
Spatial Overlap and Habitat Selection of Corvid Species in European Cities
 , , , , , ,
, , , , , ,  , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
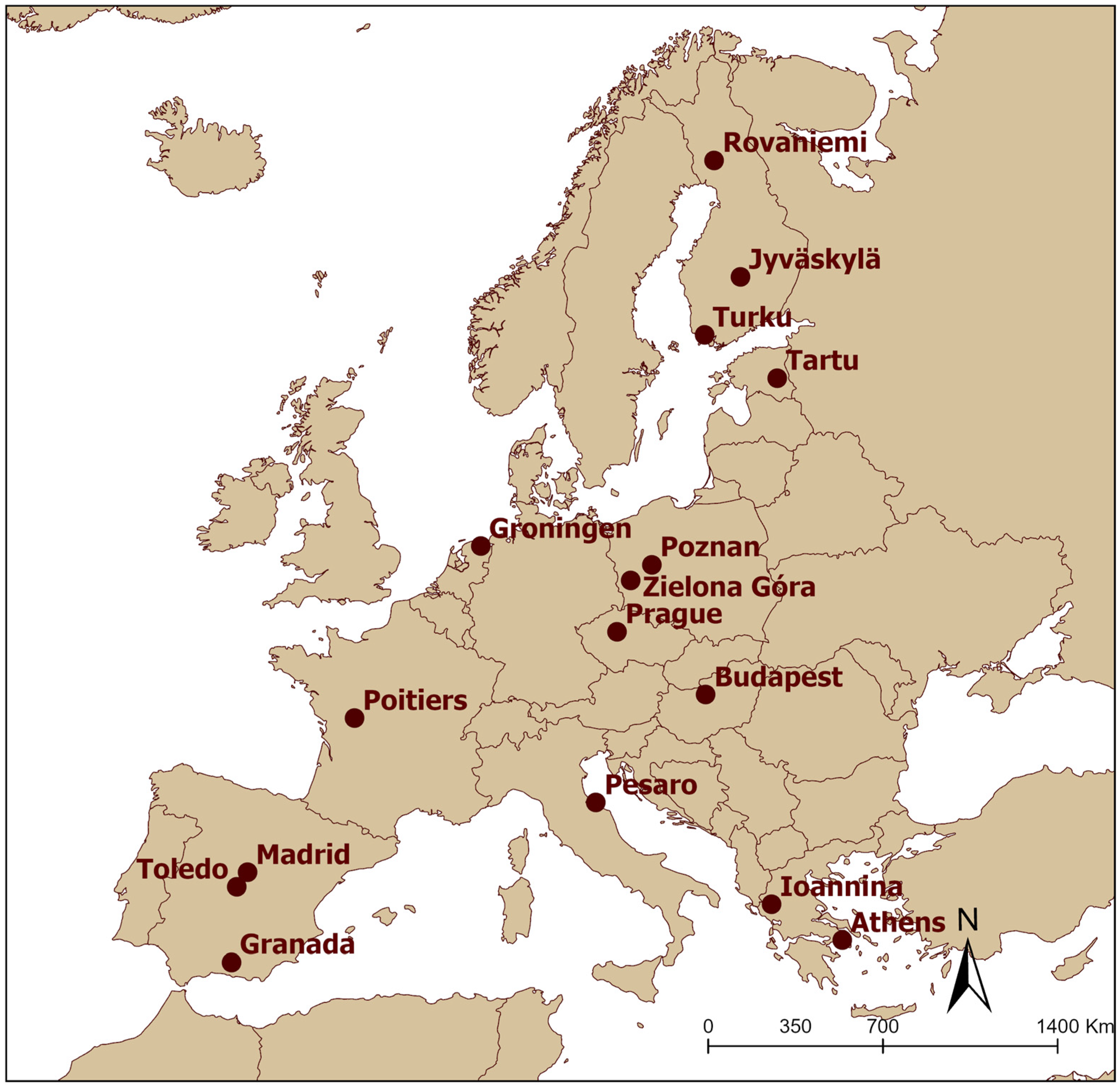
2.1. Study Area
2.2. Field Data Collection
2.3. Variables Studied in Terms of Corvid Habitat Selection
2.4. Classification of the Carrion Crow and the Hooded Crow
2.5. Statistical Analyses
3. Results
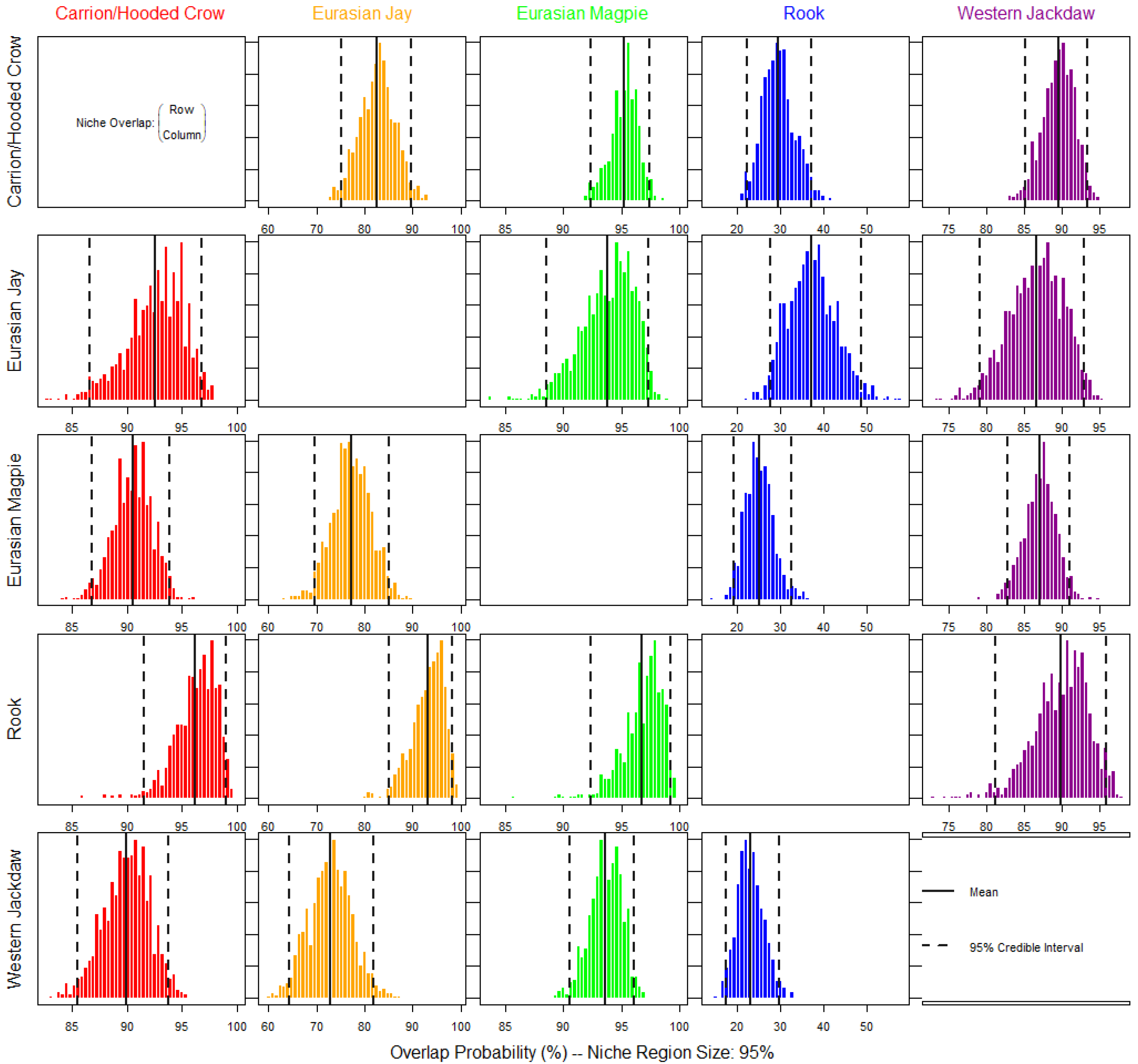
3.1. Spatial Overlap
3.2. Habitat Selection
4. Discussion
4.1. Corvids’ Abundance and Distribution in Urban Areas
4.2. Corvids’ Urban Habitat Selection
4.3. Corvids’ Spatial and Habitat Overlap
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McDonald, R.I.; Mansur, A.V.; Ascensão, F.; Colbert, M.; Crossman, K.; Elmqvist, T.; Gonzalez, A.; Güneralp, B.; Haase, D.; Hamann, M.; et al. Research Gaps in Knowledge of the Impact of Urban Growth on Biodiversity. Nat. Sustain. 2020, 3, 16–24. [Google Scholar] [CrossRef]
- Center for International Earth Science Information Network—CIESIN; Columbia University; International Food Policy Research Institute—IFPRI; The World Bank; Centro Internacional de Agricultura Tropical—CIAT. Global Rural-Urban Mapping Project, Version 1 (GRUMPv1): Urban Extents Grid; Palisades; NASA Socioeconomic Data and Applications Center (SEDAC): New York, NY, USA, 2011. [Google Scholar] [CrossRef]
- United Nations—Department of Economic and Social Affairs—Population Division. World Urbanization Prospects: The 2018 Revision; United Nations: New York, NY, USA, 2019; ISBN 9789211483192. [Google Scholar]
- Alberti, M.; Correa, C.; Marzluff, J.M.; Hendry, A.P.; Palkovacs, E.P.; Gotanda, K.M.; Hunt, V.M.; Apgar, T.M.; Zhou, Y. Global Urban Signatures of Phenotypic Change in Animal and Plant Populations. Proc. Natl. Acad. Sci. USA 2017, 114, 8951–8956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global Change and the Ecology of Cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prugh, L.R.; Stoner, C.J.; Epps, C.W.; Bean, W.T.; Ripple, W.J.; Laliberte, A.S.; Brashares, J.S. The Rise of the Mesopredator. Bioscience 2009, 59, 779–791. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a Major Cause of Biotic Homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Clergeau, P.; Croci, S.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.L.; Dinetti, M. Avifauna Homogenisation by Urbanisation: Analysis at Different European Latitudes. Biol. Conserv. 2006, 127, 336–344. [Google Scholar] [CrossRef]
- McKinney, M.L.; Lockwood, J.L. Biotic Homogenization: A Few Winners Replacing Many Losers in the next Mass Extinction. Trends Ecol. Evol. 1999, 14, 450–453. [Google Scholar] [CrossRef]
- Garcia-Porta, J.; Sol, D.; Pennell, M.; Sayol, F.; Kaliontzopoulou, A.; Botero, C.A. Niche Expansion and Adaptive Divergence in the Global Radiation of Crows and Ravens. Nat. Commun. 2022, 13, 2086. [Google Scholar] [CrossRef]
- Matsyura, A.V.; Zimaroyeva, A.A.; Jankowski, K. Spatial Patterns of Seasonal Distribution of Corvidae (the Case of Urban Habitats). Biosyst. Divers. 2016, 24, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Preininger, D.; Schoas, B.; Kramer, D.; Boeckle, M. Waste Disposal Sites as All-You-Can Eat Buffets for Carrion Crows (Corvus corone). Animals 2019, 9, 215. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, S.; Jiang, P.; Ding, P. Black-Billed Magpies (Pica pica) Adjust Nest Characteristics to Adapt to Urbanization in Hangzhou, China. Can. J. Zool. 2008, 86, 676–684. [Google Scholar] [CrossRef]
- Benmazouz, I.; Jokimäki, J.; Lengyel, S.; Juhász, L.; Kaisanlahti-Jokimäki, M.-L.; Kardos, G.; Paládi, P.; Kövér, L. Corvids in Urban Environments: A Systematic Global. Animals 2021, 11, 3226. [Google Scholar] [CrossRef]
- Seed, A.; Emery, N.; Clayton, N. Intelligence in Corvids and Apes: A Case of Convergent Evolution? Ethology 2009, 115, 401–420. [Google Scholar] [CrossRef]
- Sol, D.; Duncan, R.P.; Blackburn, T.M.; Cassey, P.; Lefebvre, L. Big Brains, Enhanced Cognition, and Response of Birds to Novel Environments. Proc. Natl. Acad. Sci. USA 2005, 102, 5460–5465. [Google Scholar] [CrossRef] [Green Version]
- Božič, L. Numbers, Distribution and Nest Site Characteristics of Jackdaw Corvus monedula in Slovenia and Its Conservation Status. Acrocephalus 2016, 37, 123–150. [Google Scholar] [CrossRef] [Green Version]
- Szala, K.; Dylewski, Ł.; Tobolka, M. Winter Habitat Selection of Corvids in an Urban Ecosystem. Urban Ecosyst. 2020, 23, 483–493. [Google Scholar] [CrossRef] [Green Version]
- Jokimäki, J.; Kaisanlahti-Jokimäki, M.L.; Suhonen, J. Long-Term Winter Population Trends of Corvids in Relation to Urbanization and Climate at Northern Latitudes. Animals 2022, 12, 1820. [Google Scholar] [CrossRef]
- Belaire, J.A.; Westphal, L.M.; Whelan, C.J.; Minor, E.S. Urban Residents’ Perceptions of Birds in the Neighborhood: Biodiversity, Cultural Ecosystem Services, and Disservices. Condor 2015, 117, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Julian, K.G.; Eidson, M.; Kipp, A.M.; Weiss, E.; Petersen, L.R.; Miller, J.R.; Hinten, S.R.; Marfin, A.A. Early Season Crow Mortality as a Sentinel for West Nile Virus Disease in Humans, Northeastern United States. Vector Borne Zoonotic Dis. 2002, 2, 145–155. [Google Scholar] [CrossRef]
- Pesendorfer, M.B.; Sillett, T.S.; Koenig, W.D.; Morrison, S.A. Scatter-Hoarding Corvids as Seed Dispersers for Oaks and Pines: A Review of a Widely Distributed Mutualism and Its Utility to Habitat Restoration. Condor 2016, 118, 215–237. [Google Scholar] [CrossRef]
- Jiguet, F. The Fox and the Crow. A Need to Update Pest Control Strategies. Biol. Conserv. 2020, 248, 108693. [Google Scholar] [CrossRef] [PubMed]
- Yoda, T. Managing Urban Crow Populations in Japan. Hum.-Wildl. Interact. 2019, 13, 439–446. [Google Scholar] [CrossRef]
- Evans, K.L.; Newson, S.E.; Gaston, K.J. Habitat Influences on Urban Avian Assemblages. Ibis 2009, 151, 19–39. [Google Scholar] [CrossRef]
- Jokimäki, J.; Kaisanlahti-Jokimäki, M.; Sorace, A.; Fernández-Juricic, E.; Rodriguez-Prieto, I.; Jimenez, M.D. Evaluation of the “Safe Nesting Zone” Hypothesis across an Urban Gradient: A Multi-scale Study. Ecography 2005, 28, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Jokimäki, J.; Huhta, E. Artificial Nest Predation and Abundance of Birds Along an Urban Gradient. Condor 2000, 102, 838–847. [Google Scholar] [CrossRef]
- Madden, C.F.; Arroyo, B.; Amar, A. A Review of the Impacts of Corvids on Bird Productivity and Abundance. Ibis 2015, 157, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Marzluff, J.M.; Neatherlin, E. Corvid Response to Human Settlements and Campgrounds: Causes, Consequences, and Challenges for Conservation. Biol. Conserv. 2006, 130, 301–314. [Google Scholar] [CrossRef]
- Baltensperger, A.P.; Mullet, T.C.; Schmid, M.S.; Humphries, G.R.W.; Kövér, L.; Huettmann, F. Seasonal Observations and Machine-Learning-Based Spatial Model Predictions for the Common Raven (Corvus corax) in the Urban, Sub-Arctic Environment of Fairbanks, Alaska. Polar Biol. 2013, 36, 1587–1599. [Google Scholar] [CrossRef]
- Nakahara, T.; Kuroe, M.; Hasegawa, O.; Hayashi, Y.; Mori, S.; Eguchi, K. Nest Site Characteristics of the Newly Established Eurasian Magpie Pica pica Population in Hokkaido, Japan. Ornithol. Sci. 2015, 14, 99–109. [Google Scholar] [CrossRef]
- Xu, Y.; Cao, Z.; Wang, B. Effect of Urbanization Intensity on Nest-Site Selection by Eurasian Magpies (Pica pica). Urban Ecosyst. 2020, 23, 1099–1105. [Google Scholar] [CrossRef]
- Ciebiera, O.; Czechowski, P.; Morelli, F.; Piekarski, R.; Bocheński, M.; Chachulska-Serweta, J.; Jerzak, L. Selection of Urbanized Areas by Magpie Pica pica in a Medium Size City in Poland. Animals 2021, 11, 1738. [Google Scholar] [CrossRef]
- Jokimäki, J.; Suhonen, J.; Vuorisalo, T.; Kövér, L.; Kaisanlahti-Jokimäki, M.L. Urbanization and Nest-Site Selection of the Black-Billed Magpie (Pica pica) Populations in Two Finnish Cities: From a Persecuted Species to an Urban Exploiter. Landsc. Urban Plan. 2017, 157, 577–585. [Google Scholar] [CrossRef] [Green Version]
- Šálek, M.; Grill, S.; Riegert, J. Nest-Site Selection of an Avian Urban Exploiter, the Eurasian Magpie Pica pica, across the Urban-Rural Gradient. J. Vertebr. Biol. 2020, 70, 20086.1–20086.11. [Google Scholar] [CrossRef]
- Morrison, M.L. The Habitat Sampling and Analysis Paradigm Has Limited Value in Animal Conservation: A Prequel. J. Wildl. Manag. 2012, 76, 438–450. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Concluding Remarks. Cold Spring Harb. Symp. Quant. Biol. 1957, 22, 145–159. [Google Scholar] [CrossRef]
- Whittaker, R.H.; Levin, S.A.; Rootj, R.B. Niche, Habitat, and Ecotope. Am. Nat. 1973, 107, 321–338. [Google Scholar] [CrossRef]
- Marko, P.B. Sympatry. In Encyclopedia of Ecology; Academic Press: Cambridge, MA, USA, 2008; pp. 3450–3458. [Google Scholar] [CrossRef]
- Matsubara, H. Comparative Study of Territoriality and Habitat Use in Syntopic Jungle Crow (Corvus macrorhynchos) and Carrion Crow (C. corone). Ornithol. Sci. 2003, 2, 103–111. [Google Scholar] [CrossRef]
- Waite, R.K. Sympatric Corvids: Effects of Social Behaviour, Aggression and Avoidance on Feeding. Behav. Ecol. Sociobiol. 1984, 15, 55–59. [Google Scholar] [CrossRef]
- Baglione, V.; Canestrari, D. Kleptoparasitism and Temporal Segregation of Sympatric Corvids Foraging in a Refuse Dump. Auk 2009, 126, 566–578. [Google Scholar] [CrossRef]
- Gianpasquale, C.; Alberto, M. The Occurrence and Density of Three Sympatric Corvids in a Mediterranean Agroecosystem Explained by Land Use. J. Ornithol. 2019, 160, 1133–1150. [Google Scholar] [CrossRef]
- Rolando, A.; Giachello, P. Interspecific Coexistence in Corvids in an Alpine Valley of Northwestern Italy. Bolletino Zool. 1992, 59, 281–288. [Google Scholar] [CrossRef]
- Bibby, C.J.; Burgess, N.D.; Hillis, D.M.; Hill, D.A.; Mustoe, S. Bird Census Techniques; Elsevier: Amsterdam, The Netherlands, 1992; ISBN 0120958317. [Google Scholar]
- Voříšek, P.; Klvaňová, A.; Wotton, S.; Gregory, R.D. A Best Practice Guide for Wild Bird Monitoring Schemes; Royal Society for the Protection of Birds: Sandy, UK, 2008. [Google Scholar]
- Liordos, V.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.L.; Valsamidis, E.; Kontsiotis, V.J. Patch, Matrix and Disturbance Variables Negatively Influence Bird Community Structure in Small-Sized Managed Green Spaces Located in Urban Core Areas. Sci. Total Environ. 2021, 801, 149617. [Google Scholar] [CrossRef] [PubMed]
- Suhonen, J.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L.; Morelli, F.; Benedetti, Y.; Rubio, E.; Pérez-Contreras, T.; Sprau, P.; Tryjanowski, P.; Møller, A.P.; et al. Occupancy-Frequency Distribution of Birds in Land-Sharing and-Sparing Urban Landscapes in Europe. Landsc. Urban Plan. 2022, 226, 104463. [Google Scholar] [CrossRef]
- Kéry, M.; Royle, J.A.; Schmid, H. Modeling Avian Abundance from Replicated Counts Using Binomial Mixture Models. Ecol. Appl. 2005, 15, 1450–1461. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Major, R.E.; Lyons, M.B.; Martin, J.M.; Kingsford, R.T. The Effects of Local and Landscape Habitat Attributes on Bird Diversity in Urban Greenspaces. Ecosphere 2018, 9, e02347. [Google Scholar] [CrossRef] [Green Version]
- Chace, J.F.; Walsh, J.J. Urban Effects on Native Avifauna: A Review. Landsc. Urban Plan. 2006, 74, 46–69. [Google Scholar] [CrossRef]
- Liordos, V.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L.; Valsamidis, E.; Kontsiotis, V.J. Niche Analysis and Conservation of Bird Species Using Urban Core Areas. Sustainability 2021, 13, 6327. [Google Scholar] [CrossRef]
- Muñoz-Pedreros, A.; González-Urrutia, M.; Encina-Montoya, F.; Norambuena, H.V. Effects of Vegetation Strata and Human Disturbance on Bird Diversity in Green Areas in a City in Southern Chile. Avian Res. 2018, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- Peris, S.; Montelongo, T. Birds and Small Urban Parks: A Study in a High Plateau City. Turk. J. Zool. 2014, 38, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Amaya-Espinel, J.D.; Hostetler, M.; Henríquez, C.; Bonacic, C. The Influence of Building Density on Neotropical Bird Communities Found in Small Urban Parks. Landsc. Urban Plan. 2019, 190, 103578. [Google Scholar] [CrossRef]
- Miller, S.G.; Knight, R.L.; Miller, C.K. Wildlife Responses to Pedestrians and Dogs. Wildl. Soc. Bull. 2001, 29, 124–132. [Google Scholar]
- Adams, C.A.; Blumenthal, A.; Fernández-Juricic, E.; Bayne, E.; St Clair, C.C. Effect of Anthropogenic Light on Bird Movement, Habitat Selection, and Distribution: A Systematic Map Protocol. Environ. Evid. 2019, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Ciach, M.; Fröhlich, A. Habitat Type, Food Resources, Noise and Light Pollution Explain the Species Composition, Abundance and Stability of a Winter Bird Assemblage in an Urban Environment. Urban Ecosyst. 2017, 20, 547–559. [Google Scholar] [CrossRef] [Green Version]
- Francis, C.D.; Ortega, C.P.; Cruz, A. Noise Pollution Changes Avian Communities and Species Interactions. Curr. Biol. 2009, 19, 1415–1419. [Google Scholar] [CrossRef] [Green Version]
- Morelli, F.; Benedetti, Y.; Ibáñez-Álamo, J.D.; Tryjanowski, P.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L.; Suhonen, J.; Díaz, M.; Møller, A.P.; Moravec, D.; et al. Effects of Urbanization on Taxonomic, Functional and Phylogenetic Avian Diversity in Europe. Sci. Total Environ. 2021, 795, 148874. [Google Scholar] [CrossRef]
- Parkin, D.T.; Collinson, M.; Helbig, A.J.; Knox, A.G.; Sangster, G. The Taxonomic Status of Carrion and Hooded Crows. Br. Birds 2003, 96, 274–290. [Google Scholar]
- Vrezec, A. Historical Occurrence of the Hooded/Carrion Crow (Corvus cornix/corone) in Urban Areas of Europe with Emphasis on Slovenia. Annales Ser. Hist. Nat. 2010, 20, 131–140. [Google Scholar]
- Keller, V.; Herrando, S.; Voríšek, P.; Franch, M.; Kipson, M.; Milanesi, P.; Martí, D.; Anton, M.; Klvanová, A.; Kalyakin, M.V. European Breeding Bird Atlas 2: Distribution, Abundance and Change; Lynx Edicions: Cerdanyola del Vallès, Spain, 2020; ISBN 8416728380. [Google Scholar]
- Pearman, P.B.; Lavergne, S.; Roquet, C.; Wüest, R.; Zimmermann, N.E.; Thuiller, W. Phylogenetic Patterns of Climatic, Habitat and Trophic Niches in a European Avian Assemblage. Glob. Ecol. Biogeogr. 2014, 23, 414–424. [Google Scholar] [CrossRef]
- Storchová, L.; Hořák, D. Life-History Characteristics of European Birds. Glob. Ecol. Biogeogr. 2018, 27, 400–406. [Google Scholar] [CrossRef]
- Metzler, D.; Knief, U.; Peñalba, J.V.; Wolf, J.B.W. Assortative Mating and Epistatic Mating-Trait Architecture Induce Complex Movement of the Crow Hybrid Zone. Evolution 2021, 75, 3154–3174. [Google Scholar] [CrossRef]
- Wolf, J.B.W.; Bayer, T.; Haubold, B.; Schilhabel, M.; Rosenstiel, P.; Tautz, D. Nucleotide Divergence vs. Gene Expression Differentiation: Comparative Transcriptome Sequencing in Natural Isolates from the Carrion Crow and Its Hybrid Zone with the Hooded Crow. Mol. Ecol. 2010, 19, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Knief, U.; Bossu, C.M.; Saino, N.; Hansson, B.; Poelstra, J.; Vijay, N.; Weissensteiner, M.; Wolf, J.B.W. Epistatic Mutations under Divergent Selection Govern Phenotypic Variation in the Crow Hybrid Zone. Nat. Ecol. Evol. 2019, 3, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Mantel, N. The Detection of Disease Clustering and a Generalized Regression Approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]
- Dray, S.; Dufour, A. The Ade4 Package: Implementing the Duality Diagram for Ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Lysy, M.; Stasko, A.D.; Swanson, H.K. Package ‘NicheROVER’ Niche Region and Niche Overlap Metrics for Multidimensional Ecological Niches. 2021. Available online: https://CRAN.R-project.org/package=nicheROVER (accessed on 7 June 2021).
- Swanson, H.K.; Lysy, M.; Power, M.; Stasko, A.D.; Johnson, J.D.; Reist, J. Ecological Niches and Niche Overlap R Eports. Ecology 2015, 96, 318–324. [Google Scholar] [CrossRef]
- Escoriza, D.; Amat, F. Habitat Partitioning and Overlap by Large Lacertid Lizards in Southern Europe. Diversity 2021, 13, 155. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Lme4: Linear Mixed-Effects Models Using Eigen and S4—R Package 2014. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Barton, K. MuMIn: Multi-Model Inference. R Package Version 1.15.1. 2020. Available online: http://CRAN.R-project.org/package=MuMIn (accessed on 7 June 2021).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing Version 4.0.3; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- François, C.; Alexandre, L.; Julliard, R. Effects of Landscape Urbanization on Magpie Occupancy Dynamics in France. Landsc. Ecol. 2008, 23, 527–538. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Jerzak, L.; Zhang, Z. Nest Site Selection of the Magpie Pica pica Sericea in Beijing of China. Stud. Biol. 2010, 4, 71–82. [Google Scholar] [CrossRef]
- Jokimäki, J.; Suhonen, J. Distribution and Habitat Selection of Wintering Birds in Urban Environments. Landsc. Urban Plan. 1998, 39, 253–263. [Google Scholar] [CrossRef]
- Jerzak, L. Synurbizacja sroki Pica pica w Eurazji; Uniwersytet Zielonogórski: Zielona Góra, Poland, 2002. [Google Scholar]
- Czechowski, P.; Bocheński, M.; Ciebiera, O. Decline Of Jackdaws Corvus monedula In The City Of Zielona Góra. Int. Stud. Sparrows 2013, 37, 32–36. [Google Scholar] [CrossRef] [Green Version]
- Salvati, L. Nest Site and Breeding Habitat Characteristics in Urban Jackdaws Corvus monedula in Rome (Italy). Acta Ornithol. 2002, 37, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Meyrier, E.; Jenni, L.; Bötsch, Y.; Strebel, S.; Erne, B.; Tablado, Z. Happy to Breed in the City? Urban Food Resources Limit Reproductive Output in Western Jackdaws. Ecol. Evol. 2017, 7, 1363–1374. [Google Scholar] [CrossRef]
- Vuorisalo, T.; Andersson, H.; Hugg, T.; Lahtinen, R.; Laaksonen, H.; Lehikoinen, E. Urban Development from an Avian Perspective: Causes of Hooded Crow (Corvus corone cornix) Urbanisation in Two Finnish Cities. Landsc. Urban Plan. 2003, 62, 69–87. [Google Scholar] [CrossRef]
- The European Red List of Birds. Luxembourg: Publications Office of the European Union. Birdlife Int. 2021, 28, 3–19. [Google Scholar]
- Jadczyk, P.; Drzeniecka-Osiadacz, A. Feeding Strategy of Wintering Rooks Corvus frugilegus L. in Urban Habitats. Pol. J. Ecol. 2013, 61, 587–596. [Google Scholar]
- Obukhova, N.Y. Nesting Dynamics of Corvids (Corvidae) in the City of Moscow and Moscow Oblast. Biol. Bull. 2018, 45, 1096–1105. [Google Scholar] [CrossRef]
- Zimaroyeva, A.A.; Matsyura, A.V.; Jankowski, K. Spatial Patterns of Habitat Distribution of Corvidae (the Case of Urban-Rural Gradient). Biosyst. Divers. 2016, 24, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Díaz, M. Food Choice by Seed-Eating Birds in Relation to Seed Chemistry. Comp. Biochem. Physiol. Part A Physiol. 1996, 113, 239–246. [Google Scholar] [CrossRef]
- Andersson, M.; Wallander, J.; Isaksson, D. Predator Perches: A Visual Search Perspective. Funct. Ecol. 2009, 23, 373–379. [Google Scholar] [CrossRef]
- Fernández-Juricic, E.; Blumstein, D.T.; Abrica, G.; Manriquez, L.; Adams, L.B.; Adams, R.; Daneshrad, M.; Rodriguez-Prieto, I. Relationships of Anti-Predator Escape and Post-Escape Responses with Body Mass and Morphology: A Comparative Avian Study. Evol. Ecol. Res. 2006, 8, 731–752. [Google Scholar]
- Tzortzakaki, O.; Kati, V.; Kassara, C.; Tietze, D.T.; Giokas, S. Seasonal Patterns of Urban Bird Diversity in a Mediterranean Coastal City: The Positive Role of Open Green Spaces. Urban Ecosyst. 2018, 21, 27–39. [Google Scholar] [CrossRef]
- Mason, C.F.; Macdonald, S.M. Distribution of Foraging Rooks, Corvus frugilegus, and Rookeries in a Landscape in Eastern England Dominated by Winter Cereals. Folia Zool. 2004, 53, 179–188. [Google Scholar]
- Waite, R.K. Winter Habitat Selection and Foraging Behaviour in Sympatric Corvids. Ornis Scand. 1984, 15, 55–62. [Google Scholar] [CrossRef]
- Zmihorski, M.; Halba, R.; Mazgajski, T.D. Long-Term Spatio-Temporal Dynamics of Corvids Wintering in Urban Parks of Warsaw, Poland. Ornis Fenn. 2010, 87, 61–68. [Google Scholar]
- Pons, J.; Pausas, J.G. Modelling Jay (Garrulus Glandarius) Abundance and Distribution for Oak Regeneration Assessment in Mediterranean Landscapes. For. Ecol. Manag. 2008, 256, 578–584. [Google Scholar] [CrossRef]
- Mazgajski, T.D.; Zmihorski, M.; Halba, R.; Woźniak, A. Long-Term Population Trends of Corvids Wintering in Urban Parks in Central Poland. Pol. J. Ecol. 2008, 56, 521–526. [Google Scholar]
- Sorace, A.; Gustin, M. Distribution of Generalist and Specialist Predators along Urban Gradients. Landsc. Urban Plan. 2009, 90, 111–118. [Google Scholar] [CrossRef]
- Bossema, I. Jays and Oaks: An Eco-Ethological Study of a Symbiosis. Behaviour 1979, 70, 1–116. [Google Scholar] [CrossRef] [Green Version]
- Morán-López, T.; Alonso, C.L.; Díaz, M. Landscape Effects on Jay Foraging Behavior Decrease Acorn Dispersal Services in Dehesas. Acta Oecologica 2015, 69, 52–64. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.D.; Cleeton, S.H.; Lyons, T.P.; Miller, J.R. Urbanization and the Predation Paradox: The Role of Trophic Dynamics in Structuring Vertebrate Communities. Bioscience 2012, 62, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Evans, K.L.; Hatchwell, B.J.; Parnell, M.; Gaston, K.J. A Conceptual Framework for the Colonisation of Urban Areas: The Blackbird Turdus Merula as a Case Study. Biol. Rev. 2010, 85, 643–667. [Google Scholar] [CrossRef]
- Tobias, J.A.; Ottenburghs, J.; Pigot, A.L. Avian Diversity: Speciation, Macroevolution, and Ecological Function. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 533–560. [Google Scholar] [CrossRef]
- Moi, D.A.; García-Ríos, R.; Hong, Z.; Daquila, B.V.; Mormul, R.P. Intermediate Disturbance Hypothesis in Ecology: A Literature Review. Ann. Zool. Fenn. 2020, 57, 67–78. [Google Scholar] [CrossRef]
- Kulemeyer, C.; Asbahr, K.; Gunz, P.; Frahnert, S.; Bairlein, F. Functional Morphology and Integration of Corvid Skulls—A 3D Geometric Morphometric Approach. Front. Zool. 2009, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Rolando, A. Data on Eco-Ethology of Coexistence in Corvids in North-Western Italy. Bolletino Zool. 1988, 55, 315–321. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Variables Correlated | RM | p-Value |
|---|---|---|
| Carrion/Hooded Crow × Rook | 0.100 | <0.01 |
| Carrion/Hooded Crow × Western Jackdaw | −0.010 | >0.05 |
| Carrion/Hooded Crow × Eurasian Jay | 0.110 | <0.01 |
| Carrion/Hooded Crow × Eurasian Magpie | 0.150 | <0.001 |
| Rook × Western Jackdaw | 0.035 | >0.05 |
| Rook × Eurasian Jay | −0.016 | >0.05 |
| Rook × Eurasian Magpie | 0.084 | <0.01 |
| Western Jackdaw × Eurasian Jay | −0.002 | >0.05 |
| Western Jackdaw × Eurasian Magpie | 0.010 | >0.05 |
| Eurasian Jay × Eurasian Magpie | 0.080 | <0.01 |
| Variable | Estimate | SE | z-Value | p-Value |
|---|---|---|---|---|
| Carrion/Hooded Crow | ||||
| Intercept | −3.050 | 0.737 | 4.141 | p < 0.001 |
| Bare soil | 0.022 | 0.007 | 2.977 | 0.003 |
| Building floors | 0.068 | 0.048 | 1.421 | 0.155 |
| Bush | 0.002 | 0.008 | 0.192 | 0.848 |
| Cats | −0.019 | 0.083 | 0.224 | 0.823 |
| Dogs | 0.118 | 0.074 | 1.595 | 0.111 |
| Grass | 0.016 | 0.005 | 3.200 | 0.001 |
| Light | −0.000 | 0.004 | 0.058 | 0.954 |
| Noise | −0.007 | 0.011 | 0.587 | 0.557 |
| Pedestrians | −0.008 | 0.004 | 1.756 | 0.079 |
| Tree | −0.000 | 0.006 | 0.069 | 0.945 |
| Rook | ||||
| Intercept | −5.551 | 1.952 | 2.843 | 0.004 |
| Bare soil | 0.019 | 0.017 | 1.115 | 0.265 |
| Building floors | −0.173 | 0.139 | 1.246 | 0.213 |
| Bush | −0.044 | 0.026 | 1.706 | 0.088 |
| Cats | −0.012 | 0.292 | 0.041 | 0.968 |
| Dogs | −0.390 | 0.291 | 1.343 | 0.179 |
| Grass | 0.034 | 0.011 | 2.947 | 0.003 |
| Light | −0.016 | 0.016 | 0.990 | 0.322 |
| Noise | 0.004 | 0.031 | 0.139 | 0.890 |
| Pedestrians | −0.010 | 0.019 | 0.512 | 0.609 |
| Tree | −0.034 | 0.018 | 1.923 | 0.055 |
| Western Jackdaw | ||||
| Intercept | −1.605 | 0.7629 | 2.104 | 0.035 |
| Bare soil | −0.016 | 0.0070 | 2.350 | 0.019 |
| Building floors | 0.053 | 0.038 | 1.374 | 0.170 |
| Bush | −0.008 | 0.008 | 0.922 | 0.357 |
| Cats | 0.043 | 0.086 | 0.498 | 0.619 |
| Dogs | −0.140 | 0.084 | 1.665 | 0.096 |
| Grass | −0.009 | 0.005 | 1.736 | 0.083 |
| Light | 0.004 | 0.003 | 1.304 | 0.192 |
| Noise | −0.009 | 0.011 | 0.800 | 0.424 |
| Pedestrians | 0.000 | 0.002 | 0.198 | 0.843 |
| Tree | −0.010 | 0.006 | 1.557 | 0.120 |
| Eurasian Jay | ||||
| Intercept | −4.795 | 1.551 | 3.092 | 0.002 |
| Bare soil | 0.005 | 0.012 | 0.419 | 0.675 |
| Building floors | −0.238 | 0.135 | 1.766 | 0.077 |
| Bush | 0.017 | 0.013 | 1.331 | 0.183 |
| Cats | 0.131 | 0.105 | 1.250 | 0.211 |
| Dogs | 0.089 | 0.112 | 0.796 | 0.426 |
| Grass | 0.010 | 0.010 | 1.041 | 0.298 |
| Light | −0.012 | 0.009 | 1.322 | 0.186 |
| Noise | 0.026 | 0.021 | 1.237 | 0.216 |
| Pedestrians | −0.027 | 0.015 | 1.767 | 0.077 |
| Tree | 0.019 | 0.009 | 2.186 | 0.029 |
| Eurasian Magpie | ||||
| Intercept | −2.914 | 0.695 | 4.191 | <0.001 |
| Bare soil | 0.015 | 0.006 | 2.776 | 0.006 |
| Building floors | 0.021 | 0.030 | 0.709 | 0.478 |
| Bush | 0.002 | 0.007 | 0.245 | 0.806 |
| Cats | 0.004 | 0.075 | 0.053 | 0.957 |
| Dogs | −0.025 | 0.062 | 0.401 | 0.689 |
| Grass | 0.027 | 0.004 | 6.644 | <0.001 |
| Light | −0.003 | 0.003 | 1.151 | 0.250 |
| Noise | 0.021 | 0.009 | 2.359 | 0.018 |
| Pedestrians | −0.012 | 0.004 | 3.385 | <0.001 |
| Tree | −0.003 | 0.005 | 0.645 | 0.519 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou Zeid, F.; Morelli, F.; Ibáñez-Álamo, J.D.; Díaz, M.; Reif, J.; Jokimäki, J.; Suhonen, J.; Kaisanlahti-Jokimäki, M.-L.; Markó, G.; Bussière, R.; et al. Spatial Overlap and Habitat Selection of Corvid Species in European Cities. Animals 2023, 13, 1192. https://doi.org/10.3390/ani13071192
Abou Zeid F, Morelli F, Ibáñez-Álamo JD, Díaz M, Reif J, Jokimäki J, Suhonen J, Kaisanlahti-Jokimäki M-L, Markó G, Bussière R, et al. Spatial Overlap and Habitat Selection of Corvid Species in European Cities. Animals. 2023; 13(7):1192. https://doi.org/10.3390/ani13071192
Chicago/Turabian StyleAbou Zeid, Farah, Federico Morelli, Juan Diego Ibáñez-Álamo, Mario Díaz, Jiří Reif, Jukka Jokimäki, Jukka Suhonen, Marja-Liisa Kaisanlahti-Jokimäki, Gábor Markó, Raphaël Bussière, and et al. 2023. "Spatial Overlap and Habitat Selection of Corvid Species in European Cities" Animals 13, no. 7: 1192. https://doi.org/10.3390/ani13071192