
A New Species of the Pythonomorph Carentonosaurus from the Cenomanian of Algora (Guadalajara, Central Spain) †


Abstract
:Simple Summary
Abstract
1. Introduction

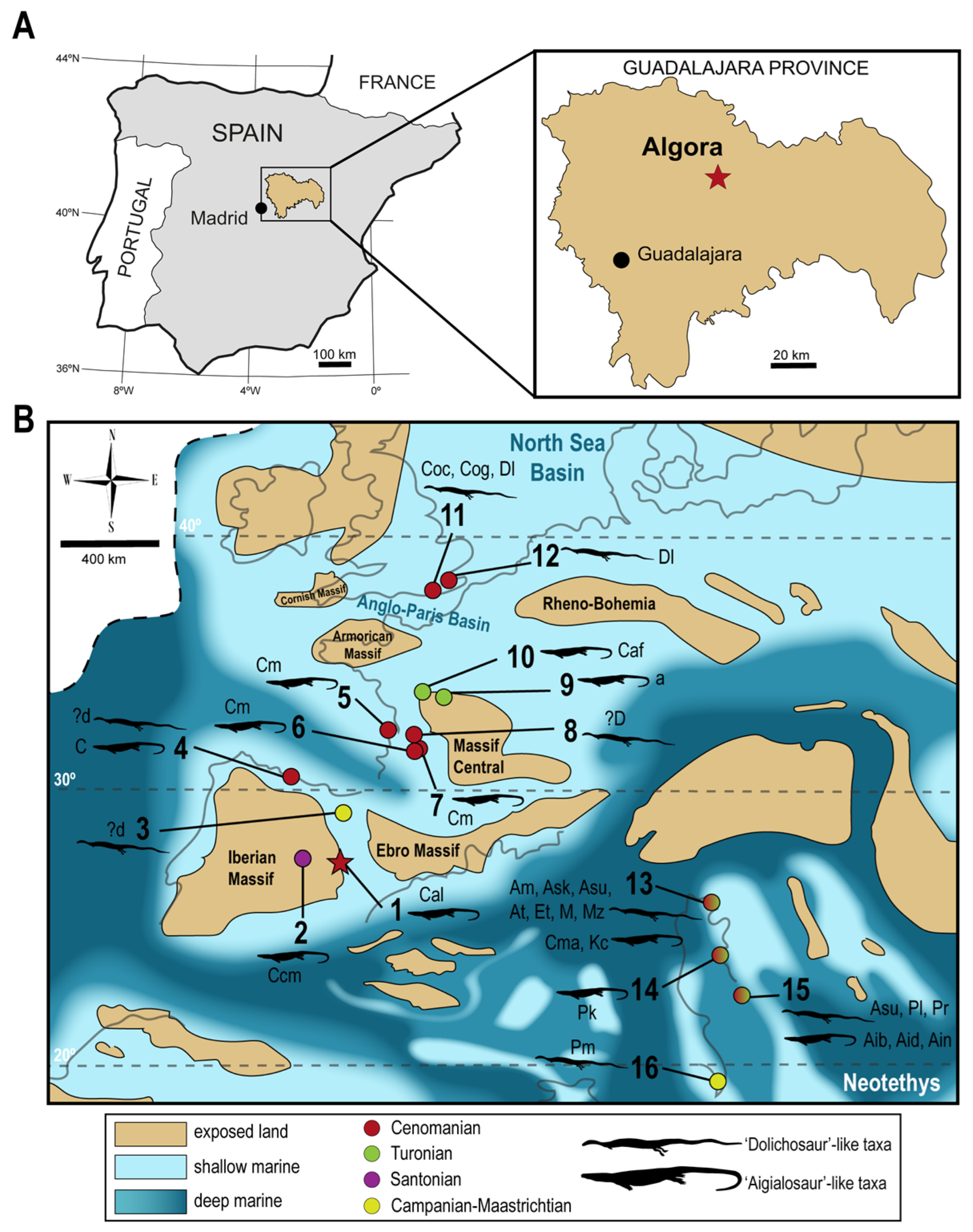
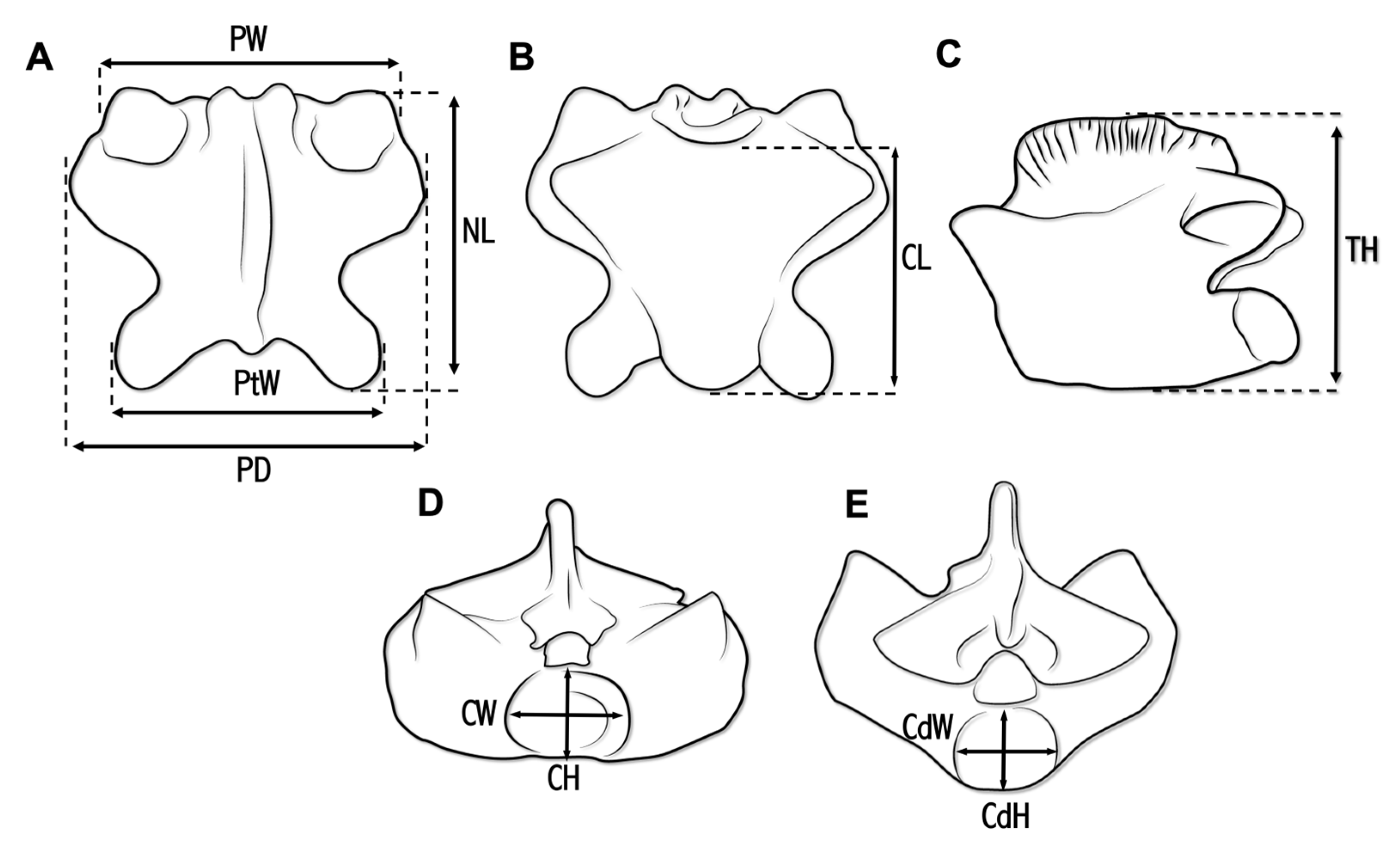
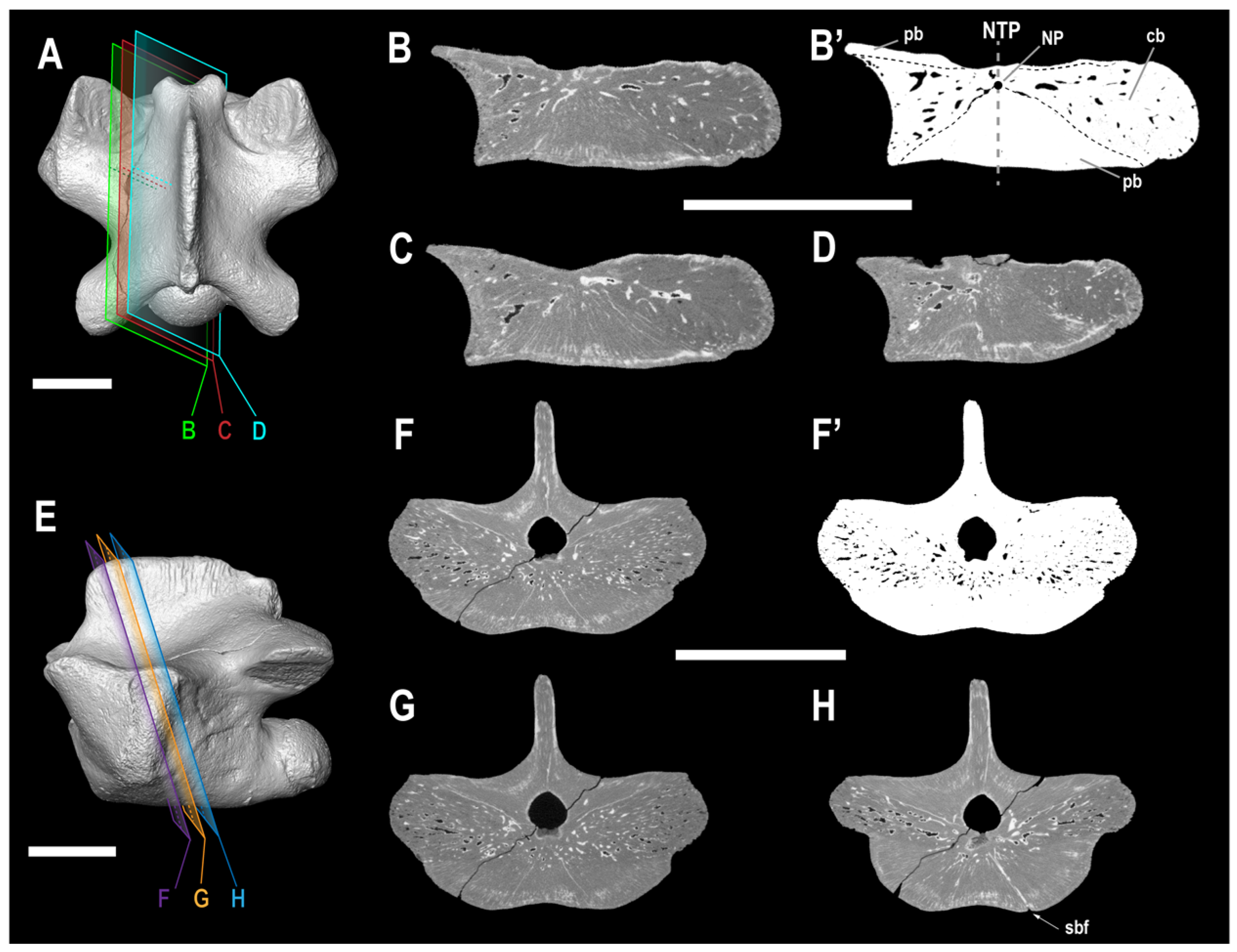
2. Materials and Methods
3. Systematic Paleontology



4. Results
4.1. Anatomical Position of ALG 200 within the Vertebral Column
4.2. Ontogenetic Stage of ALG 200
5. Discussion
5.1. Systematic Assignment of ALG 200
5.1.1. General Considerations
5.1.2. Comparison of ALG 200 with Non-Mosasaurid, Non-Ophidian Pythonomorphs
5.1.3. Systematic Attribution of ALG 200 within the Genus Carentonosaurus
5.2. Paleoecological and Functional Implications for ALG 200
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bardet, N.; Houssaye, A.; Rage, J.-C.; Pereda-Suberbiola, X. The Cenomanian-Turonian (late Cretaceous) radiation of marine squamates (Reptilia): The role of the Mediterranean Tethys. Bull. Soc. Géol. Fr. 2008, 179, 605–622. [Google Scholar] [CrossRef]
- Pérez-García, A.; Bardet, N.; Fregenal-Martínez, M.A.; Martín-Jiménez, M.; Mocho, P.; Narváez, I.; Torices, A.; Vullo, R.; Ortega, F. Cenomanian vertebrates from Algora (central Spain): New data on the establishment of the European Upper Cretaceous continental faunas. Cretac. Res. 2020, 115, 104566. [Google Scholar] [CrossRef]
- Grande, L. An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy. In The Resurrection of Holostei, 6th ed.; American Society of Ichthyologists and Herpetologists, Ed.; Copeia: Lawrence, KS, USA, 2010; pp. 1–871. [Google Scholar]
- Parkinson, C. Upper Greensand and Chloritic Marl, Isle of Wight. Q. J. Geol. Soc. 1881, 37, 370–375. [Google Scholar] [CrossRef]
- Pérez-García, A. A new turtle taxon (Podocnemidoidea, Bothremydidae) reveals the oldest known dispersal event of the crown Pleurodira from Gondwana to Laurasia. J. Syst. Palaeontol. 2017, 15, 709–731. [Google Scholar] [CrossRef]
- Evans, S.E. At the feet of the dinosaurs: The early history and radiation of lizards. Biol. Rev. Camb. Philos. Soc. 2003, 78, 513–551. [Google Scholar] [CrossRef] [PubMed]
- Rage, J.-C. Mesozoic and Cenozoic squamates of Europe. Paleobiodivers Paleoenviron. 2013, 93, 517–534. [Google Scholar] [CrossRef]
- Bolet, A.; Evans, S.E. A new lizard from the Early Cretaceous of Catalonia (Spain), and the Mesozoic lizards of the Iberian Peninsula. Cretac. Res. 2010, 31, 447–457. [Google Scholar] [CrossRef]
- Lee, M.S.Y. The phylogeny of varanoid lizards and the affinities of snakes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1997, 352, 53–91. [Google Scholar] [CrossRef] [Green Version]
- Campbell Mekarski, M.; Pierce, S.E.; Caldwell, M.W. Spatiotemporal Distributions of Non-ophidian Ophidiomorphs, With Implications for Their Origin, Radiation, and Extinction. Front. Earth Sci. 2019, 7, 245. [Google Scholar] [CrossRef] [Green Version]
- Vullo, R.; Bernárdez, E.; Buscalioni, A.D. Vertebrates from the middle?—Late Cenomanian La Cabaña Formation (Asturias, northern Spain): Palaeoenvironmental and palaeobiogeographic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 276, 120–129. [Google Scholar] [CrossRef]
- Pereda-Suberbiola, X.; Corral, J.C.; Astibia, H.; Badiola, A.; Bardet, N.; Berreteaga, A.; Buffetaut, E.; Buscalioni, A.D.; Cappetta, H.; Cavin, L.; et al. Late Cretaceous continental and marine vertebrate assemblages from the Laño quarry (Basque- Cantabrian Region, Iberian Peninsula): An update. J. Iber. Geol. 2015, 41, 101–124. [Google Scholar] [CrossRef] [Green Version]
- Paparella, I.; Palci, A.; Nicosia, U.; Caldwell, M.W. A new fossil marine lizard with soft tissues from the Late Cretaceous of southern Italy. R. Soc. Open Sci. 2018, 5, 172411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Buffrénil, V.; Bardet, N.; Pereda-Suberbiola, X.; Bouya, B. Specialisation of bone structure in Pachyvaranus crassispondylus Arambourg, 1952, an aquatic squamate from the Late Cretaceous of the southern Tethyan margin. Lethaia 2008, 41, 59–69. [Google Scholar] [CrossRef]
- Houssaye, A.; de Buffrénil, V.; Rage, J.-C.; Bardet, N. An analysis of vertebral ‘pachyostosis’ in Carentonosaurus mineaui (Mosasauroidea, Squamata) from the Cenomanian (early Late Cretaceous) of France, with comments on its phylogenetic and functional significance. J. Vertebr. Paleontol. 2008, 28, 685–691. [Google Scholar] [CrossRef]
- Houssaye, A. A preliminary report on the evolution of the vertebral microanatomy within mosasauroids (Reptilia, Squamata). In Proceedings of the Second Mosasaur Meeting, Special Issue 3; Everhart, M.J., Ed.; Sternberg Museum of Natural History, Fort Hays State University: Hays, KS, USA, 2008; pp. 81–89. [Google Scholar]
- Houssaye, A. “Pachyostosis” in aquatic amniotes: A review. Integr. Zool. 2009, 4, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Houssaye, A. Palaeoecological and morphofunctional interpretation of bone mass increase: An example in Late Cretaceous shallow marine squamates. Biol. Rev. 2012, 88, 117–139. [Google Scholar] [CrossRef] [PubMed]
- Janetschke, N.; Wilmsen, M. Sequence stratigraphy of the lower Upper Cretaceous Elbtal Group (Cenomanian–Turonian of Saxony, Germany). Z. Der Dtsch. Ges. Fur Geowiss. 2014, 165, 179–207. [Google Scholar] [CrossRef]
- Hontecillas, D.; Houssaye, A.; Laplana, C.; Sevilla, P.; Arsuaga, J.L.; Pérez-González, A.; Baquedano, E.; Knoll, F. Reworked marine pythonomorph (Reptilia, Squamata) remains in Late Pleistocene cave deposits in central Spain. Cretac. Res. 2015, 54, 188–202. [Google Scholar] [CrossRef]
- Rage, J.-C.; Néraudeau, D. A new pachyostotic squamate reptile from the Cenomanian of France. Palaeontology 2004, 47, 1195–1210. [Google Scholar] [CrossRef]
- Vullo, R.; Rage, J.-C.; Néraudeau, D. Anuran and squamate remains from the Cenomanian (Late Cretaceous) of Charentes, western France. J. Vertebr. Paleontol. 2011, 31, 279–291. [Google Scholar] [CrossRef]
- Bardet, N.; Pereda Suberbiola, X.; Metais, E. Un lézard varanoïde (Squamata, Mosasauroidea) dans le Crétacé supérieur de Touraine—A varanoid lizard (Squamata, Mosasauroidea) from the Upper Cretaceous of Touraine. Geol. Fr. 1998, 1, 69–72. [Google Scholar]
- Houssaye, A. A new aquatic pythonomorph (Reptilia, Squamata) from the Turonian (Late Cretaceous) of France. C. R. Palevol 2010, 9, 39–45. [Google Scholar] [CrossRef]
- Caldwell, M.W. Description and phylogenetic relationships of a new species of Coniasaurus Owen, 1850 (Squamata). J. Vertebr. Paleontol. 1999, 19, 438–455. [Google Scholar] [CrossRef]
- Caldwell, M.W. On the aquatic squamate Dolichosaurus longicollis Owen, 1850 (Cenomanian, Upper Cretaceous), and the evolution of elongate necks in Squamates. J. Vertebr. Paleontol. 2000, 20, 720–735. [Google Scholar] [CrossRef]
- Caldwell, M.W.; Cooper, J. Redescription, palaeobiogeography, and palaeoecology of Coniasaurus crassidens Owen, 1850 (Squamata) from the English Chalk (Cretaceous; Cenomanian). Zool. J. Linn. Soc. 1999, 127, 423–452. [Google Scholar] [CrossRef]
- Nopcsa, F.B. Eidolosaurus und Pachyophis, Zwei neue Neocom-Reptilien. Palaeontographica 1923, 65, 97–154. [Google Scholar]
- Carroll, R.L.; Debraga, M. Aigialosaurs: Mid-Cretaceous varanoid lizards. J. Vertebr. Paleontol. 1992, 12, 66–86. [Google Scholar] [CrossRef]
- Caldwell, M.W.; Carroll, R.L.; Kaiser, H. The pectoral girdle and forelimb of Carsosaurus marchesetti (Aigialosauridae), with a preliminary phylogenetic analysis of mosasauroids and varanoids. J. Vertebr. Paleontol. 1995, 15, 516–531. [Google Scholar] [CrossRef]
- Lee, M.S.Y.; Caldwell, M.W. Adriosaurus and the Affinities of Mosasaurs, Dolichosaurs, and Snakes. J. Paleontol. 2000, 74, 915–937. [Google Scholar] [CrossRef]
- Lee, M.S.Y.; Scanlon, J.D. Snake phylogeny based on osteology, soft anatomy and ecology. Biol. Rev. 2002, 77, 333–401. [Google Scholar] [CrossRef]
- Caldwell, M.W.; Lee, M.S.Y. Reevaluation of the Cretaceous marine lizard Acteosaurus crassicostatus Calligaris, 1993. J. Paleontol. 2004, 78, 617–619. [Google Scholar] [CrossRef]
- Caldwell, M.W.; Palci, A. A new basal mosasauroid from the Cenomanian (U Cretaceous) of Slovenia with a review of mosasauroid phylogeny and evolution. J. Vertebr. Paleontol. 2007, 27, 863–880. [Google Scholar] [CrossRef]
- Caldwell, M.W.; Palci, A. A new species of marine ophidiomorph lizard, Adriosaurus skrbinensis, from the Upper Cretaceous of Slovenia. J. Vertebr. Paleontol. 2010, 30, 747–755. [Google Scholar] [CrossRef]
- Palci, A.; Caldwell, M.W. Vestigial forelimbs and axial elongation in a 95 millionyear- old non-snake squamate. J. Vertebr. Paleontol. 2007, 27, 1–7. [Google Scholar] [CrossRef]
- Palci, A.; Caldwell, M.W. Redescription of Acteosaurus tommasinii von Meyer, 1860, and a discussion of evolutionary trends within the clade Ophidiomorpha. J. Vertebr. Paleontol. 2010, 30, 94–108. [Google Scholar] [CrossRef]
- Mekarski, M.M. The Origin and Evolution of Aquatic Adaptations in Cretaceous Squamates. Ph.D. Thesis, Department of Biological Sciences, University of Alberta, Edmonton, AB, Canada, 2017. [Google Scholar] [CrossRef]
- Mekarski, M.C.; Japundžić, D.; Krizmanić, K.; Caldwell, M.W. Description of a new basal mosasauroid from the Late Cretaceous of Croatia, with comments on the evolution of the mosasauroid forelimb. J. Vertebr. Paleontol. 2019, 39, e1577872. [Google Scholar] [CrossRef]
- Pierce, S.E.; Caldwell, M.W. Redescription and phylogenetic position of the Adriatic (Upper Cretaceous; Cenomanian) dolichosaur Pontosaurus lesinensis (Kornhuber, 1873). J. Vertebr. Paleontol. 2004, 24, 373–386. [Google Scholar] [CrossRef]
- Dutchak, A.R.; Caldwell, M.W. Redescription of Aigialosaurus dalmaticus Kramberger, 1892, a Cenomanian mosasauroid lizard from Hvar Island, Croatia. Can. J. Earth. Sci. 2006, 43, 1821–1834. [Google Scholar] [CrossRef]
- Dutchak, A.R.; Caldwell, M.W. A redescription of Aigialosaurus (= Opetiosaurus) bucchichi (Kornhuber, 1901) (Squamata: Aigialosauridae) with comments on mosasauroid systematics. J. Vertebr. Paleontol. 2009, 29, 437–452. [Google Scholar] [CrossRef]
- Houssaye, A.; Bardet, N.; Rage, J.-C.; Pereda Suberbiola, X.; Bouya, B.; Amaghzaz, M.; Amalik, M. A review of Pachyvaranus crassispondylus Arambourg, 1952, a pachyostotic marine squamate from the latest Cretaceous phosphates of Morocco and Syria. Geol. Mag. 2011, 148, 237–249. [Google Scholar] [CrossRef]
- Gônet, J.; Laurin, M.; Girondot, M. BoneProfileR: The next step to quantify, model, and statistically compare bone section compactness profiles. Palaeontol. Electron. 2022, 25, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Oppel, N.M. Die Ordnunugen, Familien, und Gattungen der Reprtilien. Munchen 1811, 1811, 1–87. [Google Scholar] [CrossRef] [Green Version]
- Fürbringer, M. Beitrag zur Systematik und Genealogie der Reptilien. Jena. Z. Naturw. 1900, 27, 597–685. [Google Scholar]
- Cope, E.D. On the reptilian order Pythonomorpha and Streptosauria. Proc. Boston Soc. Nat. Hist. 1869, 12, 250–261. [Google Scholar]
- Houssaye, A.; Mazurier, A.; Herrel, A.; Volpato, V.; Tafforeau, P.; Boistel, R.; de Buffrénil, V. Vertebral microanatomy in squamates: Structure, growth and ecological correlates. J. Anat. 2010, 217, 715–727. [Google Scholar] [CrossRef]
- Houssaye, A.; Bardet, N. A baby mosasauroid (Reptilia, Squamata) from the Turonian of Morocco—Tethysaurus ‘junior’ discovered? Cretac. Res. 2013, 46, 208–215. [Google Scholar] [CrossRef]
- Scanlon, J.D.; Lee, M.S.Y.; Archer, M. Mid-Tertiary elapid snakes (Squamata, Colubroidea) from Riversleigh, northern Australia: Early steps in a continent-wide adaptive radiation. Geobios 2003, 36, 573–601. [Google Scholar] [CrossRef]
- Estes, R.; de Queiroz, K.; Gauthier, J. Phylogenetic relationships within Squamata. In Phylogenetic Relationships of the Lizard Families: Essays Commemorating Charles L. Camp; Estes, R., Pregill, G., Eds.; Standford Univ. Press: Stanford, CA, USA, 1988; pp. 119–281. [Google Scholar]
- Houssaye, A.; Bardet, N.; Narváez, I.; Ortega, F. Squamate finding in “Lo Hueco” (Late Campanian-Early Maastrichtian, Cuenca Province, Spain): The second non-marine pythonomorph lizard. Paläontologische Z. 2013, 87, 415–422. [Google Scholar] [CrossRef]
- Houssaye, A.; Rage, J.-C.; Torcida Fernández-Baldor, F.; Huerta, P.; Bardet, N.; Pereda Suberbiola, X. A new varanoid squamate from the Early Cretaceous (Barremian–Aptian) of Burgos, Spain. Cretac. Res. 2013, 41, 127–135. [Google Scholar] [CrossRef]
- DeMar, D.G., Jr.; Jones, M.E.H.; Carrano, M.T. A nearly complete skeleton of a new eusphenodontian from the Upper Jurassic Morrison Formation, Wyoming, USA, provides insight into the evolution and diversity of Rhynchocephalia (Reptilia: Lepidosauria). J. Syst. Palaeontol. 2022, 20, 2093139. [Google Scholar] [CrossRef]
- Ivanov, M.; Vasilyan, D.; Böhme, M.; Zazhigin, V.S. Miocene snakes from northeastern Kazakhstan: New data on the evolution of snake assemblages in Siberia. Hist. Biol. 2018, 31, 1–20. [Google Scholar] [CrossRef]
- Garberoglio, F.F.; Gómez, R.O.; Apesteguía, S.; Caldwell, M.W.; Sánchez, M.L.; Veiga, G. A new specimen with skull and vertebrae of Najash rionegrina (Lepidosauria: Ophidia) from the early Late Cretaceous of Patagonia. J. Syst. Palaeontol. 2019, 17, 1533–1550. [Google Scholar] [CrossRef]
- Georgalis, G.L.; Arca, M.; Rook, L.; Tuveri, C. A new colubroid snake (Serpentes) from the early Pleistocene of Sardinia, Italy. Boll. Soc. Paleontol. Ital. 2019, 58, 277–294. [Google Scholar] [CrossRef]
- Georgalis, G.L.; Villa, A.; Ivanov, M.; Vasilyan, D.; Delfino, M. Fossil amphibians and reptiles from the Neogene locality of Maramena (Greece), the most diverse European herpetofauna at the Miocene/Pliocene transition boundary. Palaeontol. Electron. 2019, 22, 1–99. [Google Scholar] [CrossRef] [PubMed]
- Georgalis, G.L.; Rabi, M.; Smith, K.T. Taxonomic revision of the snakes of the genera Palaeopython and Paleryx (Serpentes, Constrictores) from the Paleogene of Europe. Swiss. J. Palaeontol. 2021, 140, 1–140. [Google Scholar] [CrossRef]
- Palci, A. On the Origin and Evolution of the Ophidia. Ph.D. Thesis, Department of Biological Sciences, University of Alberta, Edmonton, AB, Canada, 2014. [Google Scholar] [CrossRef]
- Gauthier, J.A.; Kearney, M.; Maisano, J.A.; Rieppel, O.; Behlke, A.D.B. Assembling the squamate tree of life: Perspectives from the phenotype and the fossil record. Bull. Peabody Mus. Nat. Hist. 2012, 53, 3–308. [Google Scholar] [CrossRef]
- Martill, D.M.; Tischlinger, H.; Longrich, N.R. A four-legged snake from the Early Cretaceous of Gondwana. Science 2015, 349, 416–419. [Google Scholar] [CrossRef] [Green Version]
- Bell, G.L.; Polcyn, M.J. Dallasaurus turneri, a new primitive mosasauroid from the Middle Turonian of Texas and comments on the phylogeny of Mosasauridae (Squamata). Neth. J. Geosci. 2005, 84, 177–194. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, J.; Caldwell, M.W.; Jagt, J.W.M. New data on the postcranial anatomy of the California mosasaur Plotosaurus bennisoni (Camp, 1942) (Upper Cretaceous: Maastrichtian), and the taxonomic status of P. tuckeri (Camp, 1942). J. Vertebr. Paleontol. 2008, 28, 1043–1054. [Google Scholar] [CrossRef]
- Bullard, T.S. Anatomy and systematics of North American tylosaurine mosasaurs. Ph.D. Thesis, Department of Biological Sciences, University of Alberta, Edmonton, AB, Canada, 2006. [Google Scholar]
- Makádi, L.; Caldwell, M.W.; Ősi, A.; Butler, R.J. The First Freshwater Mosasauroid (Upper Cretaceous, Hungary) and a New Clade of Basal Mosasauroids. PLoS ONE 2012, 7, e51781. [Google Scholar] [CrossRef] [Green Version]
- Bardet, N.; Suberbiola, X.P.; Jalil, N.-E. A new mosasauroid (Squamata) from the Late Cretaceous (Turonian) of Morocco. C. R. Palevol 2003, 2, 607–616. [Google Scholar] [CrossRef]
- Augusta, B.; Zaher, H.; Polcyn, M.; Fiorillo, A.; Jacobs, L. A Review of Non-Mosasaurid (Dolichosaur and Aigialosaur) Mosasaurians and Their Relationships to Snakes. In The Origin and Early Evolutionary History of Snakes, Systematics Association Special Volume Series; Gower, D., Zaher, H., Eds.; Cambridge University Press: Cambridge, UK, 2022; pp. 157–179. [Google Scholar] [CrossRef]
- Reeder, T.W.; Townsend, T.M.; Mulcahy, D.G.; Noonan, B.P.; Wood, P.L., Jr.; Sites, J.W., Jr.; Wiens, J.J. Integrated Analyses Resolve Conflicts over Squamate Reptile Phylogeny and Reveal Unexpected Placements for Fossil Taxa. PLoS ONE 2015, 10, e0118199. [Google Scholar] [CrossRef] [Green Version]
- Madzia, D.; Cau, A. Inferring `weak spots’ in phylogenetic trees: Application to mosasauroid nomenclature. PeerJ 2017, 5, e3782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, S.E.; Manabe, M.; Noro, M.; Isaji, S.; Yamaguchi, M. A long-bodied lizard from the Lower Cretaceous of Japan. Palaeontology 2006, 49, 1143–1165. [Google Scholar] [CrossRef]
- Dal Sasso, C.; Pinna, G. Aphanizocnemus libanensis n. gen. n. sp., a new dolichosaur (Reptilia, Varanoidea) from the Upper Cretaceous of Lebanon. Paleontol. Lomb. 1997, 7, 1–31. [Google Scholar]
- Kornhuber, A. Carsosaurus marchesettii, a new fossil lacertilian from the Cretaceous formation of Karst near Komen. Abh Geol Reichsanst. Wien 1893, 17, 1–15. [Google Scholar]
- Owen, R. Description of the fossil reptiles of the Chalk Formation. In The Geology and Fossils of the Tertiary and Cretaceous Formations of Sussex; Dixon, F., Ed.; Longman, Brown, Green and Longmans: London, UK, 1850; pp. 378–400, 403–404. [Google Scholar]
- Bell, B.A.; Murry, P.A.; Osten, L.W. Coniasaurus Owen, 1850 from North America. J. Paleontol. 1982, 56, 520–524. [Google Scholar]
- Shimada, K.; Bell, G.L., Jr. Coniasaurus Owen, 1850 (Reptilia: Squamata), from the Upper Cretaceous Niobrara Chalk of western Kansas. J. Paleontol. 2006, 80, 589–593. [Google Scholar] [CrossRef]
- Shimada, K.; Schumacher, B.A.; Parkin, J.A.; Palermo, J.M. Fossil marine vertebrates from the lowermost Greenhorn Limestone (Upper Cretaceous: Middle Cenomanian) in southeastern Colorado. J. Paleontol. 2006, 63, 1–45. [Google Scholar] [CrossRef]
- Shimada, K.; Ystesund, T.K. A dolichosaurid lizard, Coniasaurus cf C. crassidens, from the Upper Cretaceous Carlile Shale in Russell County, Kansas. Trans. Kans. Acad. Sci. 2007, 110, 236–242. [Google Scholar] [CrossRef]
- Guzzo, F.; Shimada, K. A New Fossil Vertebrate Locality of the Jetmore Chalk Member of the Upper Cretaceous Greenhorn Limestone in North-Central Kansas, U.S.A. Trans. Kans. Acad. Sci. 2018, 121, 59–68. [Google Scholar] [CrossRef]
- Scanlon, J.; Hocknull, S.A. A dolichosaurid lizard from the latest Albian (mid-Cretaceous) Winton Formation, Queensland, Australia. In Proceedings of the Second Mosasaur Meeting, Special Issue 3; Everhart, M.J., Ed.; Sternberg Museum of Natural History, Fort Hays State University: Hays, KS, USA, 2008; pp. 131–136. [Google Scholar]
- von Meyer, H. Acteosaurus tommasinii aus dem schwarzen Kreide-Schiefer von Comen am Karste. Palaeontogr. Beiträge Zur Nat. Der Vorwelt 1860, 7, 223–231. [Google Scholar]
- Cornalia, E.; Chiozza, L. Cenni geologici sull’ Istria. G. Dell’ I. R. Inst. Lomb. 1852, 3, 1–35. [Google Scholar]
- Kramberger, C.G. Aigialosaurus: Ein neue Eidechse a.d. Kreideschifern der Insel Lesina mit Rücksicht auf die bereits beschrieben Lacertiden von Comen und Lesina. Glas. Hrvat. Narav. Druz. Societas Historico-Naturalis Croatica U Zagrebu. 1892, 7, 74–106. [Google Scholar]
- Caldwell, M.W. A new species of Pontosaurus (Squamata, Pythonomorpha) from the Upper Cretaceous of Lebanon and a phylogenetic analysis of Pythonomorpha. Mem. Soc. Ital. Scien. Natur. Mus. Civ. 2006, 34, 1–42. [Google Scholar]
- Polcyn, M.J.; Tchernov, E.; Jacobs, L.L. The Cretaceous biogeography of the Eastern Mediterranean with a description of a new basal mosasauroid fron ’Ein Yabrun, Israel. In Proceedings of the Second Gondwanan Dinosaur Symposium; Tomida, Y., Rich, T.H., Vickers-Rich, P., Eds.; National Science Museum Monographs: Tokyo, Japan, 1999; pp. 259–290. [Google Scholar]
- Polcyn, M.J.; Tchernov, E.; Jacobs, L.L. Haasiasaurus gen. nov., a new generic name for the basal mosasauroid Haasia Polcyn et al., 1999. J. Vertebr. Paleontol. 2003, 23, 476. [Google Scholar] [CrossRef]
- Kornhuber, A. Opetiosaurus bucchichi. Eine neue fossile Eidechse aus der unteren Kreide von Lesina in Dalmatien. Abh. Der Kk Geol. Reichsanst. 1901, 17, 1–24. [Google Scholar]
- Sood, M.S. The anatomy of the vertebral column in Serpentes. Proc. Indian Acad. Sci. 1948, 28, 1–26. [Google Scholar] [CrossRef]
- Rage, J.-C.; Vullo, R.; Néraudeau, D. The mid-Cretaceous snake Simoliophis rochebrunei Sauvage, 1880 (Squamata: Ophidia) from its type area (Charentes, southwestern France): Redescription, distribution, and palaeoecology. Cretac. Res. 2016, 58, 234–253. [Google Scholar] [CrossRef]
- Houssaye, A.; Sander, P.M.; Klein, N. Adaptive Patterns in Aquatic Amniote Bone Microanatomy—More Complex than Previously Thought. Integr. Comp. Biol. 2016, 56, 1349–1369. [Google Scholar] [CrossRef] [Green Version]
- Houssaye, A.; de Buffrénil, V. Chapter 36: Bone histology and the adaptation to aquatic life in tetrapods. In Vertebrate Skeletal Histology and Paleohistology; de Buffrénil, V., de Ricqlès, A., Zylberberg, L., Padian, K., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 744–754. [Google Scholar] [CrossRef]
- de Buffrénil, V.; Canoville, A.; D’Anastasio, R.; Domning, D.P. Evolution of Sirenian Pachyosteosclerosis, a Modelcase for the Study of Bone Structure in Aquatic Tetrapods. J. Mamm. Evol. 2010, 17, 101–120. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | PW | PtW | PD | NL | CL | MH | CW | CH | CdW | CdH |
|---|---|---|---|---|---|---|---|---|---|---|
| Carentonosaurus algorensis sp. nov. | 16 | 14.5 | 20 | 17 | 13 | 14 | 7 | 5.5 | 5 | 4 |
| Carentonosaurus mineaui | 11.7 * | 10.3 | 14.1 * | 11.2 | 9.3 * | 12.3 | 5 | 3.4 | 4.3 * | 3 |
| 1 | 2 | 3 | 4 | 5 | 6 | Mean | |
|---|---|---|---|---|---|---|---|
| ImageJ | 97.11 | 96.91 | 96.99 | 95.94 | 95.96 | 97.53 | 96.74 |
| BoneProfileR | 93.80 | 93.50 | 93.75 | 96.00 | 96.10 | 96.10 | 94.88 |
| Section mean | 95.46 | 95.21 | 95.37 | 95.97 | 96.03 | 96.82 | 95.81 |
| Mean | 95.34 | 96.27 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabezuelo-Hernández, A.; Pérez-García, A. A New Species of the Pythonomorph Carentonosaurus from the Cenomanian of Algora (Guadalajara, Central Spain). Animals 2023, 13, 1197. https://doi.org/10.3390/ani13071197
Cabezuelo-Hernández A, Pérez-García A. A New Species of the Pythonomorph Carentonosaurus from the Cenomanian of Algora (Guadalajara, Central Spain). Animals. 2023; 13(7):1197. https://doi.org/10.3390/ani13071197
Chicago/Turabian StyleCabezuelo-Hernández, Alberto, and Adán Pérez-García. 2023. "A New Species of the Pythonomorph Carentonosaurus from the Cenomanian of Algora (Guadalajara, Central Spain)" Animals 13, no. 7: 1197. https://doi.org/10.3390/ani13071197