New Insights into the Variation and Admixture of the Cave-Dwelling Spider Trogloneta yunnanensis in South China Karst


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
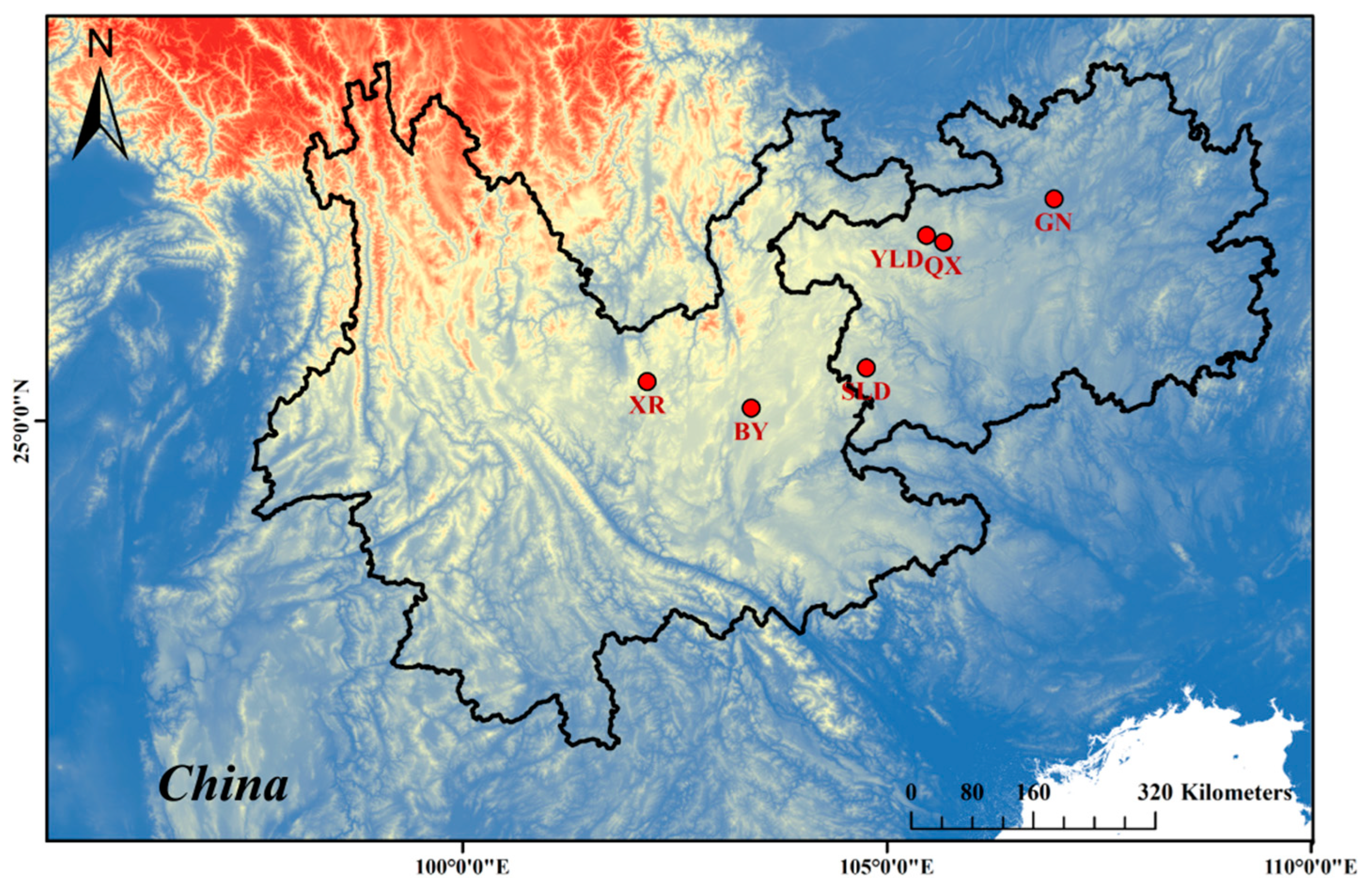
2.1. Sampling
2.2. DNA Extraction, Sample Preparation, and Gene Sequencing
2.3. Population Genetic Analysis
2.4. Phylogenetic Analysis
2.5. Divergence Time Estimation
3. Results
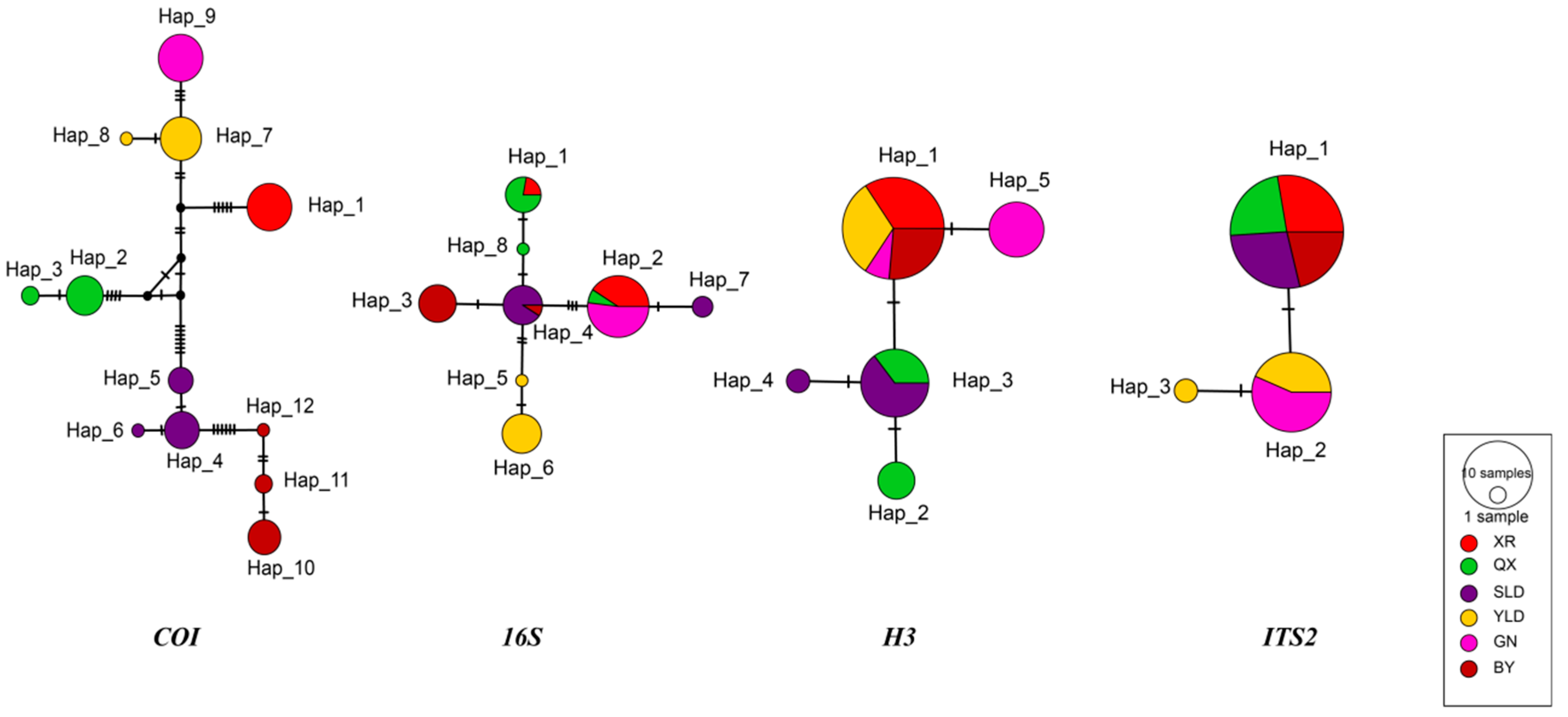
3.1. Genetic Diversity and Structure of Trogloneta yunnanensis
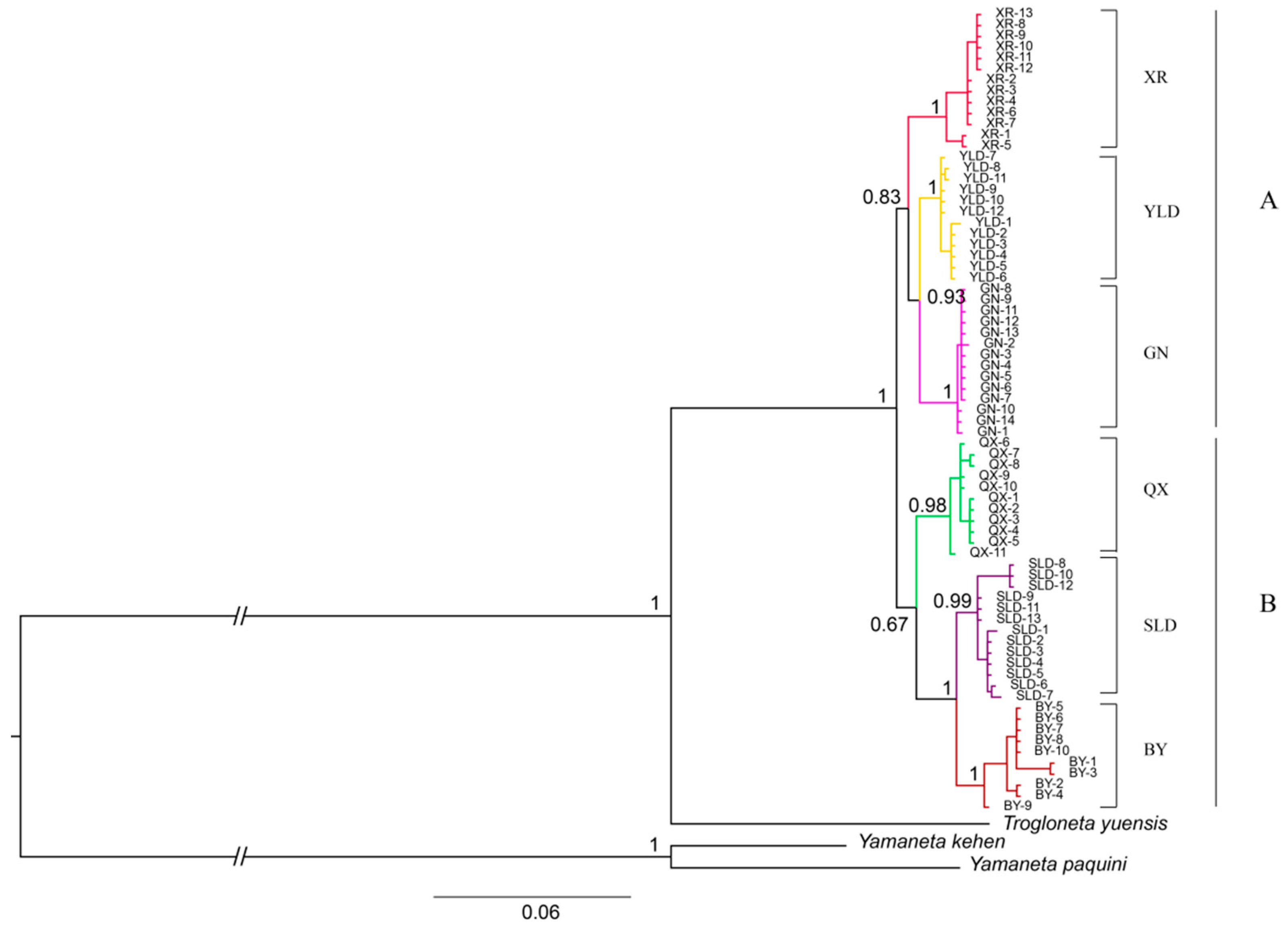
3.2. Phylogenetic Relationships
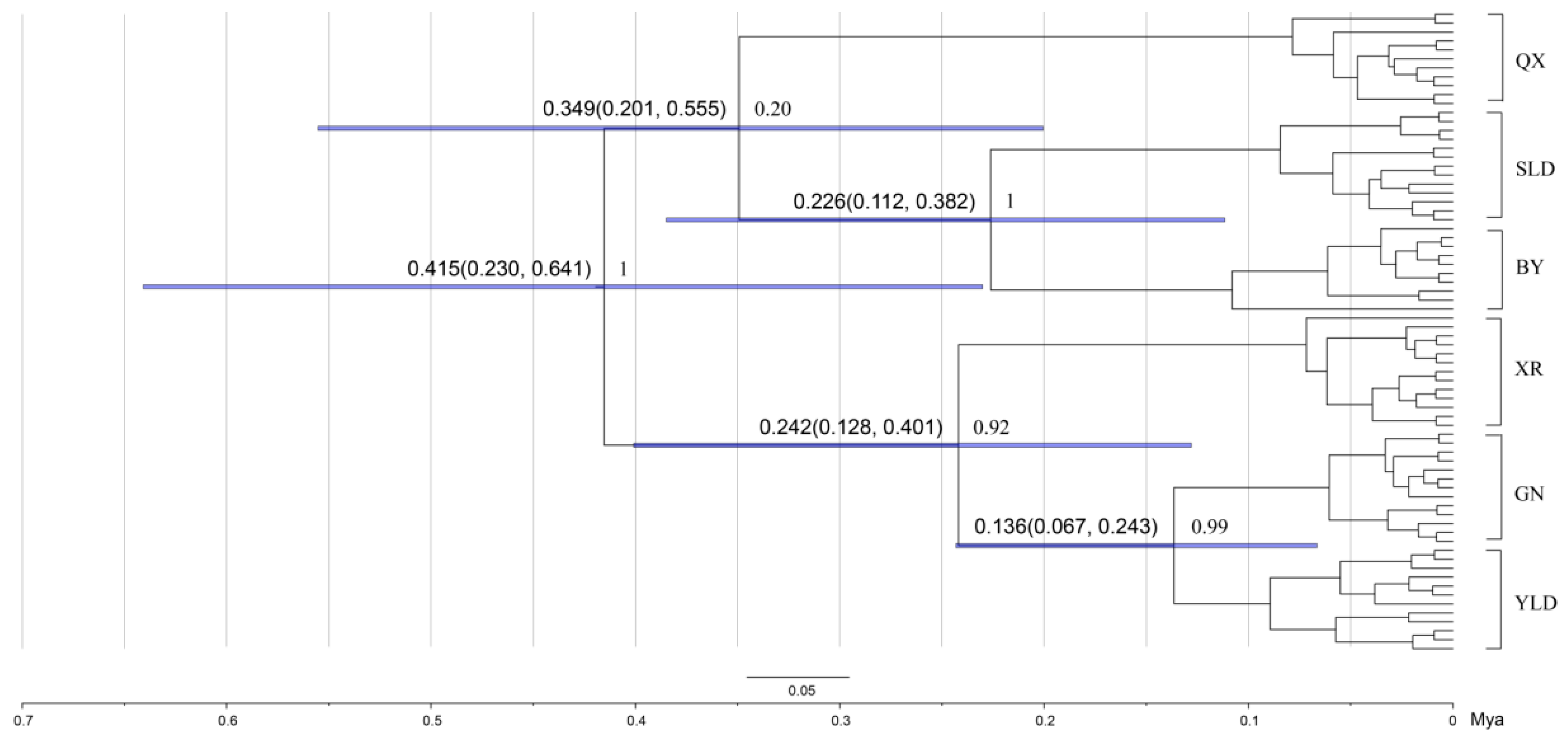
3.3. Divergence Time Estimation
4. Discussion
4.1. Variation and Admixture
4.2. Impact of Geographical Isolation
4.3. Evolutionary History
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cerca, J. On the Origin of Cryptic Species: Insights from the Stygocapitella Species Complex. Ph.D. Thesis, University of Oslo, Oslo, Norway, 2020. [Google Scholar]
- Mammola, S.; Lunghi, E.; Bilandžija, H.; Cardoso, P.; Grimm, V.; Schmidt, S.I.; Hesselberg, T.; Martínez, A. Collecting eco-evolutionary data in the dark: Impediments to subterranean research and how to overcome them. Ecol. Evol. 2021, 11, 5911–5926. [Google Scholar] [CrossRef]
- Howarth, F.G. Ecology of Cave Arthropods. Annu. Rev. Entomol. 1983, 28, 365–389. [Google Scholar] [CrossRef]
- Culver, D.C.; Holsinger, J.R. How Many Species of Troglobites Are There? Natl. Speleol. Soc. 1992, 54, 79–80. [Google Scholar]
- Juan, C.; Guzik, M.T.; Jaume, D.; Cooper, S.J. Evolution in caves: Darwin’s ‘wrecks of ancient life’ in the molecular era. Mol. Ecol. 2010, 19, 3865–3880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Moreno, J.L.; Balázs, G.; Wilkins, B.; Herczeg, G.; Bracken-Grissom, H.D. The role of isolation on contrasting phylogeographic patterns in two cave crustaceans. BMC Evol. Biol. 2017, 17, 247. [Google Scholar] [CrossRef] [Green Version]
- Faille, A.; Tänzler, R.; Toussaint, E.F. On the Way to Speciation: Shedding Light on the Karstic Phylogeography of the Microendemic Cave Beetle Aphaenops cerberus in the Pyrenees. J. Hered. 2015, 106, 692–699. [Google Scholar] [CrossRef] [Green Version]
- Ballarin, F.; Li, S. Diversification in tropics and subtropics following the mid-Miocene climate change: A case study of the spider genus Nesticella. Glob. Chang. Biol. 2018, 24, e577–e591. [Google Scholar] [CrossRef]
- Gibert, J.; Deharveng, L. Subterranean Ecosystems: A Truncated Functional Biodiversity. Bioscience 2002, 52, 473–481. [Google Scholar] [CrossRef]
- Culver, D.C.; Master, L.L.; Christman, M.C.; Hobbs, H.H. Obligate Cave Fauna of the 48 Contiguous United States. Conserv. Biol. 2000, 14, 386–401. [Google Scholar] [CrossRef]
- Snowman, C.; Zigler, K.; Hedin, M. Caves as islands: Mitochondrial phylogeography of the cave-obligate spider species Nesticus barri (Araneae: Nesticidae). J. Arachnol. 2010, 38, 49–56. [Google Scholar] [CrossRef]
- Hedin, M.C. Molecular phylogenetics at the population/species interface in cave spiders of the southern Appalachians (Araneae:Nesticidae:Nesticus). Mol. Biol. Evol. 1997, 14, 309–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammola, S.; Isaia, M.; Arnedo, M.A. Alpine endemic spiders shed light on the origin and evolution of subterranean species. PeerJ 2015, 3, e1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammola, S.; Hormiga, G.; Arnedo, M.A.; Isaia, M. Unexpected diversity in the relictual European spiders of the genus Pimoa (Araneae: Pimoidae). Invertebr. Syst. 2016, 30, 566–587. [Google Scholar] [CrossRef] [Green Version]
- Paquin, P.; Hedin, M. The power and perils of ‘molecular taxonomy’: A case study of eyeless and endangered Cicurina (Araneae: Dictynidae) from Texas caves. Mol. Ecol. 2004, 13, 3239–3255. [Google Scholar] [CrossRef]
- Caccone, A. Gene flow in cave arthropods: A qualitative and quantitative approach. Evolution 1985, 39, 1223–1235. [Google Scholar] [CrossRef]
- Weckstein, J.D.; Johnson, K.P.; Murdoch, J.D.; Krejca, J.K.; Takiya, D.M.; Veni, G.; Reddell, J.R.; Taylor, S.J. Comparative phylogeography of two codistributed subgenera of cave crickets (Orthoptera: Rhaphidophoridae: Ceuthophilus spp.). J. Biogeogr. 2016, 43, 1450–1463. [Google Scholar] [CrossRef]
- Katz, A.D.; Taylor, S.J.; Davis, M.A. At the confluence of vicariance and dispersal: Phylogeography of cavernicolous springtails (Collembola: Arrhopalitidae, Tomoceridae) codistributed across a geologically complex karst landscape in Illinois and Missouri. Ecol. Evol. 2018, 8, 10306–10325. [Google Scholar] [CrossRef] [Green Version]
- Mammola, S.; Cardoso, P.; Ribera, C.; Pavlek, M.; Isaia, M.J.J.o.Z.S.; Research, E. A synthesis on cave-dwelling spiders in Europe. J. Zool. Syst. Evol. Res. 2018, 56, 301–316. [Google Scholar] [CrossRef]
- Luo, Z.; Tang, S.; Jiang, Z.; Chen, J.; Fang, H.; Li, C. Conservation of Terrestrial Vertebrates in a Global Hotspot of Karst Area in Southwestern China. Sci. Rep. 2016, 6, 25717. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R. Zoogeography of China; Press, S., Ed.; China Science Publishing & Media: Beijing, China, 2011. [Google Scholar]
- Yang, J.; Yang, J.; Chen, X. A re-examination of the molecular phylogeny and biogeography of the genus Schizothorax (Teleostei: Cyprinidae) through enhanced sampling, with emphasis on the species in the Yunnan–Guizhou Plateau, China. J. Zool. Syst. Evol. Res. 2012, 50, 184–191. [Google Scholar] [CrossRef]
- Yan, Y.L. The Origin and Evolution of Cave-Dwelling Group of Triplophysa Fishes (Teleostei, Cypriniformes, Nemacheilidae). Ph.D. Thesis, Southwest University, Chongqing, China, 2017. [Google Scholar]
- Li, X. Nemacheilidae Cavefishes of China—Taxonomy, Adaption and Zoogeography. Ph.D. Thesis, Shanghai Ocean University, Shanghai, China, 2018. [Google Scholar]
- Zhang, Y.; Li, S. A spider species complex revealed high cryptic diversity in South China caves. Mol. Phylogenet. Evol. 2014, 79, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.S.; Jiang, W.S.; Whitten, T.; Yang, J.X.; Chen, X.Y. Drought and China’s cave species. Science 2013, 340, 272. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Zhu, M. On some species of cave arachnids of China. In Sixtieth Anniversary of the Founding of China Zoological Society: Memorial Volume Dedicated to the Hundredth Anniversary of the Birthday of the Late Prof. Sisan Chen (Z. Chen); Chen, Y.Z., Ed.; China Science and Technology Press: Beijing, China, 1994; pp. 35–36. [Google Scholar]
- Lin, Y.; Li, S. Mysmenid Spiders of China (Araneae: Mysmenidae). Ann. Zool. 2008, 58, 487–520. [Google Scholar] [CrossRef]
- Li, Y.; Lin, Y. Taxonomic review of the Asian Trogloneta species (Araneae, Mysmenidae). Zookeys 2019, 817, 41–60. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Li, S. Two new species of the genera Mysmena and Trogloneta (Mysmenidae, Araneae) from Southwestern China. Zookeys 2013, 303, 33–51. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Miller, J.A.; Lin, Y.; Shu, Y. Further study of two Chinese cave spiders (Araneae, Mysmenidae), with description of a new genus. Zookeys 2019, 870, 77–100. [Google Scholar] [CrossRef]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version s2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [Green Version]
- Leigh, J.W.; Bryant, D. popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. Online 2007, 1, 47–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A.J.V.E. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Bidegaray-Batista, L.; Arnedo, M.A. Gone with the plate: The opening of the Western Mediterranean basin drove the diversification of ground-dweller spiders. BMC Evol. Biol. 2011, 11, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derkarabetian, S.; Steinmann, D.B.; Hedin, M.J.P.O. Repeated and Time-Correlated Morphological Convergence in Cave-Dwelling Harvestmen (Opiliones, Laniatores) from Montane Western North America. PLoS ONE 2010, 5, e10388. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, A.; Anastasiou, I.; Vogler, A.P. Revisiting the insect mitochondrial molecular clock: The mid-Aegean trench calibration. Mol. Biol. Evol. 2010, 27, 1659–1672. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.S.; Bolton, M.; Beard, A.; Birt, T.; Deane-Coe, P.; Raine, A.F.; GonzálezSolís, J.; Lougheed, S.C.; Friesen, V.L. Cryptic species and independent origins of allochronic populations within a seabird species complex (Hydrobates spp.). Mol. Phylogenet. Evol. 2019, 139, 106552. [Google Scholar] [CrossRef]
- Slavenko, A.; Tamar, K.; Tallowin, O.J.S.; Allison, A.; Kraus, F.; Carranza, S.; Meiri, S. Cryptic diversity and non-adaptive radiation of montane New Guinea skinks (Papuascincus; Scincidae). Mol. Phylogenet. Evol. 2020, 146, 106749. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.O.; Hutter, C.R.; Wood, P.L.; Grismer, L.L.; Das, I.; Brown, R.M. Gene flow creates a mirage of cryptic species in a Southeast Asian spotted stream frog complex. Mol. Ecol. 2020, 29, 3970–3987. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, L.A.; Mateos, M.; Mattos, G.; Liu, S.; Haye, P.A.; Paiva, P.C. Multiple transisthmian divergences, extensive cryptic diversity, occasional long-distance dispersal, and biogeographic patterns in a marine coastal isopod with an amphi-American distribution. Ecol. Evol. 2016, 6, 7794–7808. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, S. Ancient lineage, young troglobites: Recent colonization of caves by Nesticella spiders. BMC Evol. Biol. 2013, 13, 183. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Eronen, J.T.; Kaakinen, A.; Utescher, T.; Ahrens, B.; Fortelius, M. Strong winter monsoon wind causes surface cooling over India and China in the Late Miocene. Clim. Past Discuss. 2015, 11, 63–93. [Google Scholar]
- Jacques, F.; Shi, G.; Wang, W.M. Neogene zonal vegetation of China and the evolution of the winter monsoon. Bull. Geosci. 2013, 88, 175–193. [Google Scholar] [CrossRef] [Green Version]
- Colgan, D.J.; McLauchlan, A.; Wilson, G.D.F.; Livingston, S.P.; Edgecombe, G.D.; Macaranas, J.; Cassis, G.; Gray, M.R. Histone H3 and U2 snRNA DNA sequences and arthropod molecular evolution. Aust. J. Zool. 1998, 46, 419. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Hormiga, G.; Arnedo, M.; Gillespie, R.G. Speciation on a Conveyor Belt: Sequential Colonization of the Hawaiian Islands by Orsonwelles Spiders (Araneae, Linyphiidae). Syst. Biol. 2003, 52, 70–88. [Google Scholar] [CrossRef] [PubMed]
- Rix, M.G.; Harvey, M.S.; Roberts, J.D. Molecular phylogenetics of the spider family Micropholcommatidae (Arachnida: Araneae) using nuclear rRNA genes (18S and 28S). Mol. Phylogenet. Evol. 2008, 46, 1031–1048. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: New York, NY, USA, 1990. [Google Scholar]
- Yan, F.H. Molecular Systematics of the Mysmenidae (Arachnida, Araneae) from China. Master’s Thesis, Sichuan University, Chengdu, China, 2018. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sites (Abbrs.) | Sample Size | Geographic Coordinates | Collection Localities |
|---|---|---|---|---|
| Trogloneta yunnanensis | Guanniu Cave (GN) | 6♂8♀ | 27.6137° N, 106.9691° E | Guizhou: Zunyi City, Shenxi Twon, Longjiang Vill. |
| Yelaoda Cave (YLD) | 6♂6♀ | 27.1843° N, 105.4657° E | Guizhou: Dafang Co., Wenge Town, Sanhe Vill. | |
| Qingxu Cave (QX) | 4♂7♀ | 27.1030° N, 105.6699° E | Guizhou: Dafang Co., Yangchang Town, Longdong Vill. | |
| Shilida Cave (SLD) | 6♂7♀ | 25.6237° N, 104.7566° E | Guizhou: Panxian Co., Zhudong Town, Shiliping Vill. | |
| Xianren Cave (XR) | 7♂7♀ | 25.4648° N, 102.1729° E | Yunnan: Wuding Co., Maojie Town | |
| Baiyan Cave (BY) | 5♂5♀ | 25.1510° N, 103.4010° E | Yunnan: Yiliang Co., Jiuxiang Town, Dazhezong Vill. | |
| Trogloneta yuensis | Yuelu Mt. Parkland | 1♂1♀ | 28.1869° N, 112.9421° E | Hunan: Changsha City, Yuelu Dist. |
| Populations | cox1 | 16S | H3 | ITS-2 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | H | π | Hd | N | H | π | Hd | N | H | π | Hd | N | H | π | Hd | |
| XR | 13 | 1 | 0.00000 | 0.000 | 13 | 2 | 0.00336 | 0.282 | 13 | 1 | 0.00000 | 0.000 | 13 | 1 | 0.00000 | 0.000 |
| QX | 11 | 2 | 0.00052 | 0.327 | 11 | 2 | 0.00043 | 0.182 | 11 | 2 | 0.00175 | 0.545 | 11 | 1 | 0.00000 | 0.000 |
| SLD | 13 | 3 | 0.00098 | 0.564 | 13 | 2 | 0.00366 | 0.385 | 13 | 2 | 0.00090 | 0.282 | 13 | 1 | 0.00000 | 0.000 |
| YLD | 12 | 2 | 0.00026 | 0.167 | 12 | 2 | 0.00040 | 0.167 | 12 | 1 | 0.00000 | 0.000 | 12 | 2 | 0.00082 | 0.303 |
| GN | 13 | 1 | 0.00000 | 0.000 | 14 | 1 | 0 | 0 | 14 | 2 | 0.00116 | 0.363 | 13 | 1 | 0.00000 | 0.000 |
| BY | 10 | 3 | 0.00138 | 0.511 | 10 | 3 | 0.00450 | 0.511 | 10 | 1 | 0.00000 | 0.000 | 10 | 1 | 0.00000 | 0.000 |
| XR | QX | SLD | YLD | GN | |
|---|---|---|---|---|---|
| QX | 0.99 *** | ||||
| SLD | 0.98 *** | 0.96 *** | |||
| YLD | 0.99 *** | 0.97 *** | 0.97 *** | ||
| GN | 1.00 *** | 0.99 *** | 0.98 *** | 0.97 *** | |
| BY | 0.97 *** | 0.96 *** | 0.92 *** | 0.97 *** | 0.98 *** |
| XR | QX | SLD | YLD | GN | |
|---|---|---|---|---|---|
| QX | 0.01 | ||||
| SLD | 0.01 | 0.02 | |||
| YLD | 0.01 | 0.02 | 0.02 | ||
| GN | 0.00 | 0.01 | 0.01 | 0.02 | |
| BY | 0.02 | 0.02 | 0.04 | 0.02 | 0.01 |
| XR | QX | SLD | YLD | GN | |
|---|---|---|---|---|---|
| QX | 0.0184 | ||||
| SLD | 0.0253 | 0.0210 | |||
| YLD | 0.0109 | 0.0142 | 0.0212 | ||
| GN | 0.0151 | 0.0178 | 0.0253 | 0.0049 | |
| BY | 0.0222 | 0.0209 | 0.0149 | 0.0210 | 0.0252 |
| Source of Variation | d. f. | Sum of Sequence | Variance Components | Percentage of Variation |
|---|---|---|---|---|
| Among populations | 5 | 346.132 | 5.767 | 97.4 |
| Within populations | 66 | 10.145 | 0.153 | 2.6 |
| Total | 71 | 356.278 | 5.920 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Zhou, C.; Lin, Y. New Insights into the Variation and Admixture of the Cave-Dwelling Spider Trogloneta yunnanensis in South China Karst. Animals 2023, 13, 1244. https://doi.org/10.3390/ani13071244
Liu S, Zhou C, Lin Y. New Insights into the Variation and Admixture of the Cave-Dwelling Spider Trogloneta yunnanensis in South China Karst. Animals. 2023; 13(7):1244. https://doi.org/10.3390/ani13071244
Chicago/Turabian StyleLiu, Shiliang, Chuang Zhou, and Yucheng Lin. 2023. "New Insights into the Variation and Admixture of the Cave-Dwelling Spider Trogloneta yunnanensis in South China Karst" Animals 13, no. 7: 1244. https://doi.org/10.3390/ani13071244