
Habituation to Livestock Trailer and Its Influence on Stress Responses during Transportation in Goats
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Behavioral Observations
2.3. Blood Sampling
2.4. Cortisol
2.5. Creatine Kinase
2.6. Differential Leukocyte Counts
2.7. Catecholamines and Other Biogenic Amines
2.8. Statistical Analysis
3. Results
3.1. Behavior
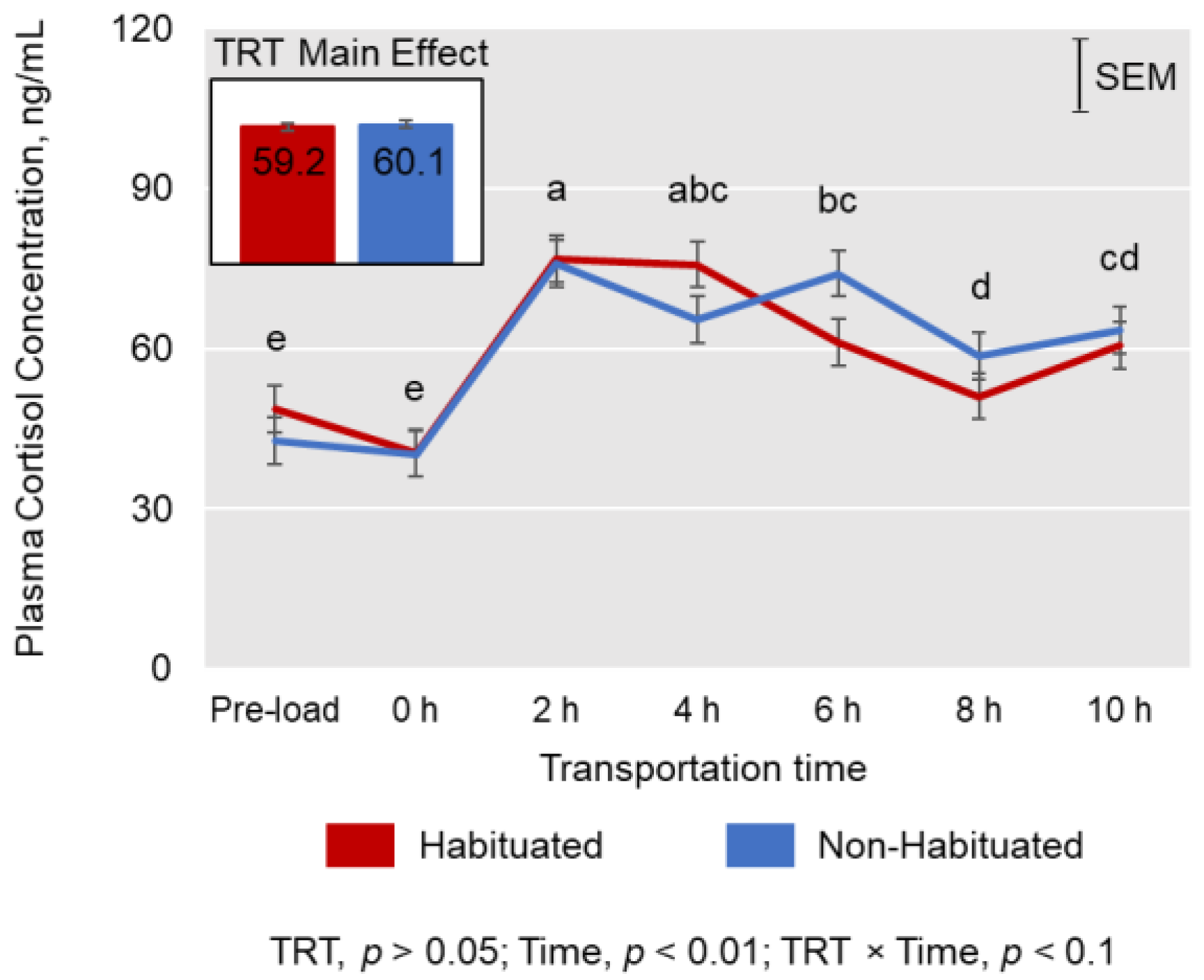
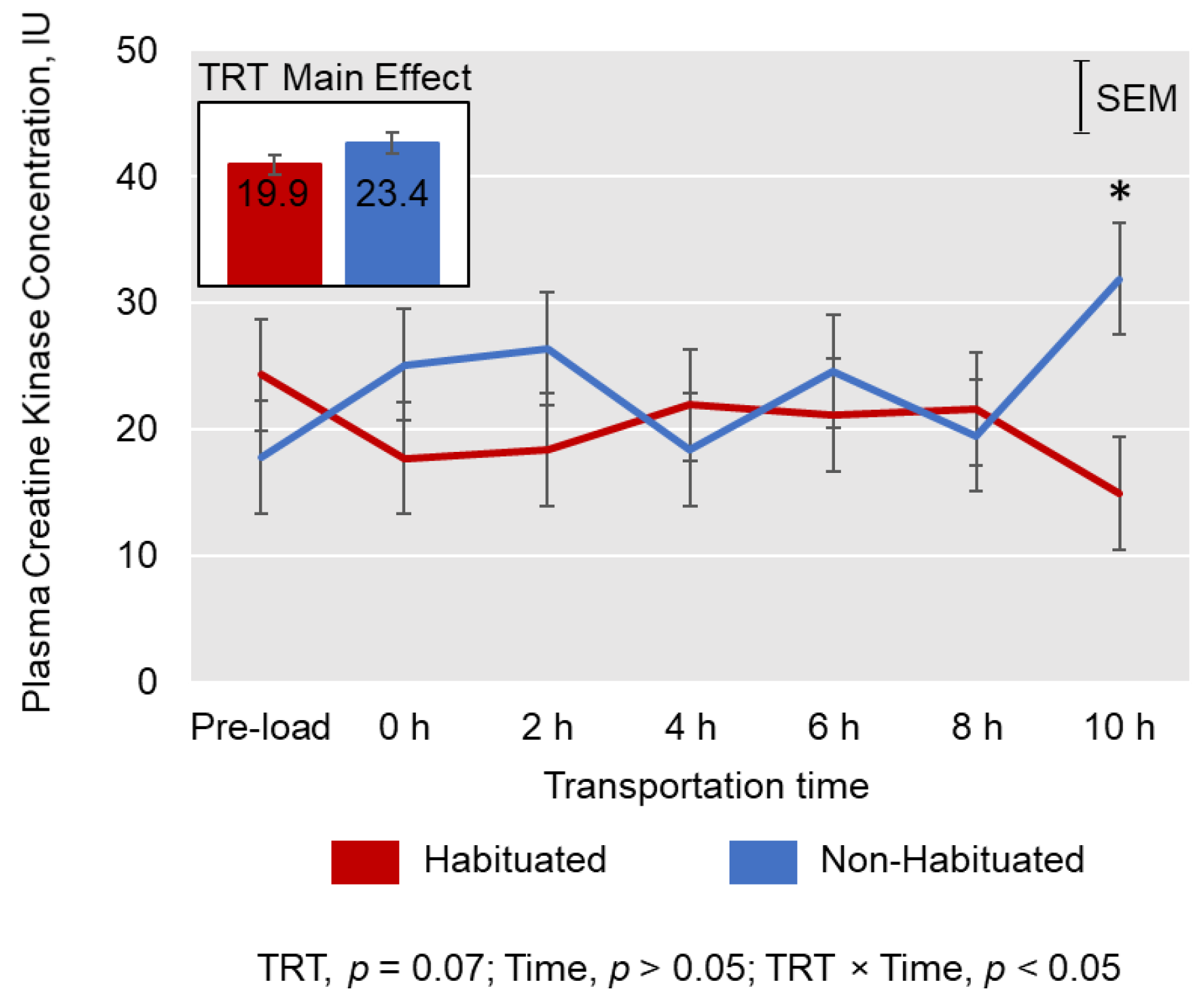
3.2. Cortisol, Creatine Kinase, and Leukocyte Counts
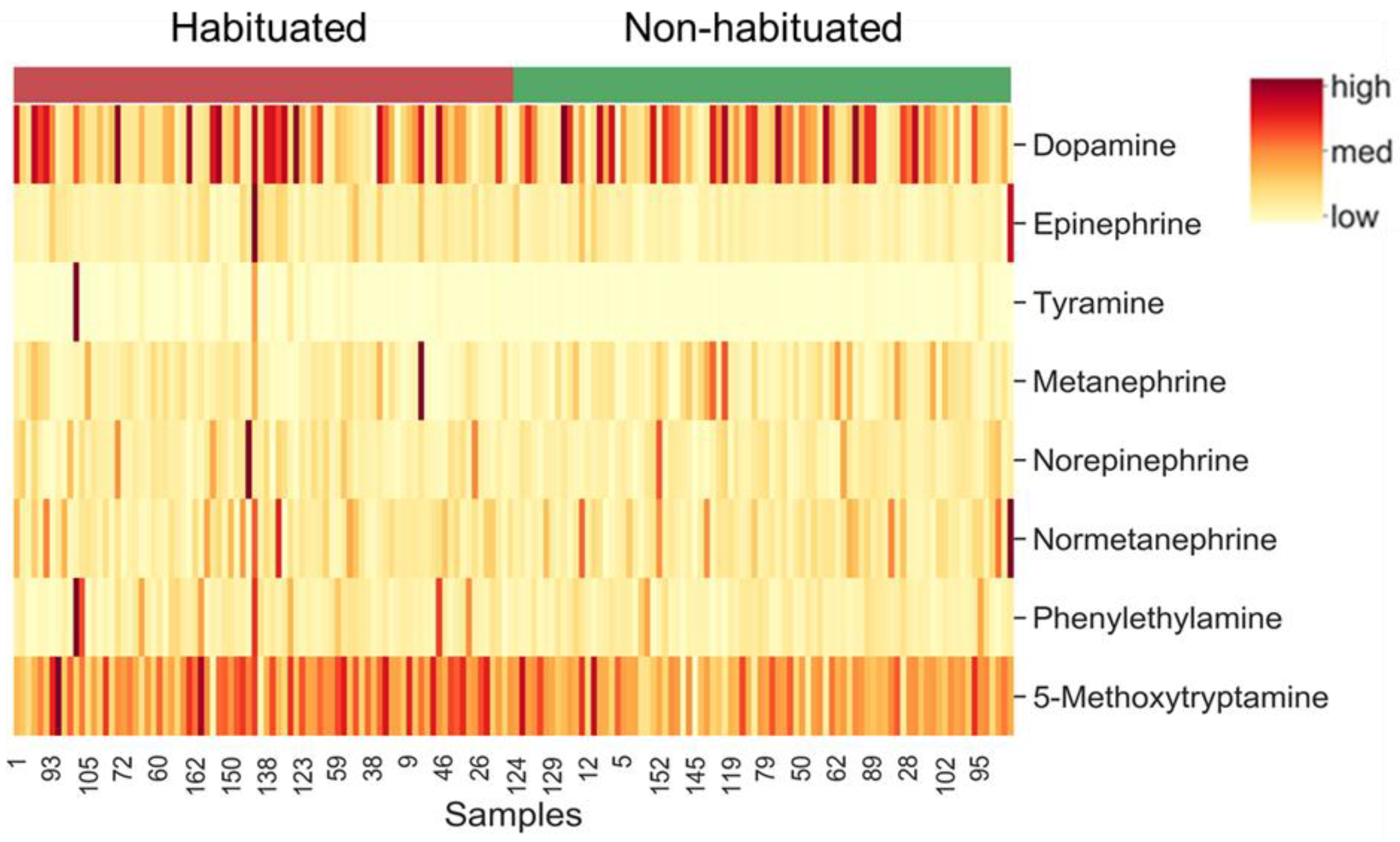
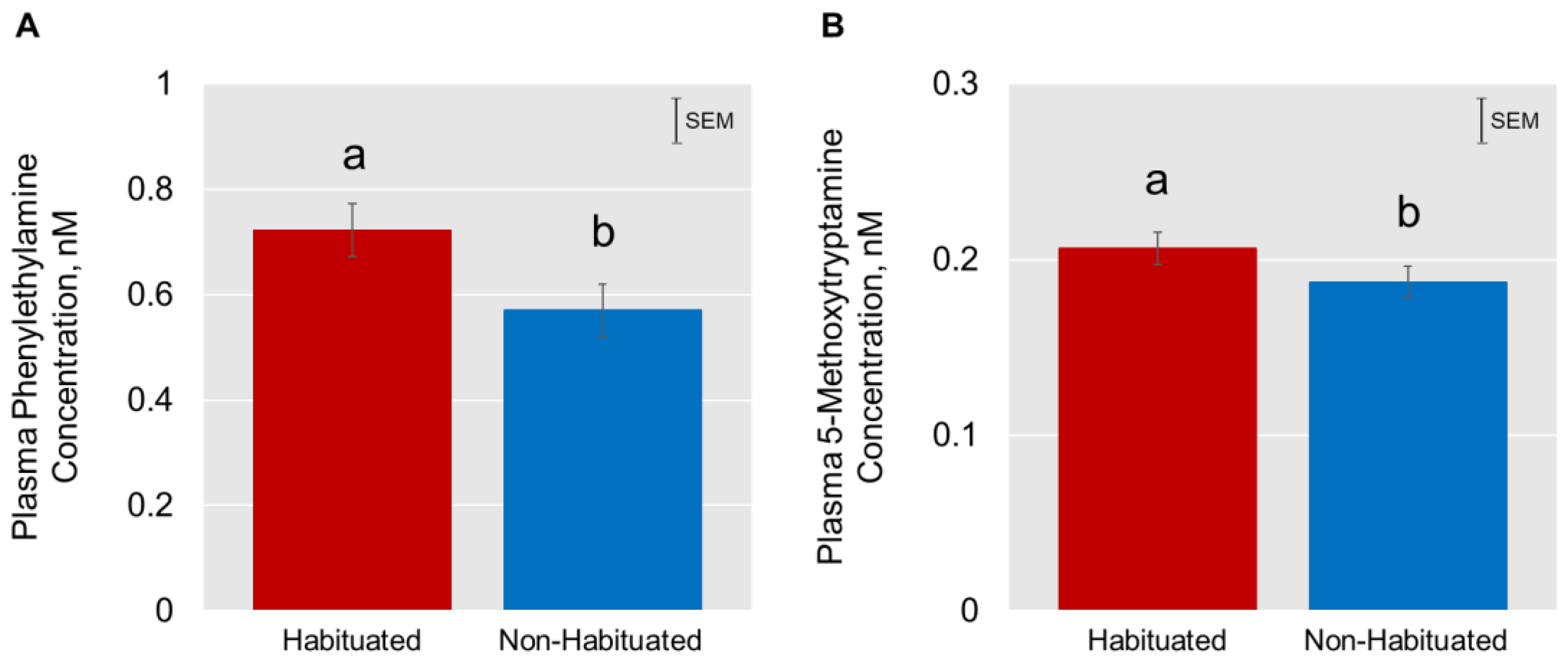
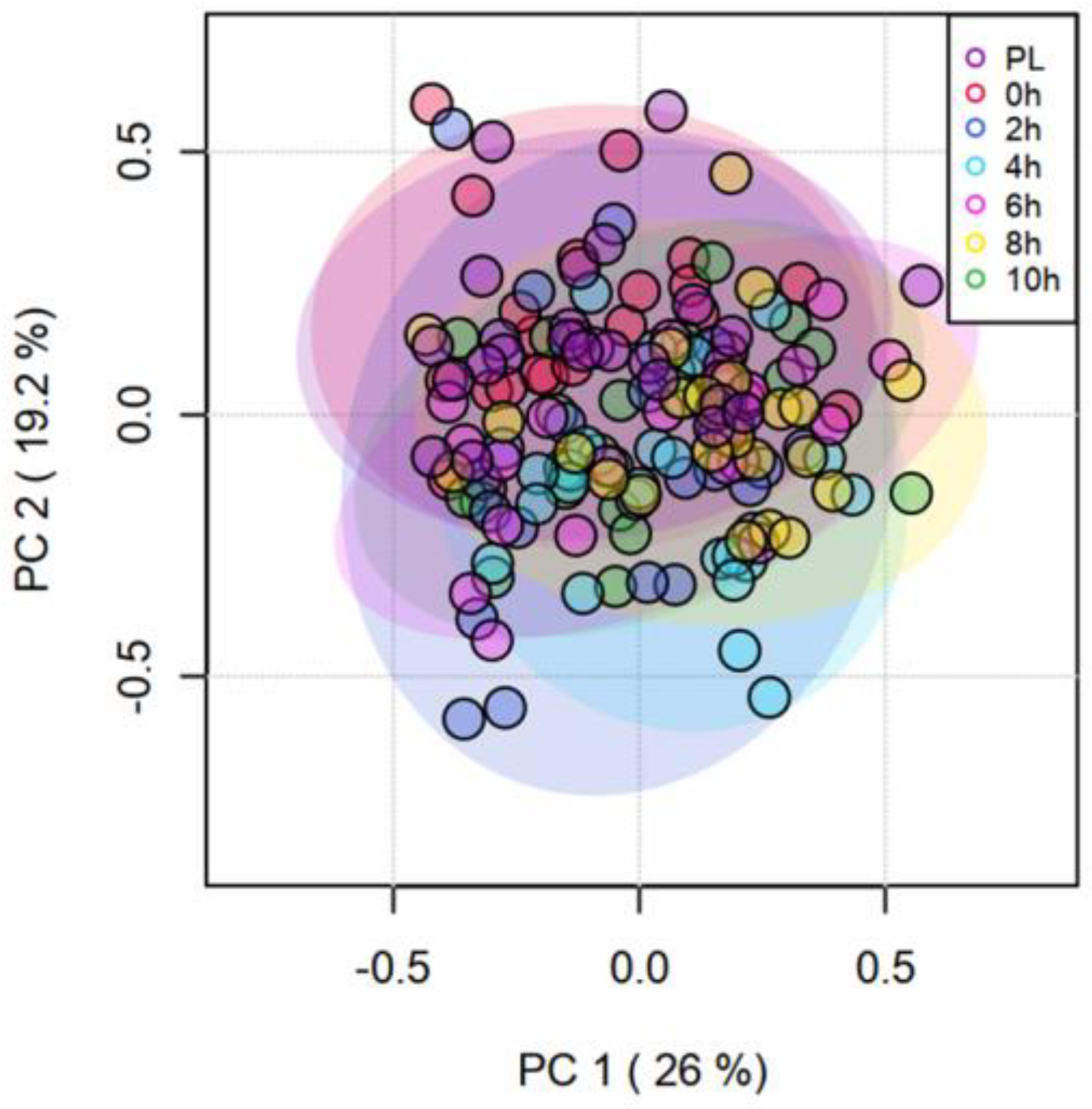
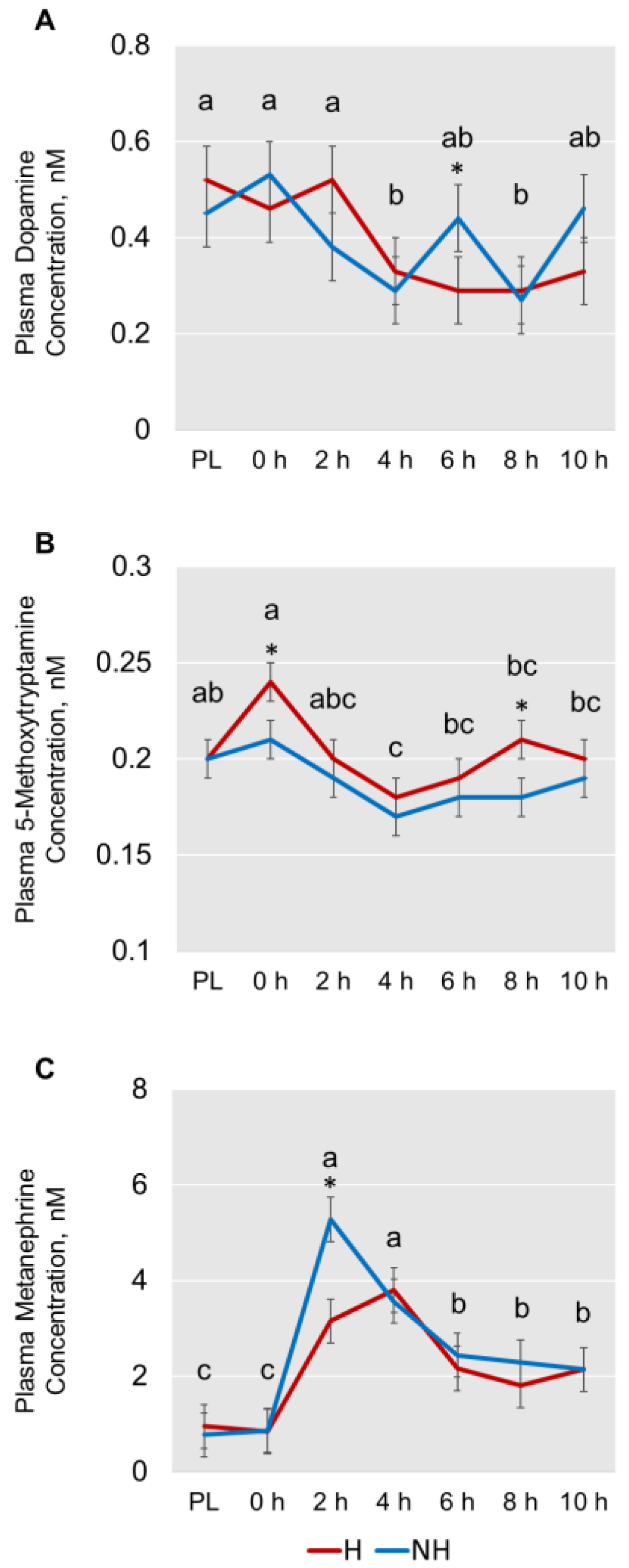
3.3. Catecholamines and Other Biogenic Amines
4. Discussion
4.1. Behavior
4.2. Cortisol, Creatine Kinase, and Leukocyte Counts
4.3. Catecholamines and Other Biogenic Amines
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kannan, G.; Terrill, T.H.; Kouakou, B.; Gazal, O.S.; Gelaye, S.; Amoah, E.A.; Samake, S. Transportation of goats: Effects on physiological stress responses and live weight loss. J. Anim. Sci. 2000, 78, 1450–1457. [Google Scholar] [CrossRef] [PubMed]
- Kadim, I.T.; Mahgoub, O.; AlKindi, A.Y.; Al-Marzooqi, W.; Al-Saqri, N.M.; Almaney, M.; Mahmoud, I.Y. Effect of transportation at high ambient temperatures on physiological responses, carcass, and meat quality characteristics in two age groups of Omani sheep. Asian-Australas. J. Anim. Sci. 2007, 20, 424–431. [Google Scholar] [CrossRef]
- Batchu, P.; Hazard, T.; Lee, J.H.; Terrill, T.H.; Kouakou, B.; Kannan, G. High-condensed tannin diet and transportation stress in goats: Effects on physiological responses, gut microbial counts and meat quality. Animals 2021, 11, 2857. [Google Scholar] [CrossRef] [PubMed]
- Kvetnansky, R.; Sabban, E.L.; Palkovits, M. Catecholaminergic systems in stress: Structural and molecular genetic approaches. Physiol. Rev. 2009, 89, 535–606. [Google Scholar] [CrossRef]
- Goldstein, D.S. Catecholamines and stress. Endocr. Regul. 2003, 37, 69–80. [Google Scholar]
- Sabelli, H. Phenylethylamine deficit and replacement in depressive illness. In Natural Medications of Psychiatric Disorders; Mischoulon, D., Rosenbaum, J.F., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2002; pp. 83–99. [Google Scholar]
- Hara, H.; Iigo, M.; Ohtani-Kaneko, R.; Nakamura, N.; Suzukin, T.; Reiter, R.J.; Hirata, K. Administration of melatonin and related indoles prevents exercise-induced cellular oxidative changes in rats. Neurosignals 1997, 6, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Wechsler, B.; Lea, S.E.G. Adaptation by learning: Its significance for farm animal husbandry. Appl. Anim. Behav. Sci. 2007, 108, 197–214. [Google Scholar] [CrossRef]
- Abramson, C.I.; Kieson, E. Conditioning methods for animals in agriculture: A review. Cienc. Anim. Bras. 2016, 17, 359–375. [Google Scholar] [CrossRef] [Green Version]
- Thompson, R.F.; Spencer, W.A. Habituation: A model phenomenon for the study of neuronal substrates of behavior. Psychol. Rev. 1966, 73, 16–43. [Google Scholar] [CrossRef] [Green Version]
- Ujita, A.; Seekford, Z.; Kott, M.; Goncherenko, G.; Dias, N.W.; Feuerbacher, E.; Bergamasco, L.; Jacobs, L.; Eversole, D.E.; Negrão, J.A.; et al. Habituation protocols improve behavioral and physiological responses of beef cattle exposed to students in an animal handling class. Animals 2021, 11, 2159. [Google Scholar] [CrossRef]
- Kannan, G.; Mench, J.A. Prior handling does not significantly reduce the stress response to preslaughter handling in broiler chickens. Appl. Anim. Behav. Sci. 1997, 51, 87–99. [Google Scholar] [CrossRef]
- Dai, F.; Mazzola, S.; Cannas, S.; Heinzl, E.U.L.; Padalino, B.; Minero, M.; Costa, E.D. Habituation to transport helps reducing stress-related behavior in donkeys during loading. Front. Vet. Sci. 2020, 7, 593138. [Google Scholar] [CrossRef] [PubMed]
- Ag Guide. ADSA-ASAS-PSA Guide for the Care and Use of Agricultural Animals in Research and Teaching, 4th ed.; ADSA, ASAS, PSA: Champaign, IL, USA, 2020; Available online: https://www.asas.org/services/ag-guide (accessed on 12 October 2020).
- Zheng, J.; Mandal, R.; Wishart, D.S. A sensitive, high-throughput LC-MS/MS method for measuring catecholamines in low volume serum. Anal. Chim. Acta 2018, 1037, 159–167. [Google Scholar] [CrossRef]
- Rusilowicz, M.; Dickinson, M.; Charlton, A.; O’Keefe, S.; Wilson, J. A batch correction method for liquid chromatography–mass spectrometry data that does not depend on quality control samples. Metabolomics 2016, 12, 56. [Google Scholar] [CrossRef] [Green Version]
- Wickham, S.L.; Collins, T.; Barnes, A.L.; Miller, D.W.; Beatty, D.T.; Stckman, D.; Blache, F.; Wemelsfelder, F.; Fleming, P.A. Qualitative behavioral assessment of transport-naïve and transport-habituated sheep. J. Anim. Sci. 2012, 90, 4523–4535. [Google Scholar] [CrossRef] [Green Version]
- Houpt, K.A.; Wickens, C.I. Handling and transport of horses. In Livestock Handling and Transport; Grandin, T., Ed.; CABI: Boston, MA, USA, 2014; pp. 347–369. [Google Scholar]
- Naldurtiker, A.; Batchu, P.; Kouakou, B.; Terrill, T.H.; McCommon, G.W.; Kannan, G. Differential gene expression analysis us-ing RNA-seq in the blood of goats exposed to transportation stress. Sci. Rep. 2023, 13, 1984. [Google Scholar] [CrossRef] [PubMed]
- Kannan, G.; Terrill, T.H.; Kouakou, B.; Gelaye, S.; Amoah, E.A. Simulated preslaughter holding and isolation effects on stress responses and live weight shrinkage in meat goats. J. Anim. Sci. 2002, 80, 1771–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannan, G.; Estrada-Reyes, Z.M.; Batchu, P.; Kouakou, B.; Terrill, T.H.; Naldurtiker, A. Social isolation of goats: Significance of visual contact with conspecifics on behavioral and physiological responses. J. Anim. Sci. 2021, 99, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kannan, G.; Batchu, P.; Naldurtiker, A.; Dykes, G.S.; Kouakou, B.; Terrill, T.H.; Gurrapu, P. Influence of excitability rate on physiological responses to stress in goats. Animals 2022, 12, 1023. [Google Scholar] [CrossRef]
- Minka, N.S.; Ayo, J.O. Physiological responses of food animals to road transportation stress. Afr. J. Biotechnol. 2009, 8, 7415–7427. [Google Scholar]
- Broom, D.M. Cortisol: Often not the best indicator of stress and poor welfare. Physiol. News 2017, 107, 30–32. [Google Scholar] [CrossRef]
- Zulkifli, I.; Norbaiyah, B.; Cheah, Y.W.; Soleimani, A.F.; Sazili, A.Q.; Rajion, M.A.; Goh, Y.M. Physiological responses in goats subjected to road transportation under the hot, humid tropical conditions. Int. J. Agric. Biol. 2010, 12, 840–844. [Google Scholar]
- Ishizaki, H.; Kariya, Y. Road transportation stress promptly increases bovine peripheral blood absolute NL cell counts and cortisol levels. J. Vet. Med. Sci. 2010, 72, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Elenkov, I.J.; Chrousos, G.P. Stress hormones, Th1/Th2 patterns, pro/anti-inflammatory cytokines and susceptibility to disease. Trends Endocrinol. Metab. 1999, 10, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Baird, M.F.; Graham, S.M.; Baker, J.S.; Bickerstaff, G.E. Creatine-kinase- and exercise-related muscle damage implications for muscle performance and recovery. J. Nutr. Metab. 2012, 12, 960363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heled, Y.; Bloom, M.S.; Wu, T.J.; Stephens, Q.; Deuster, P.A. CK-MM and ACE genotypes and physiological prediction of the creatine kinase response to exercise. J. Appl. Physiol. 2007, 103, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Earley, B.; Murray, M.; Prendiville, D.J.; Pintado, B.; Borque, C.; Canali, E. The effects of transport by road and sea on physiology, immunity and behaviour of beef cattle. Res. Vet. Sci. 2012, 92, 531–541. [Google Scholar] [CrossRef]
- Chulayo, A.Y.; Muchenje, V. The effects of pre-slaughter stress and season on the activity of plasma creatine kinase and mutton quality from different sheep breeds slaughtered at a smallholder abattoir. Asian Australas. J. Anim. Sci. 2013, 26, 1762–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teke, B.; Akdag, F.; Ekiz, B.; Ugurlu, M. Effects of different lairage times after long distance transportation on carcass and meat quality characteristics of Hungarian Simmental bulls. Meat Sci. 2014, 96, 24–229. [Google Scholar] [CrossRef]
- Evans, G.F.F.; Haller, R.G.; Wyrick, P.S.; Parkey, R.W.; Fleckenstein, J.L. Submaximal delayed-onset muscle soreness: Correlations between MR imaging findings and clinical measures. Radiology 1998, 208, 815–820. [Google Scholar] [CrossRef]
- Baik, J. Stress and dopaminergic reward system. Exp. Mol. Med. 2020, 52, 1879–1890. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, D.S.; Holmes, C. Neuronal source of plasma dopamine. Clin. Chem. 2008, 54, 1864–1871. [Google Scholar] [CrossRef] [Green Version]
- Cabib, S.; Puglisi-Allegra, S. The mesoaccumbens dopamine in coping with stress. Neurosci. Biobehav. Rev. 2012, 36, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Yaworsky, D.; Wu, A.; Hill, D.W. The use of plasma metanephrine to normetanephrine ratio to determine epinephrine poisoning. Clin. Chim. Acta 2005, 353, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.D. Mammalian central nervous system trace amines. Pharmacologic amphetamines, physiologic neuromodulators. J. Neurochem. 2004, 90, 257–271. [Google Scholar] [CrossRef]
- Pastore, P.; Favaro, G.; Badocco, D.; Tapparo, A.; Cavelli, S.; Saccini, G. Determination of biogenic amines in chocolate nby ion chromatographic separation and pulsed integrated ampometric detection with implemented wave-form at Au disposable electrode. J. Chromatogr. 2005, 1098, 111–115. [Google Scholar] [CrossRef]
- Landete, J.M.; Ferrer, S.; Polo, L.; Pardo, I. Biogenic amines in wines from three Spanish regions. J. Agric. Food Chem. 2005, 53, 1119–1124. [Google Scholar] [CrossRef]
- Lapin, I.P. Antagonism by CPP (+/−)-3-(2-carboxypiperazin-4-yl)-propyl-1-phosphonic acid, of beta-phenylethylamine (PEA)-induced hypermotility in mice of different strains. Pharmacol. Biochem. Behav. 1996, 55, 175–178. [Google Scholar] [CrossRef]
- Sotnikova, T.D.; Budygin, E.A.; Jones, S.R.; Dykstra, L.A.; Caron, M.G.; Gainetdinov, R.R. Dopamine transporter-dependent and –independent actions of trace amine beta-phenylethylamine. J. Neurochem. 2004, 91, 362–373. [Google Scholar] [CrossRef]
- Szabo, A.; Billett, E.; Turner, J. Phenylethylamine, a possible link to the antidepressant effects of exercise? Br. J. Sports Med. 2001, 35, 342–343. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-J.; Kim, H.R.; Lee, C.Y.; Hyun, S.-A.; Ko, M.Y.; Lee, B.-S.; Hwang, D.Y.; Ka, M. 2-Phenylethylamine (PEA) ameliorates corticosterone-induced depression-like phenotype via the BDNF/TrkB/CREB signaling pathway. Int. J. Mol. Sci. 2020, 21, 9103. [Google Scholar] [CrossRef] [PubMed]
- Pevet, P. Is 5-methoxytryptamine a pineal hormone? Psychoneuroendocrinology 1983, 8, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Semak, I.; Pisarchik, A.; Sweatman, T.; Szczesniewski, A.; Wortsman, J. Conversion of L-tryptophan to serotonin and melatonin in human melanoma cells. FEBS Lett. 2002, 511, 102–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, O.; Jonsson, G. In vivo formation of 5-methoxytryptamine from melatonin in rat. J. Neurochem. 1981, 36, 2013–2018. [Google Scholar] [CrossRef]
- Cannizzaro, C.; Plescia, F.; Gagliano, M.; Cannizzaro, G.; Mantia, G.; La Barbera, M.; Provenzano, G.; Cannizzaro, E. Perinatal exposure to 5-metoxytryptamine, behavioural-stress reactivity and functional response of 5-HT1A receptors in the adolescent rat. Behav. Brain Res. 2008, 186, 98–106. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior | Time 1 | Treatment (TRT) | n 6 | Time Effect 7 | TRT Effect 8 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st 5-min | 2nd 5-min | 3rd 5-min | 4th 5-min | H 4 | NH 5 | ||||||||||
| Median 2 | IQR 3 | Median | IQR | Median | IQR | Median | IQR | Median | IQR | Median | IQR | ||||
| Standing | 15 | 9 | 14 | 8 | 17 | 6 | 13 | 10 | 15 | 6.5 | 14.5 | 11.5 | 84 | p < 0.01 | NS 9 |
| Moving | 5 | 6 | 5 | 6 | 3 | 4 | 4.5 | 6 | 5 | 5.5 | 4 | 8.5 | 84 | p < 0.01 | NS |
| Agonistic | 1 | 3 | 1 | 3 | 0 | 2 | 1 | 3.5 | 1 | 0 | 1 | 3 | 84 | NS | NS |
| Climbing | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.5 | 84 | NS | NS |
| Time | Neutrophil | Lymphocyte | N/L Ratio | ||||||
|---|---|---|---|---|---|---|---|---|---|
| TRT | Time | TRT | Time | TRT | Time | ||||
| H 1 | NH 2 | H | NH | H | NH | ||||
| Pre-Load | 66.6 | 67.3 | 66.9 e | 32.3 | 36.5 | 34.4 a | 3.1 | 2.7 | 2.9 d |
| 0 h | 72.8 | 74.7 | 73.7 cde | 25.9 | 24.1 | 25.0 b | 3.2 | 3.5 | 3.3 d |
| 2 h | 80.6 | 80.7 | 80.8 abc | 18.2 | 23.1 | 20.6 bc | 5.2 | 4.9 | 5.1 c |
| 4 h | 86.5 | 87.2 | 86.8 a | 12.7 | 12.1 | 12.4 c | 7.3 | 8.2 | 7.8 a |
| 6 h | 82.8 | 78.2 | 80.5 abc | 16.4 | 20.9 | 18.6 bc | 6.0 | 6.8 | 6.4 ab |
| 8 h | 81.3 | 86.3 | 83.8 abc | 17.7 | 12.9 | 15.3 c | 5.4 | 8.4 | 6.9 ab |
| 10 h | 82.5 | 72.8 | 77.7 cd | 16.6 | 35.9 | 26.2 ab | 5.9 | 4.9 | 5.4 bc |
| N 3 | 12 | 12 | 24 | 12 | 12 | 24 | 12 | 12 | 24 |
| SEM | 3.76 | 3.76 | 2.66 | 4.57 | 4.57 | 3.23 | 0.78 | 0.78 | 0.55 |
| TRT 4 | NS 5 | NS | NS | ||||||
| Time | p < 0.01 | p < 0.01 | p < 0.01 | ||||||
| TRT × Time | NS | NS | NS | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kannan, G.; Batchu, P.; Naldurtiker, A.; Dykes, G.S.; Gurrapu, P.; Kouakou, B.; Terrill, T.H.; McCommon, G.W. Habituation to Livestock Trailer and Its Influence on Stress Responses during Transportation in Goats. Animals 2023, 13, 1191. https://doi.org/10.3390/ani13071191
Kannan G, Batchu P, Naldurtiker A, Dykes GS, Gurrapu P, Kouakou B, Terrill TH, McCommon GW. Habituation to Livestock Trailer and Its Influence on Stress Responses during Transportation in Goats. Animals. 2023; 13(7):1191. https://doi.org/10.3390/ani13071191
Chicago/Turabian StyleKannan, Govind, Phaneendra Batchu, Aditya Naldurtiker, Gregory S. Dykes, Priyanka Gurrapu, Brou Kouakou, Thomas H. Terrill, and George W. McCommon. 2023. "Habituation to Livestock Trailer and Its Influence on Stress Responses during Transportation in Goats" Animals 13, no. 7: 1191. https://doi.org/10.3390/ani13071191