
Effect of a Saccharomyces cerevisiae Postbiotic Feed Additive on Salmonella Enteritidis Colonization of Cecal and Ovarian Tissues in Directly Challenged and Horizontally Exposed Layer Pullets

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
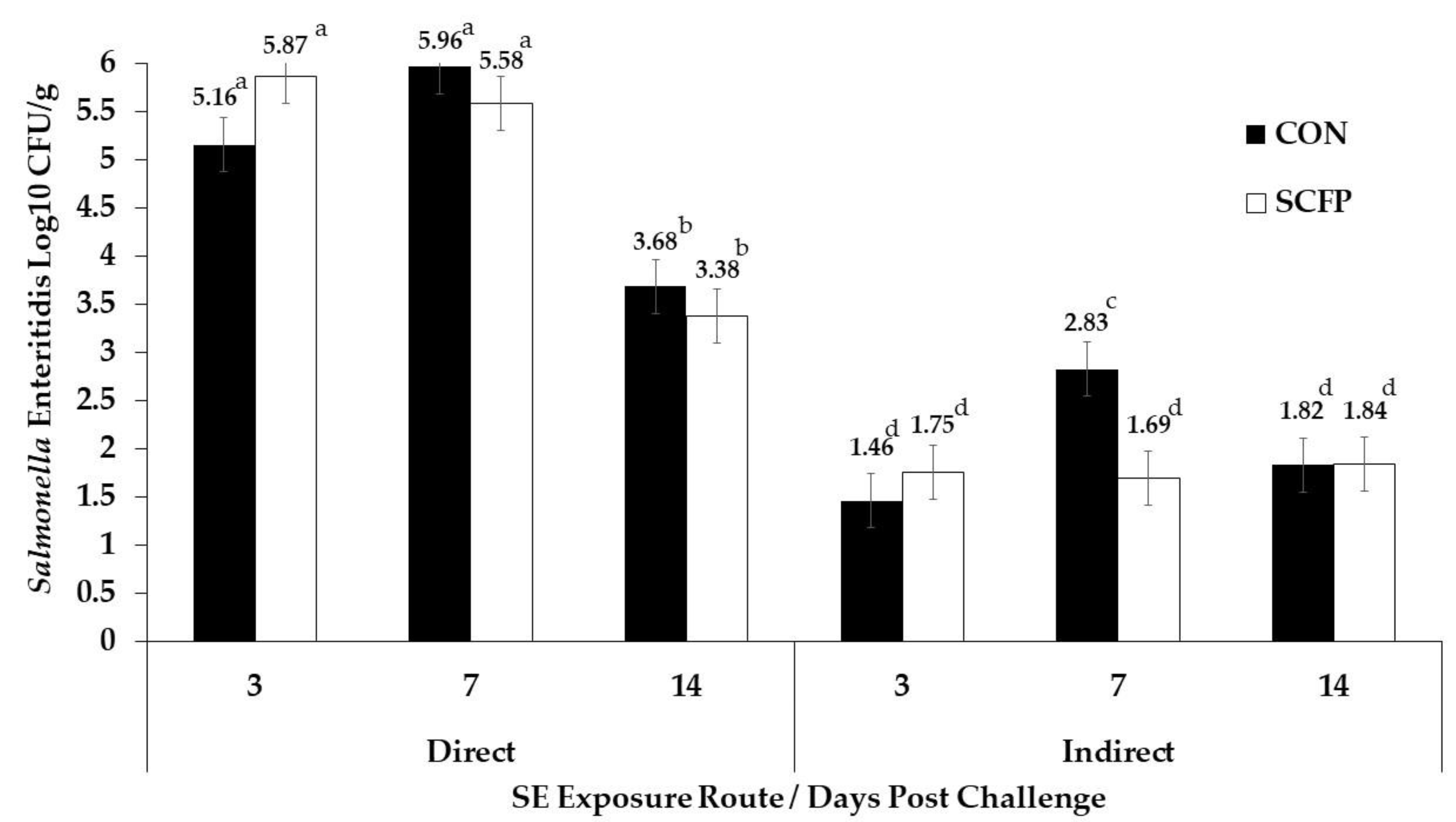
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, P.L.; Baker, A.R.; James, W.O. Strategies to control Salmonella and Campylobacter in raw poultry products. Rev. Sci. Et Tech. 1997, 16, 525–541. [Google Scholar] [CrossRef] [PubMed]
- Gast, R.K. Serotype-Specific and Serotype-Independent Strategies for Preharvest Control of Food-Borne Salmonella in Poultry. Avian Dis. 2007, 51, 817–828. [Google Scholar] [CrossRef]
- Chousalkar, K.; Gast, R.K.; Martelli, F.; Pande, V. Review of egg-related salmonellosis and reduction strategies in United States, Australia, United Kingdom and New Zealand. Crit. Rev. Microbiol. 2018, 44, 290–303. [Google Scholar] [CrossRef] [PubMed]
- O’Bryan, C.A.; Ricke, S.C.; Marcy, J.A. Public health Impact of Salmonella spp. on raw poultry: Current concepts and future prospects in the United States. Food Control 2022, 132, 108539. [Google Scholar] [CrossRef]
- Interagency Food Safety Analytics Collaboration. Foodborne Illness Source Attribution Estimates for 2017 for Salmonella, Escherichia coli O157, Listeria monocytogenes, and Campylobacter Using Multi-Year Outbreak Surveillance Data, United States. Available online: https://www.cdc.gov/foodsafety/ifsac/annual-reports.html (accessed on 20 January 2023).
- Agriculture, U.S.D.O.; Economic Research Service. Livestock, Dairy, and Poultry Outlook: November 2022. Available online: https://www.ers.usda.gov/webdocs/outlooks/105249/ldp-m-341.pdf?v=6452.1 (accessed on 20 January 2023).
- U.S. Food and Drug Administration. Prevention of Salmonella Enteritidis in Shell Eggs During Production, Storage, and Transportation; Final Rule. Fed. Regist. 2009, 74, 33030–33100. [Google Scholar]
- Trampel, D.W.; Holder, T.G.; Gast, R.K. Integrated Farm Management to Prevent Salmonella Enteritidis Contamination of Eggs. J. Appl. Poult. Res. 2014, 23, 353–365. [Google Scholar] [CrossRef]
- Al-Ajeeli, M.N.; Taylor, M.T.; Alvarado, C.Z.; Coufal, C.D. Comparison of Eggshell Surface Sanitization Technologies and Impacts on Consumer Acceptability. Poult. Sci. 2016, 95, 1191–1197. [Google Scholar] [CrossRef]
- Hofacre, C.L.; Rosales, A.G.; Da Costa, M.; Cookson, K.; Shaeffer, J.; Jones, M.K. Immunity and Protection Provided by Live Modified Vaccines Against Paratyphoid Salmonella in Poultry—An Applied Perspective. Avian Dis. 2021, 65, 295–302. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Concensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scienfitic Association of Probiotics and Prebiotics (ISAPP) concensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Concensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Concensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelli, N.; Sola-Oriol, D.; Perez, J.E. Phytogenic Feed Additives in Poultry: Achievements, Prospective and Challenges. Animals 2021, 11, 3471. [Google Scholar] [CrossRef] [PubMed]
- Tellez, G.; Pixley, C.; Wolfenden, R.E.; Layton, S.L.; Hargis, B.M. Probiotics/Direct Fed Microbials for Salmonella Control in Poultry. Food Res. Int. 2012, 45, 628–633. [Google Scholar] [CrossRef]
- Johny, A.K.; Venkitanarayanan, K. Chatper 17—Preharvest Food Safety—Potential Use of Plant-Derived Compounds in Layer Chickens. In Producing Safe Eggs; Ricke, S.C., Gast, R.K., Eds.; Elsevier: London, UK, 2016; pp. 347–372. [Google Scholar]
- Adhikari, P.; Lee, C.H.; Cosby, D.E.; Cox, N.A.; Kim, W.K. Effect of Probiotics on Fecal Excretion, Colonization in Internal Organs and Immune Gene Expression in the Ileum of Laying Hens Challenged with Salmonella Enteritidis. Poult. Sci. 2019, 98, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Micciche, A.C.; Foley, S.L.; Pavlidis, H.O.; McIntyre, D.R.; Ricke, S.C. A Review of Prebiotics Against Salmonella in Poultry: Current and Future Potential for Microbiome Research Applications. Front. Vet. Sci. 2018, 5, 191. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.I.; Sadekuzzaman, M.; Sang-Do, H. Probiotics as Potential Alternative Biocontrol Agents in teh Agriculture and Food Industries: A Review. Food Res. Int. 2017, 100, 63–73. [Google Scholar] [CrossRef]
- Wegh, C.A.; Geerlings, S.Y.; Knol, J.; Roeselers, G.; Belzer, C. Postbiotics and Their Potential Applications in Early Life Nutrition and Beyond. Int. J. Mol. Sci. 2019, 20, 4673. [Google Scholar] [CrossRef] [Green Version]
- Nelson, J.R.; McIntyre, D.R.; Pavlidis, H.O.; Archer, G.S. Reducing Stress Susceptibility of Broiler Chickens by Supplementing a Yeast Fermentation Product in the Feed or Drinking Water. Animals 2018, 8, 173. [Google Scholar] [CrossRef] [Green Version]
- Lensing, M.; van der Klis, J.D.; Yoon, I.; Moore, D.T. Efficacy of Saccharomyces cerevisiae Fermentation Product on Intestinal Health and Productivity of Coccidian-Challenged Laying Hens. Poult. Sci. 2012, 91, 1590–1597. [Google Scholar] [CrossRef]
- Labib, Z.M.; Elsamadony, H.A.; El Gabaly, L.S.; Zoghbi, A.F. Immunopathological Studies on Ducks Experimentally Infected with Duck Virus Enteritis and Salmonella Enteritidis with Special References to The Effect of XPC Prebiotic. Zagazig Vet. J. 2014, 42, 41–62. [Google Scholar] [CrossRef] [Green Version]
- Firman, J.D.; Moore, D.; McIntyre, D. Effects of Dietary Inclusion of a Saccharomyces cerevisiae Fermentation Product on Performance and Gut Characteristics of Male Turkeys to Market Weight. Int. J. Poult. Sci. 2013, 12, 141–143. [Google Scholar] [CrossRef] [Green Version]
- Price, P.T.; Byrd, J.A.; Alvarado, C.Z.; Pavlidis, H.O.; McIntyre, D.R.; Archer, G.S. Utilizing Original XPC in Feed to Reduce Stress Susceptibility of Broilers. Poult. Sci. 2018, 97, 855–859. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhang, H.J.; Wu, S.G.; Yu, S.H.; Yoon, I.; Moore, D.; Gao, Y.P.; Yan, H.J.; Qi, G.H. Effect of Saccharomyces cerevisiae Fermentation Product on Immune Functions of Broilers Challenged with Eimeria tenella. Poult. Sci. 2009, 88, 2141–2151. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, H.J.; Wu, S.G.; Yu, S.H.; Yoon, I.; Quigley, J.; Gao, Y.P.; Qi, G.H. Effects of Yeast Culture in Broiler Diets on Performance and Immunomodulatory Functions. Poult. Sci. 2008, 87, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Roto, S.; Pavlidis, H.; McIntyre, D.; Striplin, K.; Brammer, L.; Ricke, S. Effects of feeding Original XPC to broilers with a live coccidiosis vaccine under industrial conditions: Part 2. Cecal microbiota analysis. Poult. Sci. 2017, 96, 2400–2411. [Google Scholar] [CrossRef]
- Roto, S.; Park, S.; Lee, S.; Kaldhone, P.; Pavlidis, H.; Frankenbach, S.; McIntyre, D.; Striplin, K.; Brammer, L.; Ricke, S. Effects of feeding Original XPC to broilers with a live coccidiosis-vaccine under industry conditions: Part 1. Growth performance and Salmonella inhibition. Poult. Sci. 2017, 96, 1831–1837. [Google Scholar] [CrossRef]
- Chaney, W.E.; Naqvi, S.A.; Gutierrez, M.; Gernat, A.; Johnson, T.J.; Petry, D. Dietary Inclusion of a Saccharomyces cerevisiae-Derived Postbiotic Is Associated with Lower Salmonella enterica Burden in Broiler Chickens on a Commercial Farm in Honduras. Microorganisms 2022, 10, 544. [Google Scholar] [CrossRef]
- Kalani, M.; Rahimi, S.; Zahraei Salehi, T.; Hajiaghaee, R.; Behnamifar, A. Comparison the effects of probiotic and prebiotic as antibiotic alternatives on Salmonella colonization, performance, and egg quality in laying hens challenged with Salmonella enterica serotype Enteritidis. Iran. J. Vet. Res. Shiraz Univ. 2022, 23, 154–162. [Google Scholar] [CrossRef]
- Gingerich, E.; Frana, T.; Logue, C.M.; Smith, D.P.; Pavlidis, H.O.; Chaney, W.E. Effect of feeding a postbiotic derived from Saccharomyces cerevisiae fermentation as a preharvest food safety hurdle for reducing Salmonella Enteritidis in the ceca of layer pullets. J. Food Prot. 2021, 84, 275–280. [Google Scholar] [CrossRef]
- Price, P.T.; Gaydos, T.A.; Berghaus, R.D.; Baxter, V.; Hofacre, C.L.; Sims, M.D. Salmonella Enteritidis reduction in layer ceca with a Bacillus probiotic. Vet. World 2020, 13, 184–187. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Guide for the Care and Use of Laboratory Animals: Eighth Edition; The National Academis Press: Washington, DC, USA, 2011. [Google Scholar]
- Immerseel, F.; Cauwerts, K.; Devriese, L.A.; Haesebrouck, F.; Ducatelle, R. Feed Additives to Control Salmonella in Poultry. World’s Poult. Sci. J. 2002, 58, 501–513. [Google Scholar] [CrossRef]
- Al-Khalaifah, H.S. Benefits of probiotics and/or prebiotics for antibiotic-reduced poultry. Poult. Sci. 2018, 97, 3807–3815. [Google Scholar] [CrossRef]
- Rogers, A.W.L.; Tsolis, R.M.; Baumler, A.J. Salmonella versus the microbiome. Microbiol. Mol. Biol. Rev. 2020, 85, e00027-19. [Google Scholar] [CrossRef] [PubMed]
- Bucher, M.G.; Zwirzitz, B.; Oladeinde, A.; Cook, K.L.; Plymel, C.; Zock, G.; Lakin, S.; Aggrey, S.E.; Ritz, C.; Looft, T.; et al. Reused poultry litter microbiome with competitive exclusion potential against Salmonella Heidelberg. J. Environ. Qual. 2019, 49, 869–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubinelli, P.; Roto, S.; Ae Kim, S.; Hong Park, S.; Pavlidis, H.; McIntyre, D.; Ricke, S. Reduction of Salmonella Typhimurium by Fermentation Metabolites of Diamond V Original XPC in an In Vitro Anaerobic Mixed Chicken Cecal Culture. Front. Vet. Sci. 2016, 3, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofacre, C.L.; Berghaus, R.D.; Jalukar, S.; Mathis, G.F.; SMith, J.A. Effect of a Yeast Cell Wall Preparation on Cecal and Ovarian Colonization with Salmonella Enteritidis in Commercial Layers. J. Appl. Poult. Res. 2018, 27, 453–460. [Google Scholar] [CrossRef]
- Seifi, S.; Partovi, R.; Khoshbakht, R.; Gilani, A. The effect of Prebiotic Administration in the Diet at Unusual Times on Fecal Shedding of Salmonella Enteritidis and Meat Characteristics of Broilers. Int. J. Enteric Pathog. 2019, 7, 75–79. [Google Scholar] [CrossRef]
- Price, P.T.; Gaydos, T.; Legendre, H.; Krehling, J.; Macklin, K.; Padgett, J.C. Production Layer Salmonella Enteritidis Control through Dry Fed Pre & Probiotic Products. Braz. J. Poult. Sci. 2021, 23, 1–6. [Google Scholar] [CrossRef]
- Suganuma, K.; Hamasaki, T.; Hamaoka, T. Effect of dietary direct-fed microbial and yeast cell walls on cecal digesta microbiota of layer chicks inoculated with nalidixic acid resistant Salmonella Enteritidis. Poult. Sci. 2021, 100, 101385. [Google Scholar] [CrossRef]
- Gast, R.K. Understanding Salmonella enteritidis in laying chickens: The contributions of experimental infections. Int. J. Food Microbiol. 1994, 21, 107–116. [Google Scholar] [CrossRef]
- Gast, R.K.; Holt, P.S. Persistence of Salmonella Enteritidis from One Day of Age Until Maturity in Experimentally Infected Layer Chickens. Poult. Sci. 1998, 77, 1759–1762. [Google Scholar] [CrossRef] [PubMed]
- Girgis, G.; Powell, M.; Youssef, M.; Graugnard, D.E.; King, W.D.; Dawson, K.A. Effects of a mannan-rich yeast cell wall-derived preparation on cecal concentrations and tissue prevalence of Salmonella Enteritidis in layer chickens. PLoS ONE 2020, 15, e0232088. [Google Scholar] [CrossRef] [PubMed]
- Gast, R.K.; Guraya, R.; Holt, P.S. Frequency and Persistence of Fecal Shedding Following Exposure of Laying Hens to Different Oral Doses of Salmonella Enteritidis. Int. J. Poult. Sci. 2011, 10, 750–756. [Google Scholar] [CrossRef] [Green Version]
- Gast, R.K.; Guraya, R.; Jones, D.R.; Anderson, K.E. Horizontal Transmission of Salmonella Enteritidis in Experimentally Infected Laying Hens Housed in Conventional or Enriched Cages. Poult. Sci. 2014, 93, 3145–3151. [Google Scholar] [CrossRef]


{kind=link}
| Group | SCFP Inclusion | SE Challenge | Number of Birds | Number of Cages | Total Number of Birds/Treatment |
|---|---|---|---|---|---|
| Treatment | + | Direct | 120 | 60 | 240 |
| Indirect | 120 | ||||
| Control | − | Direct | 120 | 60 | 240 |
| Indirect | 120 |
| Ingredient | 0–7 Weeks Starter (20.3% Protein) | 8–18 Weeks Grower (18.0% Protein) |
|---|---|---|
| Ground corn | 1137.52 | 1269.63 |
| Soybean meal | 670.87 | 561.91 |
| Calcium carbonate | 38.93 | 34.71 |
| 21% monosodium phosphate | 35.76 | 34.43 |
| Salt | 10.05 | 9.35 |
| Soybean oil | 78.19 | 69.51 |
| Choline chloride | 1.13 | 1.21 |
| DL methionine | 9.47 | 3.12 |
| Lysine | 13.59 | 11.64 |
| Vitamin mix | 2.00 | 2.00 |
| Test article (SCFP)—test diet only | 2.50 * | 2.50 * |
| Total (lbs.) | 2000 | 2000 |
| Days | Direct Challenge | Indirect Challenge | ||
|---|---|---|---|---|
| Post-Challenge | CON | SCFP | CON | SCFP |
| 3 DPC | 97.5% (39/40) a | 100% (40/40) a | 40.0% (16/40) b | 45.0% (18/40) b |
| 7 DPC | 97.5% (39/40) a | 97.5% (39/40) a | 72.5% (29/40) b | 50.0% (20/40) c |
| 14 DPC | 90.0% (36/40) a | 80.0% (32/40) a | 52.5% (21/40) b | 42.5% (17/40) b |
| Days | Direct Challenge | Indirect Challenge | ||
|---|---|---|---|---|
| Post-Challenge | CON | SCFP | CON | SCFP |
| 3 DPC | 35.0% (14/40) a | 40.0% (16/40) a | 2.5% (1/40) b | 2.5% (1/40) b |
| 7 DPC | 30.0% (12/40) a | 32.5% (13/40) a | 5.0% (2/40) b | 5.0% (2/40) b |
| 14 DPC | 10.0% (4/40)a | 2.5% (1/40) a | 0.0% (0/40) a | 2.5% (1/40) a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaney, W.E.; McBride, H.; Girgis, G. Effect of a Saccharomyces cerevisiae Postbiotic Feed Additive on Salmonella Enteritidis Colonization of Cecal and Ovarian Tissues in Directly Challenged and Horizontally Exposed Layer Pullets. Animals 2023, 13, 1186. https://doi.org/10.3390/ani13071186
Chaney WE, McBride H, Girgis G. Effect of a Saccharomyces cerevisiae Postbiotic Feed Additive on Salmonella Enteritidis Colonization of Cecal and Ovarian Tissues in Directly Challenged and Horizontally Exposed Layer Pullets. Animals. 2023; 13(7):1186. https://doi.org/10.3390/ani13071186
Chicago/Turabian StyleChaney, W. Evan, Hannah McBride, and George Girgis. 2023. "Effect of a Saccharomyces cerevisiae Postbiotic Feed Additive on Salmonella Enteritidis Colonization of Cecal and Ovarian Tissues in Directly Challenged and Horizontally Exposed Layer Pullets" Animals 13, no. 7: 1186. https://doi.org/10.3390/ani13071186