
Total and Differential Somatic Cell Count in Italian Local Cattle Breeds: Phenotypic Variability and Effect on Milk Yield and Composition

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Editing
2.2. Statistical Analysis
3. Results
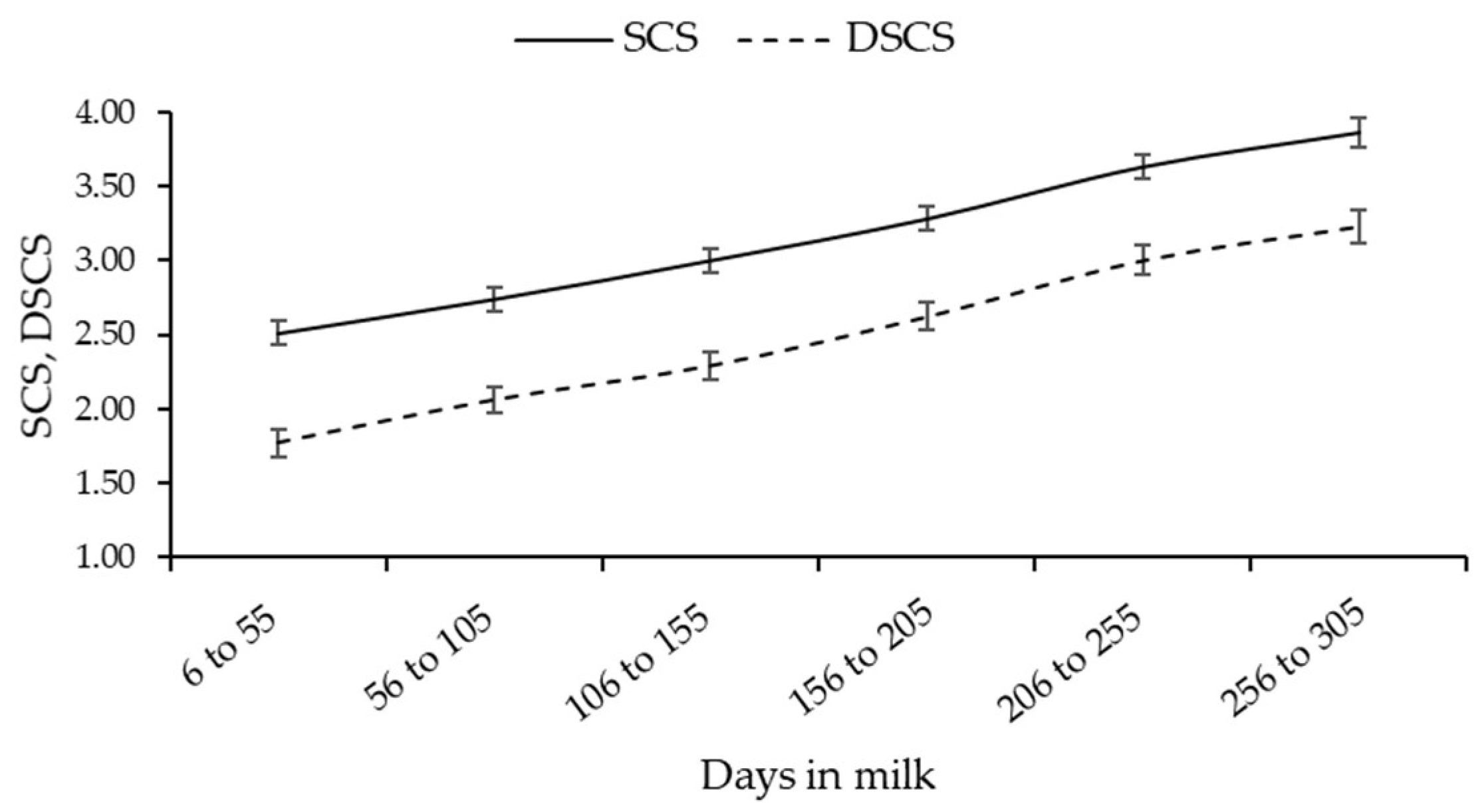
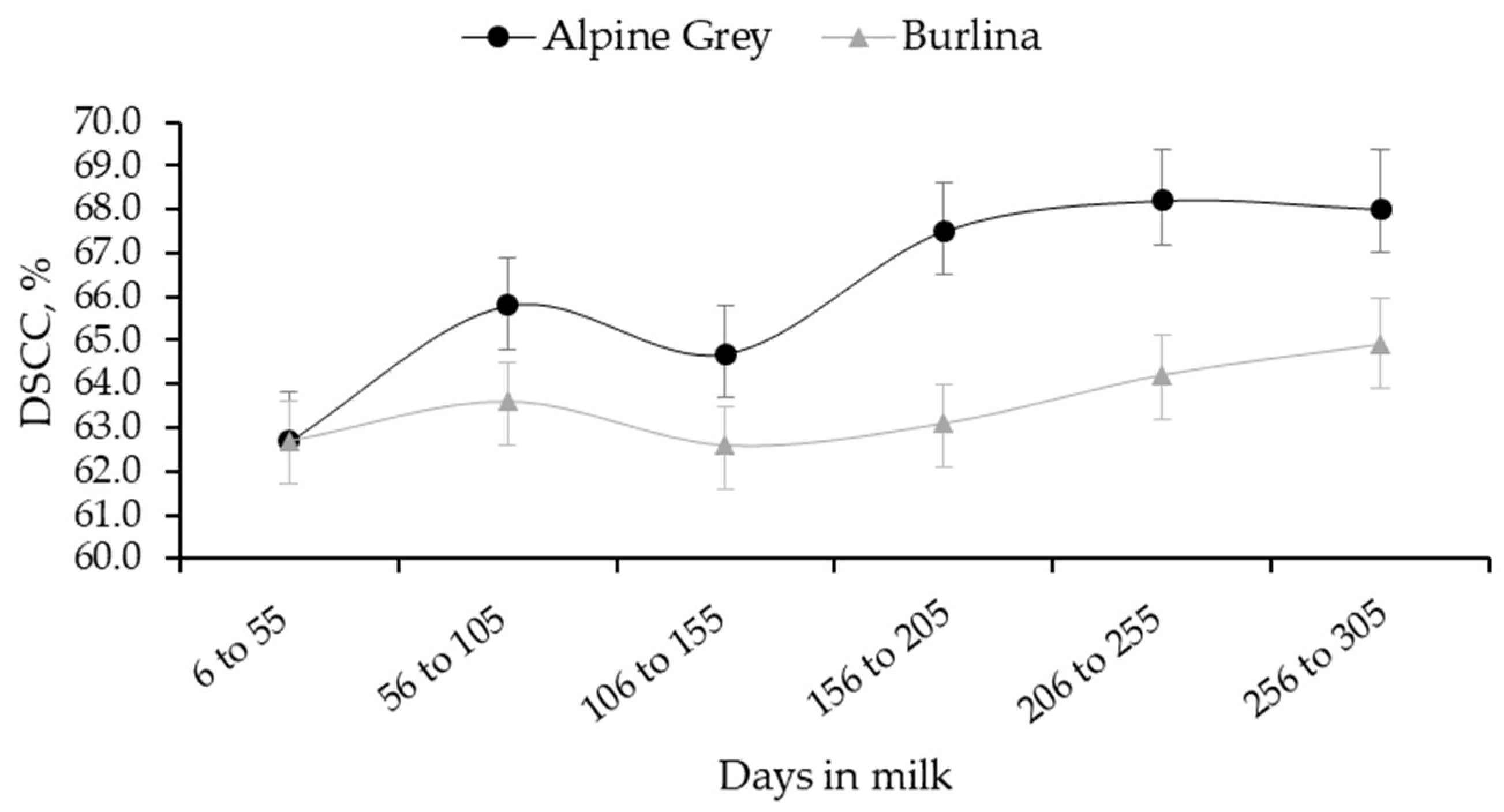
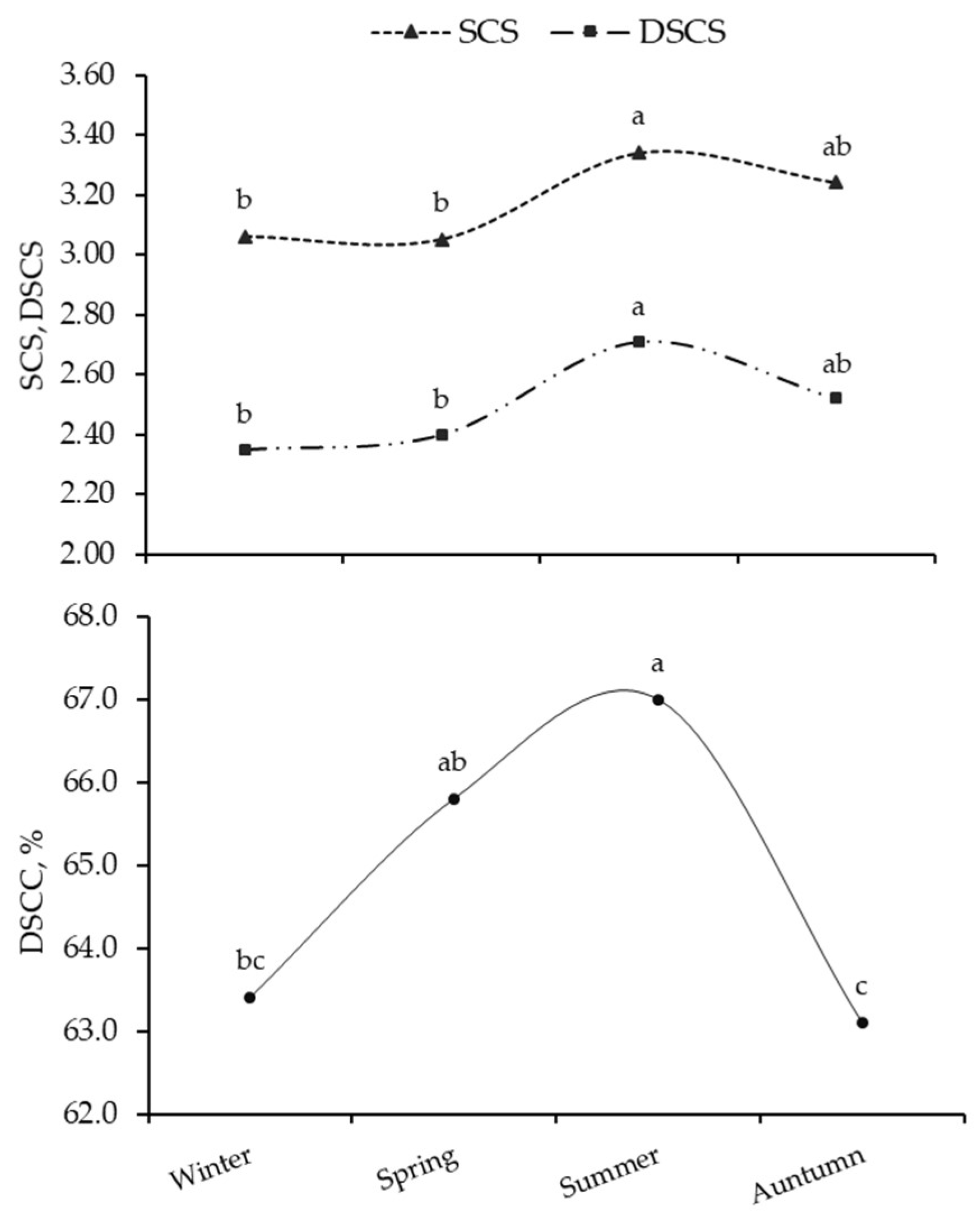
3.1. SCS and DSCC Variability
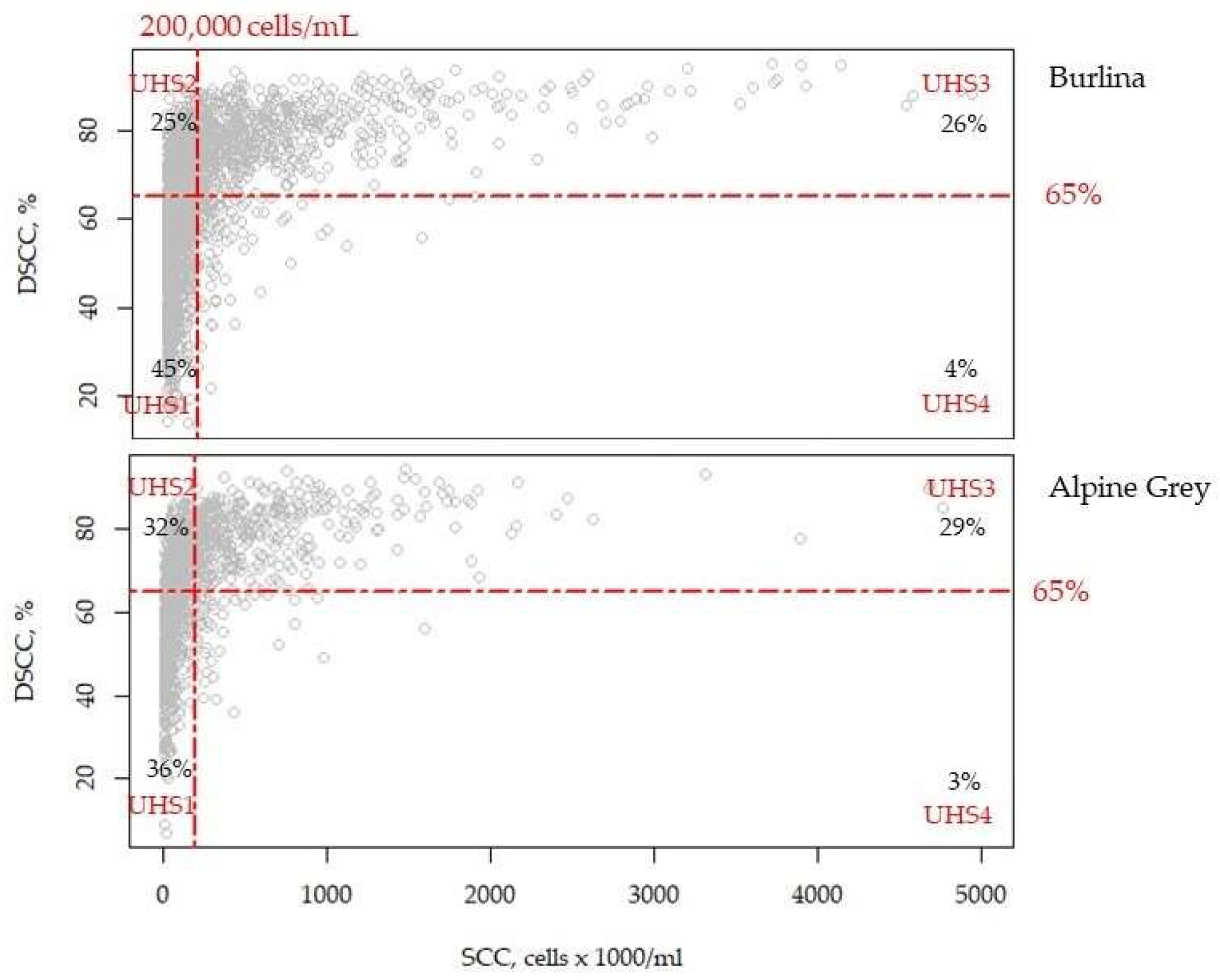
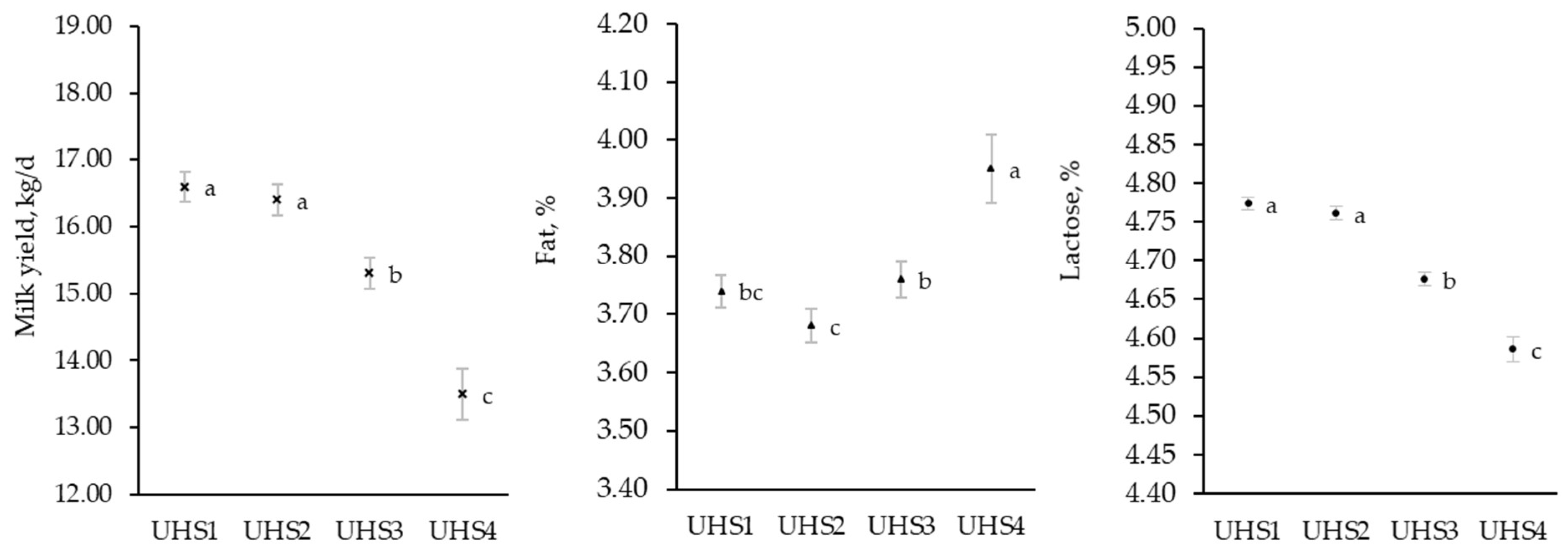
3.2. Effect of Udder Health
4. Discussion
4.1. SCS and DSCC Variability
4.2. Udder Health Status Group and Daily Production and Composition
4.3. DSCC and SCC in Practise
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Association of Breeders of Grey Alpine Cattle. Available online: www.grigioaplina.it (accessed on 23 May 2022).
- Domestic Animal Diversity Information System (DAD-IS). Available online: www.fao.org/dad-is/ (accessed on 23 May 2022).
- Dalvit, C.; De Marchi, M.; Dal Zotto, R.; Zanetti, E.; Meuwissen, T.; Cassandro, M. Genetic characterization of the Burlina cattle breed using microsatellites markers. J. Anim. Breed. Genet. 2008, 125, 137–144. [Google Scholar] [CrossRef]
- Curone, G.; Filipe, J.; Cremonesi, P.; Piccioli-Cappelli, F.; Trevisi, E.; Amadori, M. Relevance of the dairy cow biodiversity in the development of a profitable and environmentally sustainable livestock. CABI Rev. 2019, 14, 1–11. [Google Scholar] [CrossRef]
- Ruegg, P.L. A 100-Year Review: Mastitis detection, management, and prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damm, M.; Holm, C.; Blaabjerg, M.; Bro, M.N.; Schwarz, D. Differential somatic cell count—A novel method for routine mastitis screening in the frame of Dairy Herd Improvement testing programs. J. Dairy Sci. 2017, 100, 4926–4940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, D.; Kleinhans, S.; Reimann, G.; Stückler, P.; Reith, F.; Ilves, K.; Fouz, R. Investigation of dairy cow performance in different udder health groups defined based on a combination of somatic cell count and differential somatic cell count. Prev. Vet. Med. 2020, 183, 105123. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Santschi, D.E.; Durocher, J.; Lefebvre, D.M. Evaluation of the new differential somatic cell count parameter as a rapid and inexpensive supplementary tool for udder health management through regular milk recording. Prev. Vet. Med. 2020, 181, 105079. [Google Scholar] [CrossRef]
- Boutinaud, M.; Jammes, H. Potential uses of milk epithelial cells: A review. Reprod. Nutr. Dev. 2002, 42, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Leitner, G.; Eligulashvily, R.; Krifucks, O.; Perl, S.; Saran, A. Immune Cell Differentiation in Mammary Gland Tissues and Milk of Cows Chronically Infected with Staphylococcus aureus. J. Vet. Med. 2003, 50, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Leitner, G.; Shoshani, E.; Krifucks, O.; Chaffer, M.; Saran, A. Milk leucocyte population patterns in bovine udder infection of different aetiology. J. Vet. Med. 2000, 47, 581–589. [Google Scholar] [CrossRef] [Green Version]
- Ariel, O.; Gendron, D.; Dudemaine, P.L.; Gévry, N.; Ibeagha-Awemu, E.M.; Bissonnette, N. Transcriptome profiling of bovine macrophages infected by Mycobacterium avium spp. paratuberculosis depicts foam cell and innate immune tolerance phenotypes. Front. Immunol. 2020, 10, 2874. [Google Scholar] [CrossRef] [Green Version]
- Rienesl, L.; Marginter, M.; Stückler, P.; Köck, A.; Egger-Danner, C.; Sölkner, J. Use of differential somatic cell count, somatic cell score, and milk mid-infrared spectral analysis for monitoring mastitis in dairy cows during routine milk recording. Livest. Sci. 2022, 264, 105050. [Google Scholar] [CrossRef]
- Ali, A.K.A.; Shook, G.E. An optimum transformation for somatic cell concentration in milk. J. Dairy Sci. 1980, 63, 487–490. [Google Scholar] [CrossRef]
- Benedet, A.; Costa, A.; De Marchi, M.; Penasa, M. Heritability estimates of predicted blood β-hydroxybutyrate and nonesterified fatty acids and relationships with milk traits in early-lactation Holstein cows. J. Dairy Sci. 2020, 103, 6354–6363. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Associazione Italiana Allevatori (AIA). Available online: http://bollettino.aia.it/Contenuti.aspx?CD_GruppoStampe=RS&CD_Specie=C4 (accessed on 22 March 2023).
- Gottardo, P.; Penasa, M.; Righi, F.; Lopez-Villalobos, N.; Cassandro, M.; De Marchi, M. Fatty acid composition of milk from Holstein-Friesian, Brown Swiss, Simmental and Alpine Grey cows predicted by mid-infrared spectroscopy. Ital. J. Anim. Sci. 2017, 16, 380–389. [Google Scholar] [CrossRef] [Green Version]
- Visentin, G.; Penasa, M.; Niero, G.; Cassandro, M.; De Marchi, M. Phenotypic characterisation of major mineral composition predicted by mid-infrared spectroscopy in cow milk. Ital. J. Anim. Sci. 2018, 17, 549–556. [Google Scholar] [CrossRef] [Green Version]
- Penasa, M.; Cecchinato, A.; Battagin, M.; De Marchi, M.; Pretto, D.; Cassandro, M. Bayesian inference of genetic parameters for test-day milk yield, milk quality traits, and somatic cell score in Burlina cows. J. Appl. Genet. 2020, 51, 489–495. [Google Scholar] [CrossRef]
- Niero, G.; Visentin, G.; Ton, S.; De Marchi, M.; Penasa, M.; Cassandro, M. Phenotypic characterisation of milk technological traits, protein fractions, and major mineral and fatty acid composition of Burlina cattle breed. Ital. J. Anim. Sci. 2016, 15, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Cassandro, M.; Comin, A.; Ojala, M.; Dal Zotto, R.; DeMarchi, M.; Gallo, L.; Carnier, P.; Bittante, G. Genetic parameters of milk coagulation properties and their relation-ships with milk yield and quality traits in Italian Holstein cows. J. Dairy Sci. 2008, 91, 371–376. [Google Scholar] [CrossRef] [Green Version]
- Bobbo, T.; Penasa, M.; Cassandro, M. Combining total and differential somatic cell count to better assess the association of udder health status with milk yield, composition and coagulation properties in cattle. Ital. J. Anim. Sci. 2020, 19, 697–703. [Google Scholar] [CrossRef]
- Kirkeby, C.; Toft, N.; Schwarz, D.; Farre, M.; Nielsen, S.S.; Zervens, L.; Hechinger, S.; Halasa, T. Differential somatic cell count as an additional indicator for intramammary infections in dairy cows. J. Dairy Sci. 2020, 103, 1759–1775. [Google Scholar] [CrossRef] [Green Version]
- Halasa, T.; Kirkeby, C. Differential somatic cell count: Value for udder health management. Front. Vet. Sci. 2022, 7, 1153. [Google Scholar] [CrossRef] [PubMed]
- Stocco, G.; Cipolat-Gotet, C.; Stefanon, B.; Zecconi, A.; Francescutti, M.; Mountricha, M.; Summer, A. Herd and animal factors affect the variability of total and differential somatic cell count in bovine milk. J. Anim. Sci. 2023, 101, skac406. [Google Scholar] [CrossRef]
- Sharma, N.; Singh, N.K.; Bhadwal, M.S. Relationship of somatic cell count and mastitis: An overview. Asian Australas. J. Anim. Sci. 2011, 24, 429–438. [Google Scholar] [CrossRef]
- Zhao, X.; Lacasse, P. Mammary tissue damage during bovine mastitis: Causes and control. J. Anim. Sci. 2008, 86, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Bovenhuis, H.; Penasa, M. Changes in milk lactose content as indicators for longevity and udder health in Holstein cows. J. Dairy Sci. 2020, 103, 11574–11584. [Google Scholar] [CrossRef] [PubMed]
- Paape, M.; Mehrzad, J.; Zhao, X.; Detilleux, J.; Burvenich, C. Defense of the bovine mammary gland by polymorphonuclear neutrophil leukocytes. J. Mammary Gland Biol. Neoplasia 2002, 7, 109–121. [Google Scholar] [CrossRef]
- Haenlein, G.; Schultz, H.; Zikakis, J.P. Composition of protein in milk with varying leukocyte content. J. Dairy Sci. 1973, 56, 1017–1024. [Google Scholar] [CrossRef]
- Mariani, E.; Cipolat-Gotet, C.; Stefanon, B.; Zecconi, A.; Stocco, G.; Sandri, M.; Summer, A. Effect of total and differential somatic cell count on yield, composition and predicted coagulation properties from individual dairy cows. Int. J. Dairy Technol. 2022, 75, 298–307. [Google Scholar] [CrossRef]
- Herve, L.; Lollivier, V.; Quesnel, H.; Boutinaud, M. Oxytocin induces mammary epithelium disruption and could stimulate epithelial cell exfoliation. J. Mammary Gland Biol. Neoplasia 2018, 23, 139–147. [Google Scholar] [CrossRef]
- Costa, A.; Lopez-Villalobos, N.; Sneddon, N.W.; Shalloo, L.; Franzoi, M.; De Marchi, M.; Penasa, M. Invited review: Milk lactose—Current status and future challenges in dairy cattle. J. Dairy Sci. 2019, 102, 5883–5898. [Google Scholar] [CrossRef]
- Moyes, K.M.; Larsen, T.; Sørensen, P.; Ingvartsen, K.L. Changes in various metabolic parameters in blood and milk during experimental Escherichia coli mastitis for primiparous Holstein dairy cows during early lactation. J. Anim. Sci. Biotechnol. 2014, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pegolo, S.; Giannuzzi, D.; Bisutti, V.; Tessari, R.; Gelain, M.E.; Gallo, L.; Cecchinato, A. Associations between differential somatic cell count and milk yield, quality, and technological characteristics in Holstein cows. J. Dairy Sci. 2021, 104, 4822–4836. [Google Scholar] [CrossRef] [PubMed]
- Hagnestam-Nielsen, C.; Emanuelson, U.; Berglund, B.; Strandberg, E. Relationship between somatic cell count and milk yield in different stages of lactation. J. Dairy Sci. 2009, 92, 3124–3133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banga, C.B.; Neser, F.W.; Garrick, D.J. The economic value of somatic cell count in South African Holstein and Jersey cattle. S. Afr. J. Anim. Sci. 2014, 44, 173–177. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait 1 | Burlina | Alpine Grey | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Min. | Max. | Mean | SD | Min. | Max. | |
| Milk yield, kg/d | 16.99 | 6.70 | 1.80 | 37.3 | 16.99 | 5.85 | 2.20 | 33.6 |
| Fat, % | 3.80 | 0.73 | 1.45 | 6.22 | 3.57 | 0.63 | 1.57 | 5.59 |
| Protein, % | 3.35 | 0.41 | 2.21 | 4.63 | 3.37 | 0.39 | 2.43 | 4.55 |
| Casein, % | 2.62 | 0.33 | 1.64 | 3.66 | 2.65 | 0.32 | 1.81 | 3.63 |
| Casein- index | 0.78 | 0.02 | 0.69 | 0.84 | 0.78 | 0.02 | 0.69 | 0.83 |
| Lactose, % | 4.72 | 0.23 | 3.92 | 5.31 | 4.80 | 0.21 | 4.06 | 5.49 |
| SCS | 3.06 | 1.87 | −0.32 | 8.63 | 3.13 | 1.82 | −0.32 | 8.57 |
| DSCC, % | 63.5 | 16.4 | 13.7 | 95.0 | 66.8 | 14.3 | 7.2 | 94.1 |
| DSCS | 2.34 | 2.16 | −3.15 | 8.45 | 2.51 | 2.10 | −3.05 | 8.40 |
| BHB | −1.30 | 0.36 | −2.00 | −0.24 | −1.51 | 0.51 | −3.00 | −0.54 |
| Urea, mg/dL | 23.55 | 6.97 | 6.6 | 44.8 | 23.59 | 6.46 | 5.90 | 43.6 |
| Trait 2 | SCS | DSCC | DSCS |
|---|---|---|---|
| DSCC, % | 0.66 | ||
| DSCS | 0.99 | 0.77 | |
| Milk yield, kg/d | −0.23 | −0.08 | −0.22 |
| Fat, % | 0.08 | −0.08 | 0.05 |
| Protein, % | 0.20 | −0.02 ns | 0.17 |
| Casein, % | 0.16 | −0.03 ns | 0.13 |
| Casein index | −0.19 | −0.04 | −0.17 |
| Lactose, % | −0.43 | −0.13 | −0.39 |
| BHB | 0.08 | 0.03 | 0.08 |
| Urea, mg/dL | −0.01 ns | −0.01 ns | −0.01 ns |
| Trait | Breed | Parity | ||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ≥5 | ||
| SCS | Alpine Grey | 2.53 c | 2.48 c | 3.10 b | 2.87 bc | 3.93 a |
| Burlina | 2.50 c | 2.85 b | 2.94 b | 3.25 ab | 3.44 a | |
| DSCC | Alpine Grey | 64.8 b | 63.4 b | 66.7 ab | 63.4 b | 71.0 a |
| Burlina | 62.5 ab | 62.3 ab | 61.0 b | 64.2 ab | 65.8 a | |
| DSCS | Alpine Grey | 2.03 bc | 1.95 c | 2.65 b | 2.38 bc | 3.58 a |
| Burlina | 1.95 c | 2.28 bc | 2.33 bc | 2.73 ab | 2.96 a | |
| Trait | Breed | UHS1 | UHS2 | UHS3 | UHS4 |
|---|---|---|---|---|---|
| Protein, % | Alpine Grey | 3.41 b | 3.41 b | 3.48 a | 3.53 a |
| Burlina | 3.39 c | 3.35 d | 3.44 b | 3.60 a | |
| Casein, % | Alpine Grey | 2.69 b | 2.70 ab | 2.73 a | 2.74 a |
| Burlina | 2.66 b | 2.63 c | 2.69 b | 2.80 a | |
| Casein-index | Alpine Grey | 0.787 a | 0.789 a | 0.782 b | 0.777 c |
| Burlina | 0.786 a | 0.786 a | 0.781 b | 0.777 b | |
| Urea, mg/dL | Alpine Grey | 24.00 a | 23.70 a | 23.40 a | 23.50 a |
| Burlina | 23.30 a | 23.20 ab | 22.60 b | 23.50 ab | |
| BHB2 | Alpine Grey | −1.58 c | −1.54 bc | −1.47 b | −1.22 a |
| Burlina | −1.31 b | −1.32 b | −1.22 a | −1.16 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magro, S.; Costa, A.; De Marchi, M. Total and Differential Somatic Cell Count in Italian Local Cattle Breeds: Phenotypic Variability and Effect on Milk Yield and Composition. Animals 2023, 13, 1249. https://doi.org/10.3390/ani13071249
Magro S, Costa A, De Marchi M. Total and Differential Somatic Cell Count in Italian Local Cattle Breeds: Phenotypic Variability and Effect on Milk Yield and Composition. Animals. 2023; 13(7):1249. https://doi.org/10.3390/ani13071249
Chicago/Turabian StyleMagro, Silvia, Angela Costa, and Massimo De Marchi. 2023. "Total and Differential Somatic Cell Count in Italian Local Cattle Breeds: Phenotypic Variability and Effect on Milk Yield and Composition" Animals 13, no. 7: 1249. https://doi.org/10.3390/ani13071249