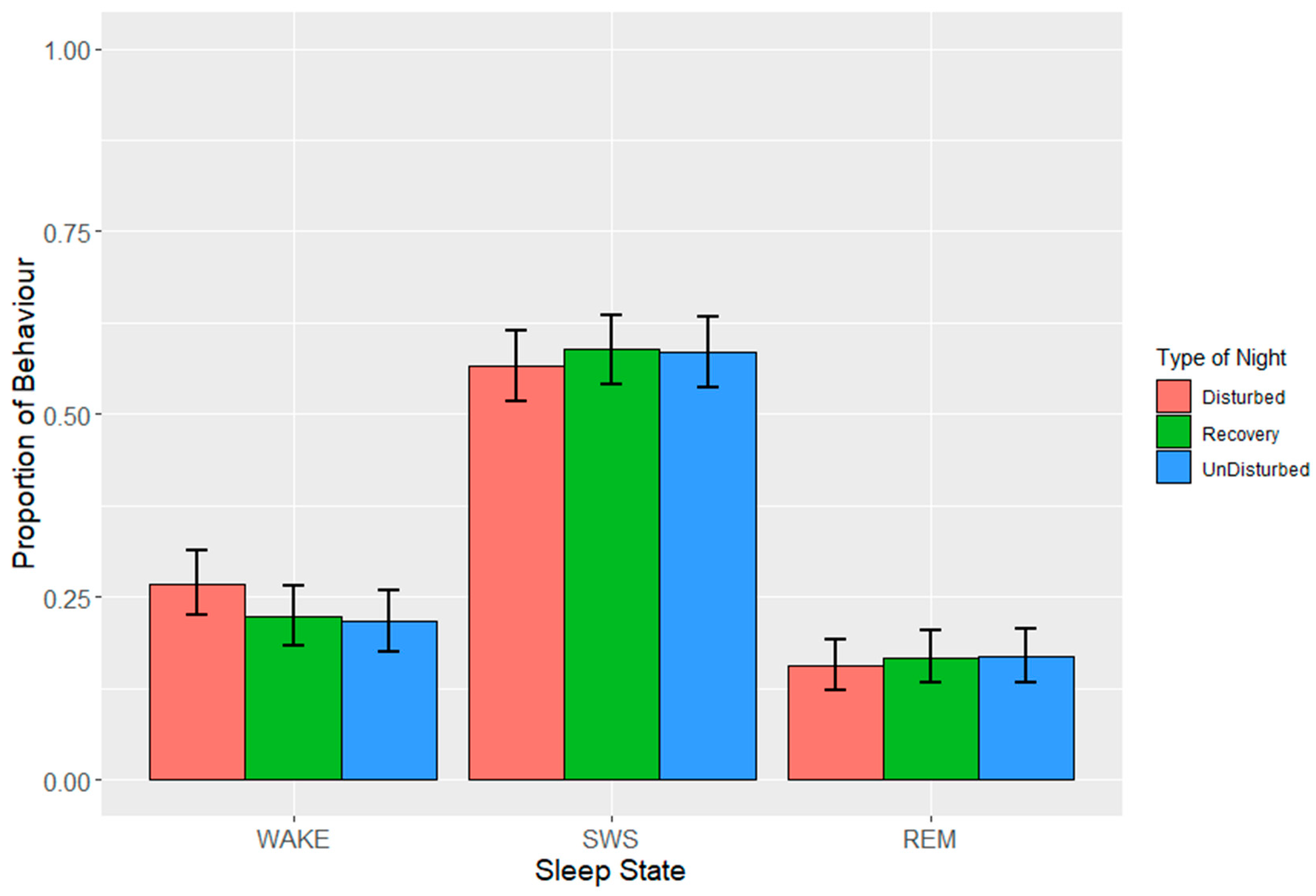
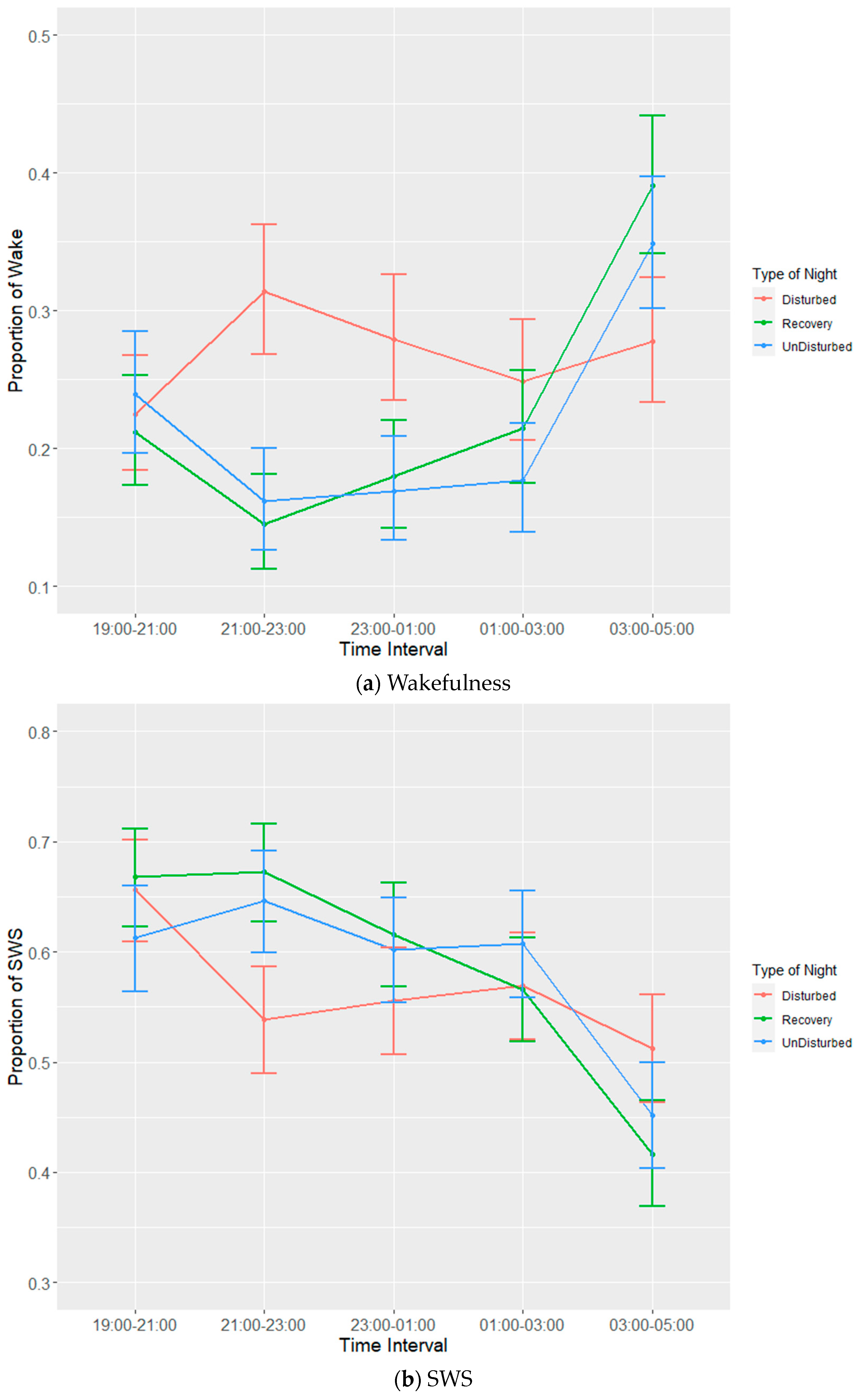
Data were grouped into 2 h (lights off) or 3.5 h (lights on) time intervals in order to account for potential temporal effects. Intervals for birds in which the total proportion of missing data + artefact
> 0.25 was omitted from further analysis after visual inspection of distributions. The counts for each sleep state were calculated for each bird in each interval from which proportions were calculated. Each 24 h period starting at lights off was classified by ‘type of night’, where experimental nights were categorized as either ‘disturbed’, ‘recovery’ or ‘undisturbed’ depending on whether disturbances were applied. In order to normalise residuals, proportions of sleep behaviours were angular transformed prior to fitting linear mixed models (LMMs). LMMs were applied separately for each sleep state and for lights off and lights on. Fixed effects were time interval, type of night and the interaction between them, and random effects were batch, bird, experimental day and time interval within experimental day. The reverse order of factors was also tested to ensure the results were similar.
p values were based on approximate F tests using Satterthwaite and Kenward–Roger methods for denominator degrees of freedom. Estimates of means ± standard errors (SEs) obtained from the models were back transformed onto the proportion scale to aid interpretation. Mean values are presented with lower and upper bounds calculated through the subtraction of the SE from the mean (lower bound) or addition of the SE to the mean (upper bound), which was subsequently back transformed, and the least and greatest values identified. Post hoc tests between estimated means were carried out using Tukey’s HSD test.
p values were compared against the standard of 0.05. Data processing and statistical analyses were carried out in the R system for statistical computing version 4.2.2 [
71], which was accessed via RStudio 2022.10.0 Build 353 (RStudio Team, 2022).















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}