


Oral Vaccination of Largemouth Bass (Micropterus salmoides) against Largemouth Bass Ranavirus (LMBV) Using Yeast Surface Display Technology
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus, Cell Lines, and Fish
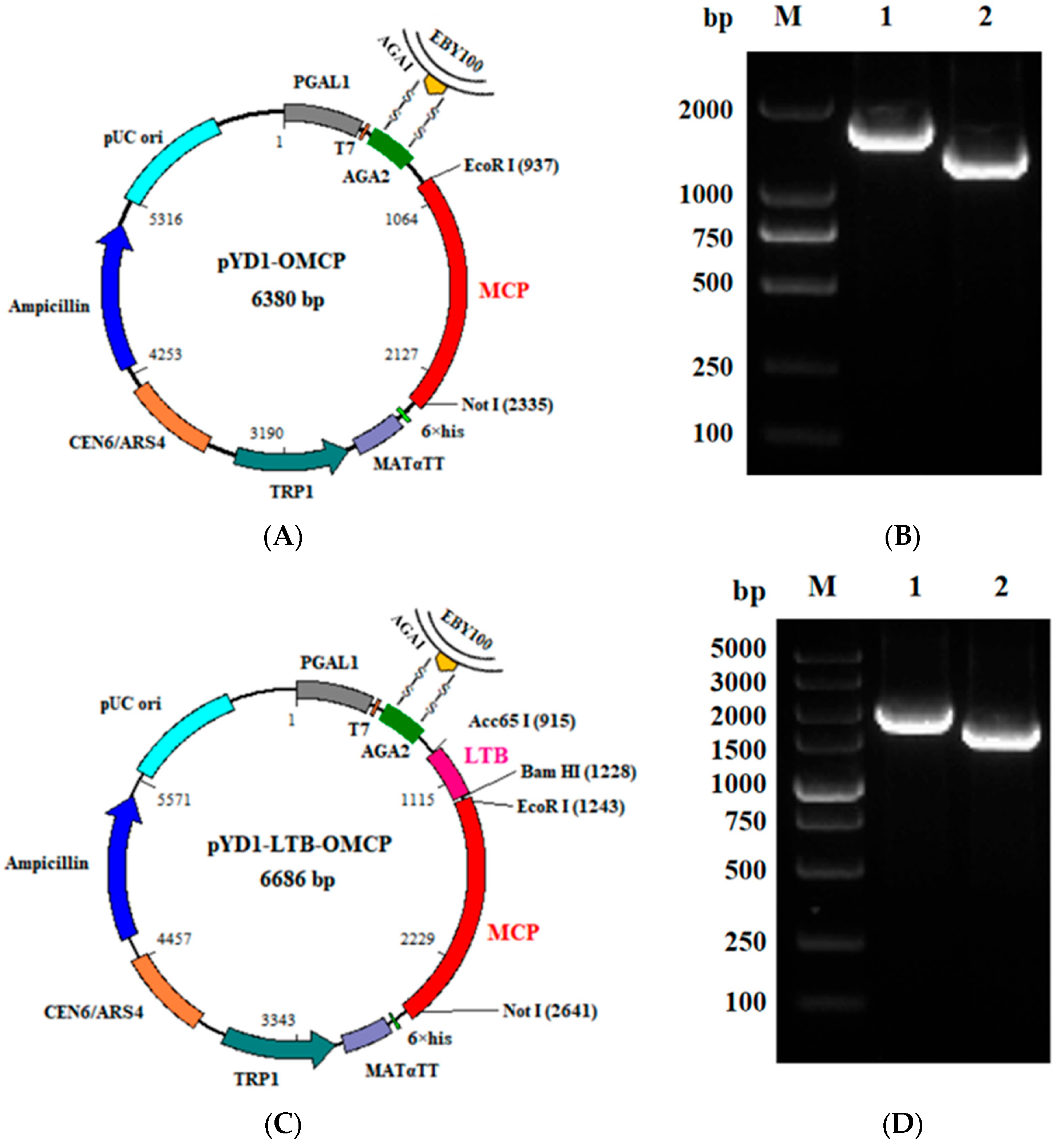
2.2. Strains and Plasmids
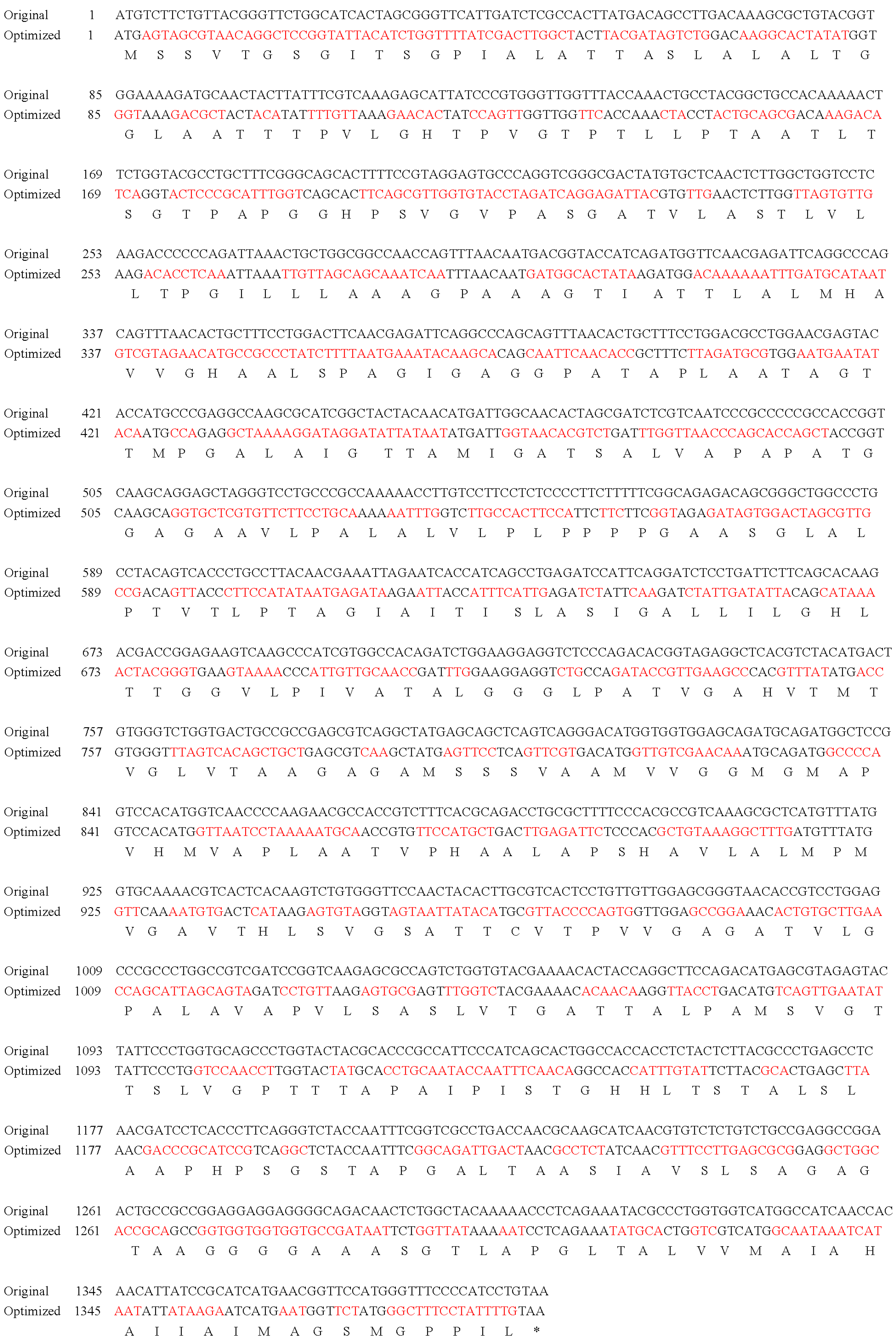
2.3. Codon Optimization and Gene Synthesis
2.4. Construction of the Recombinant Plasmids
2.5. Screening and Induction of Expression in Vaccine Strains
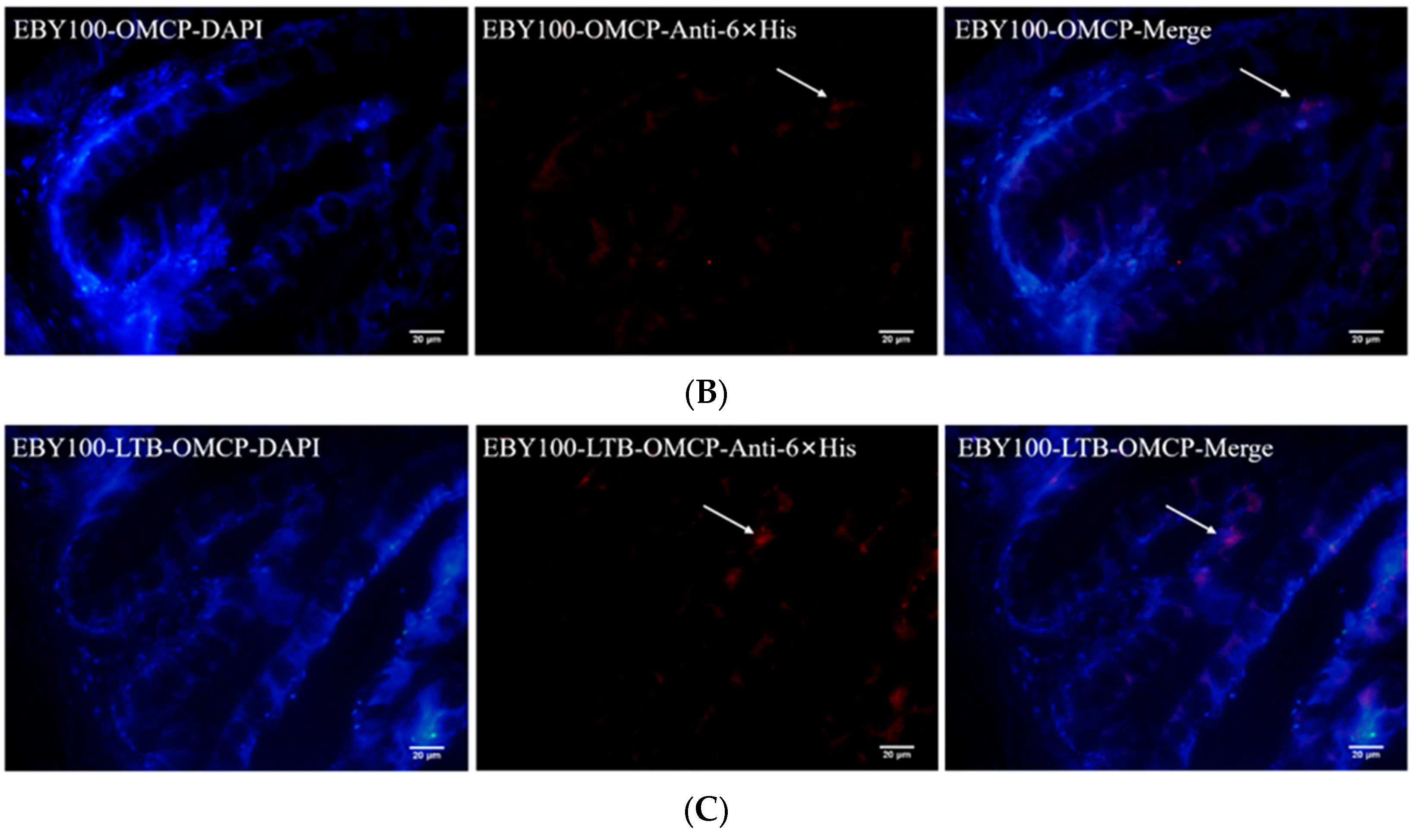
2.6. Immunofluorescence Analysis
2.7. Detection of Recombinant Yeast in the Intestines
2.8. Oral Immunization and Sample Collections
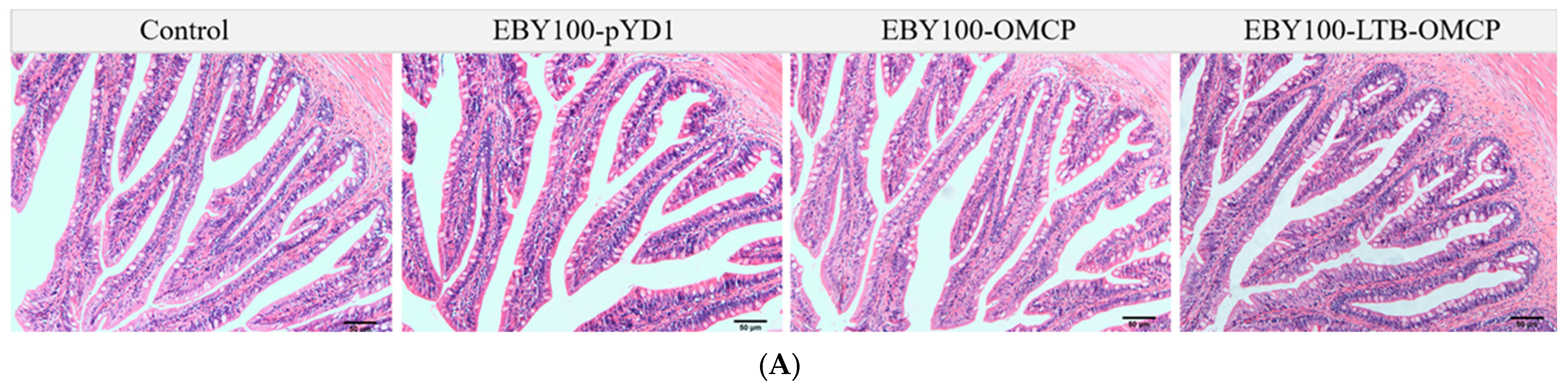
2.9. Vaccine Safety Evaluation
2.10. Determination of Immune-Related Enzyme Activity
2.11. Expression of Immune-Related Genes
2.12. Serum Neutralization Antibody Assay
2.13. LMBV Challenge
2.14. Data Analysis
3. Results
3.1. MCP Codon Optimization
3.2. The Recombinant Protein Was Displayed on the Surface of S. cerevisiae
3.3. Recombinant Yeast Was Detected in the Second Intestine
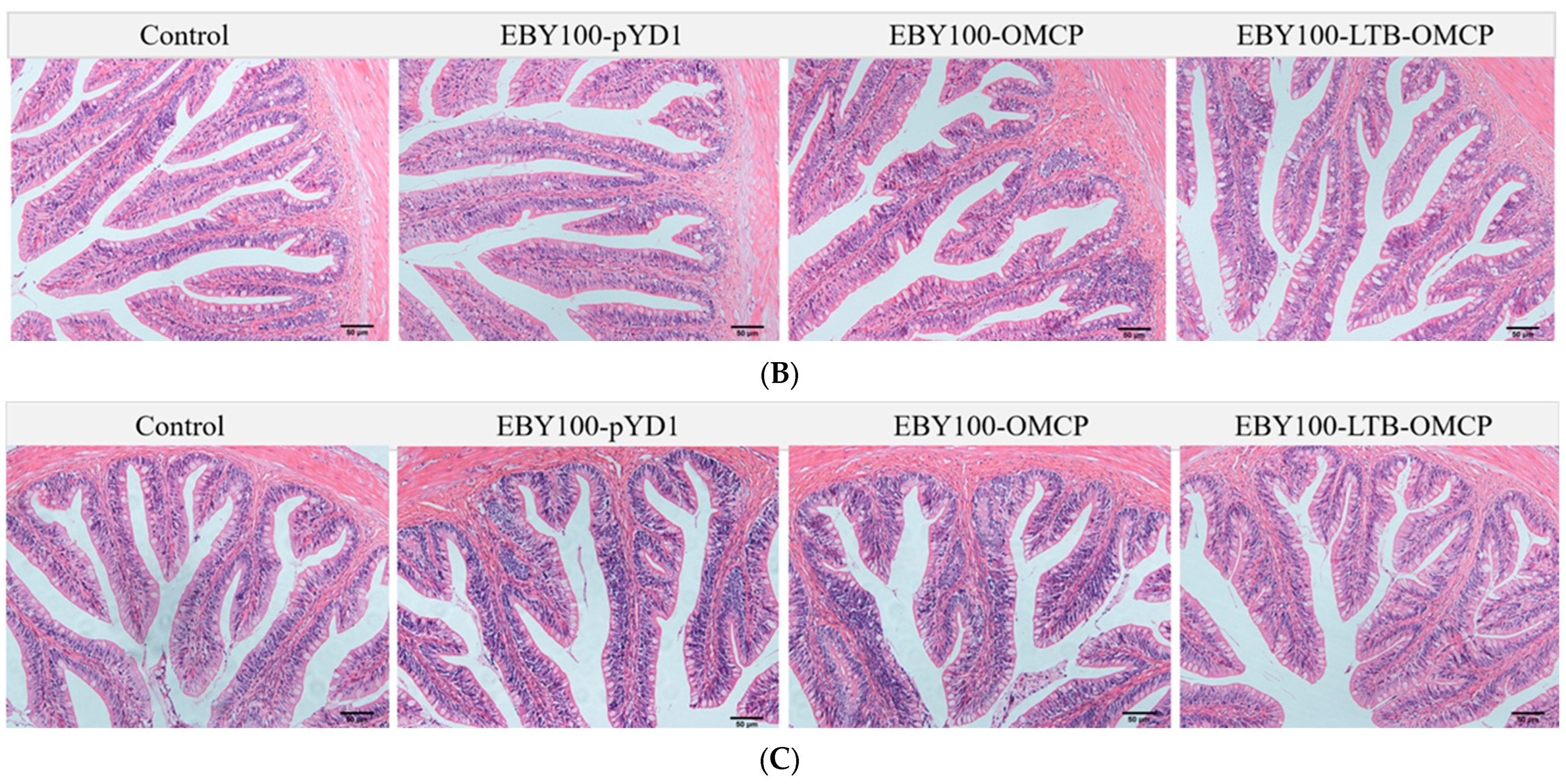
3.4. Safety Evaluation of Oral Vaccine
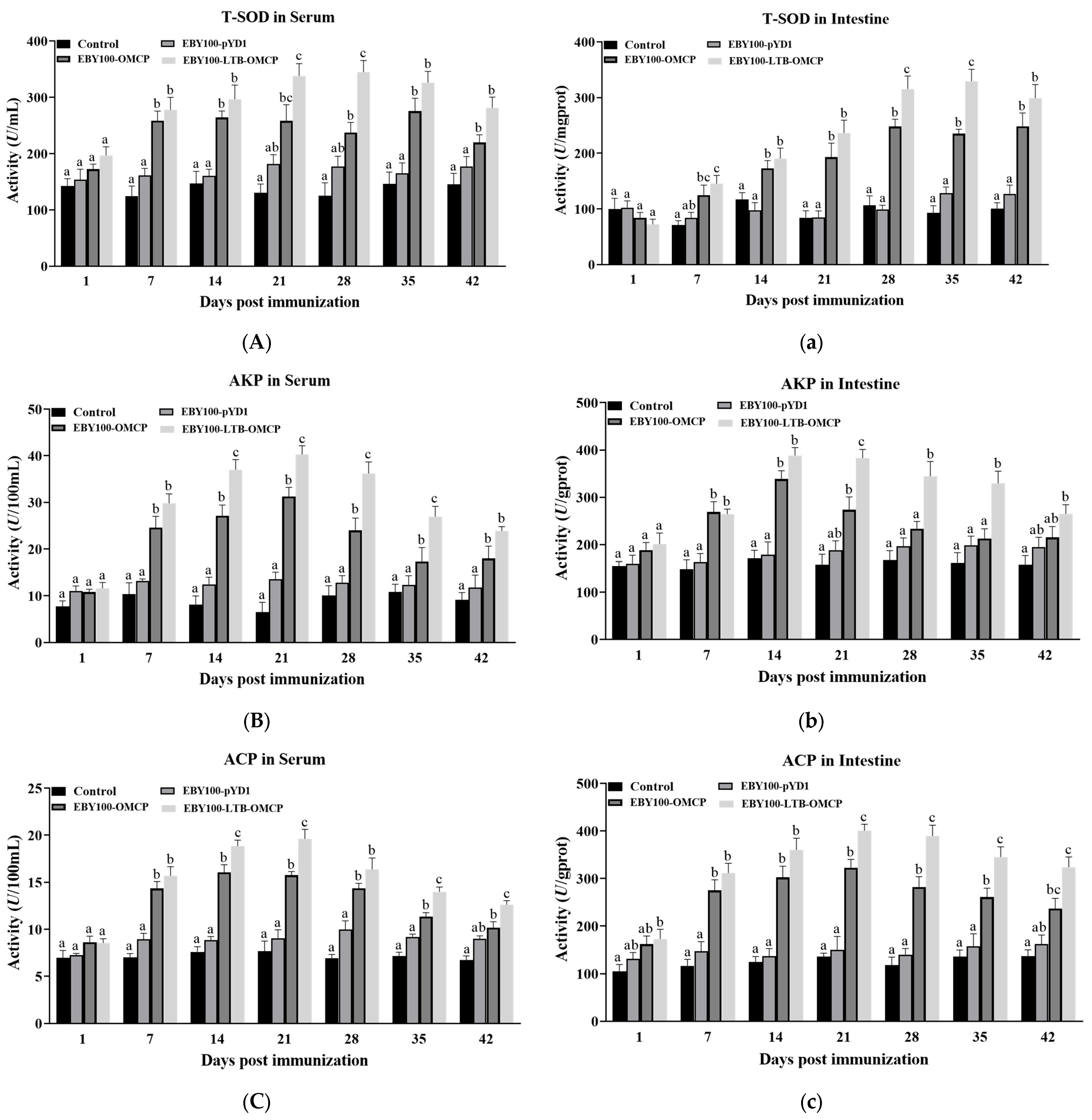
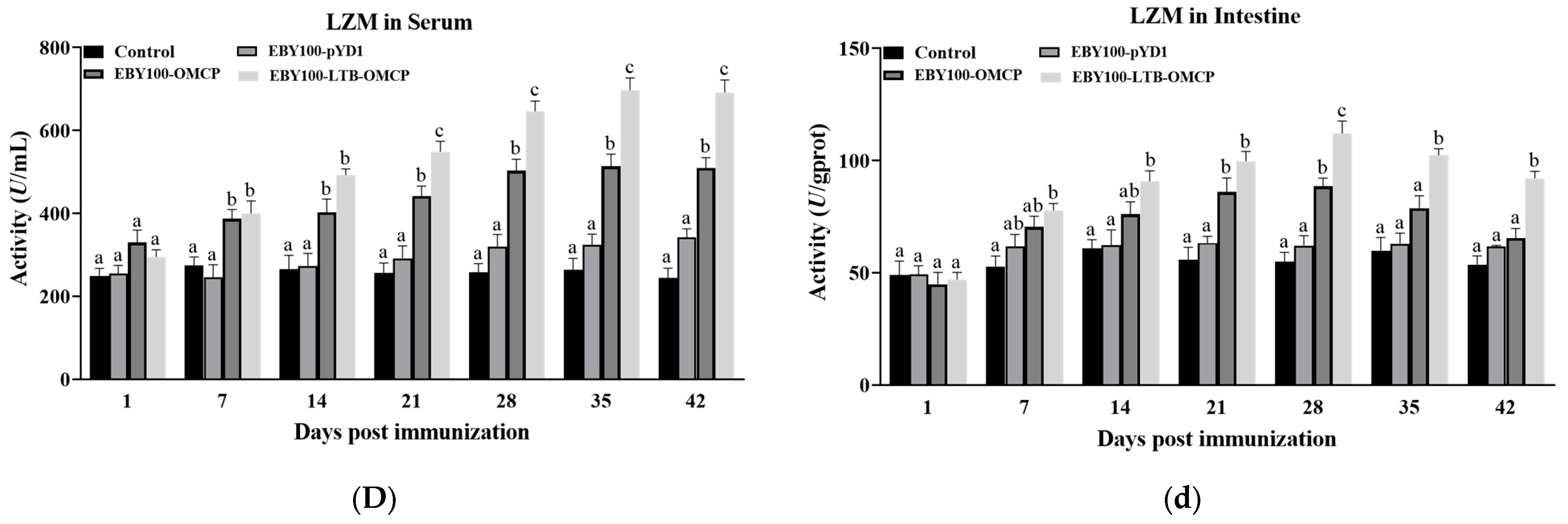
3.5. Detection of Immune-Related Enzyme Activity
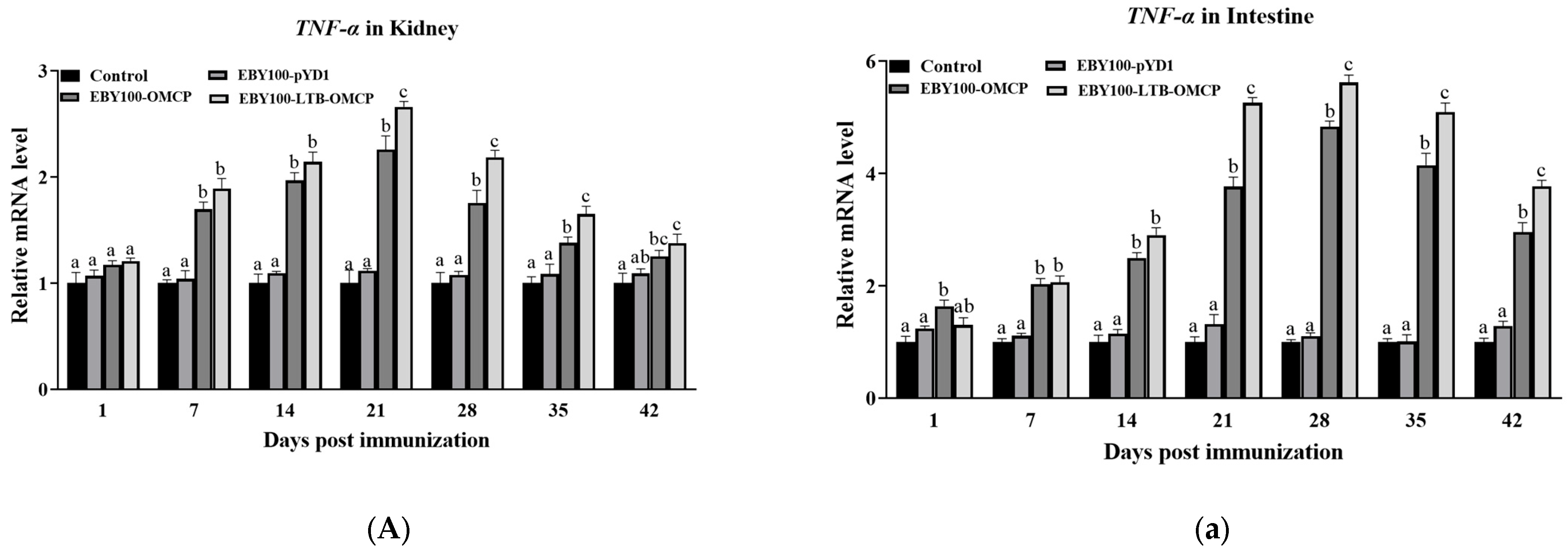
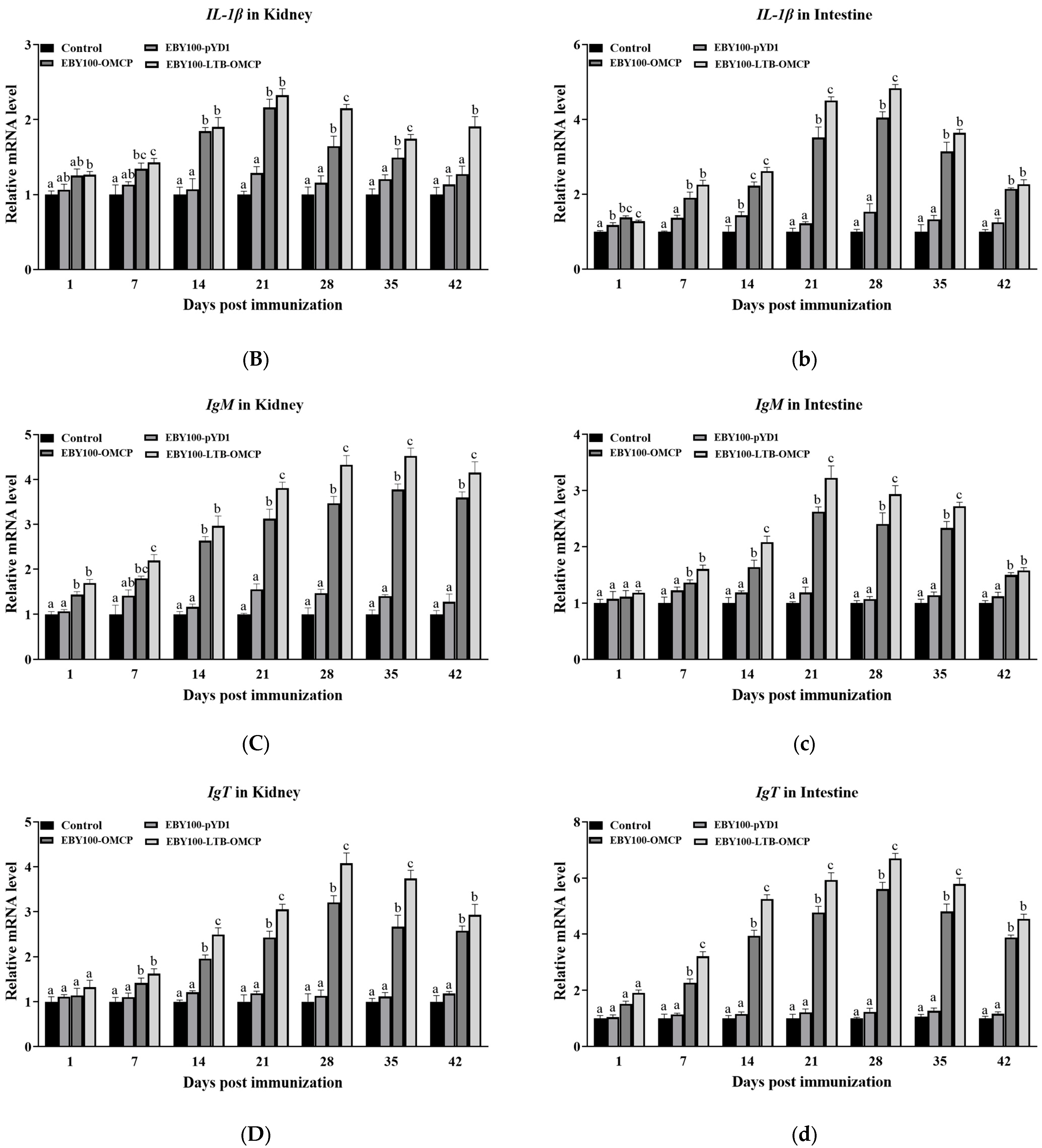
3.6. Expression of Immune-Related Genes
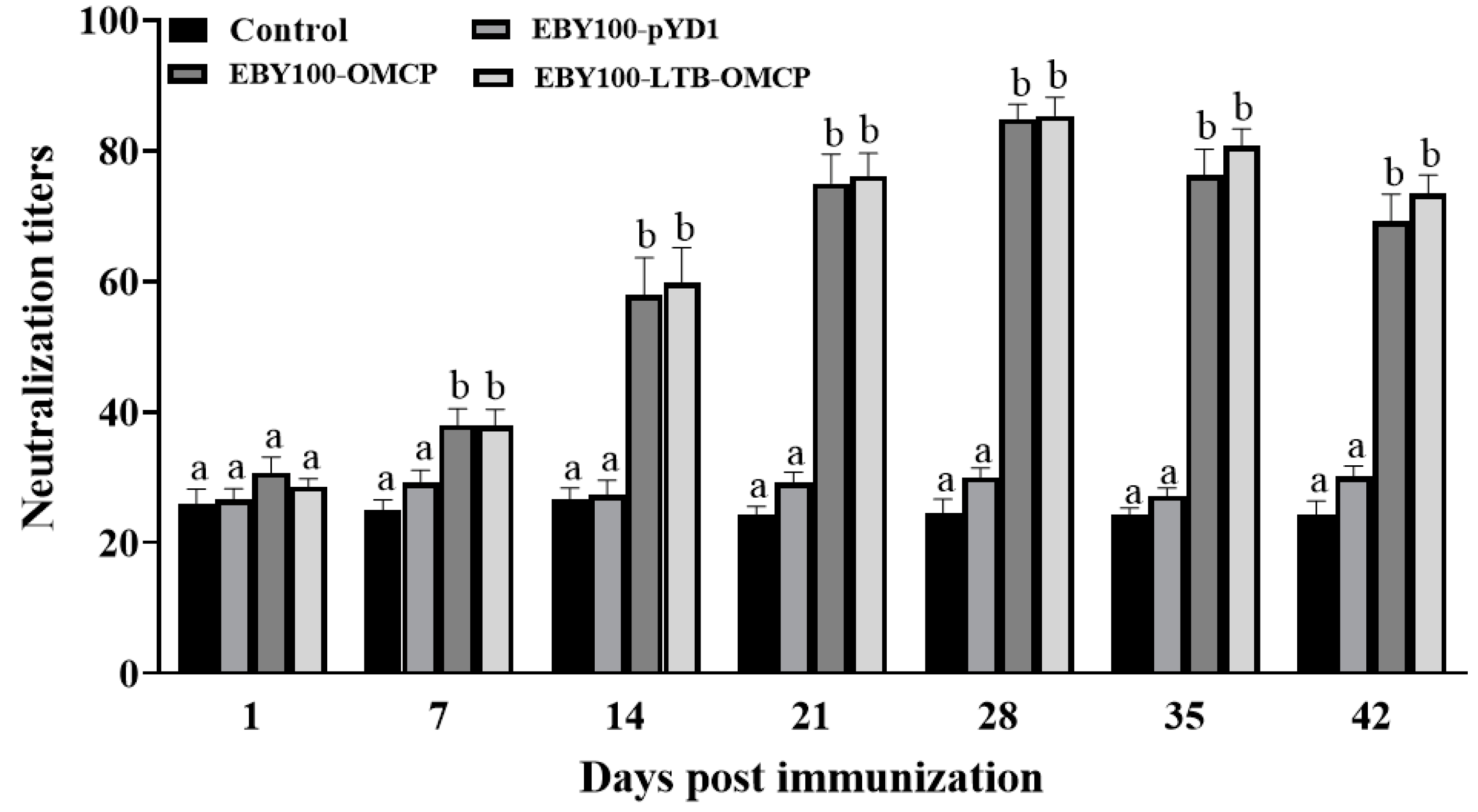
3.7. Serum Antibody Levels
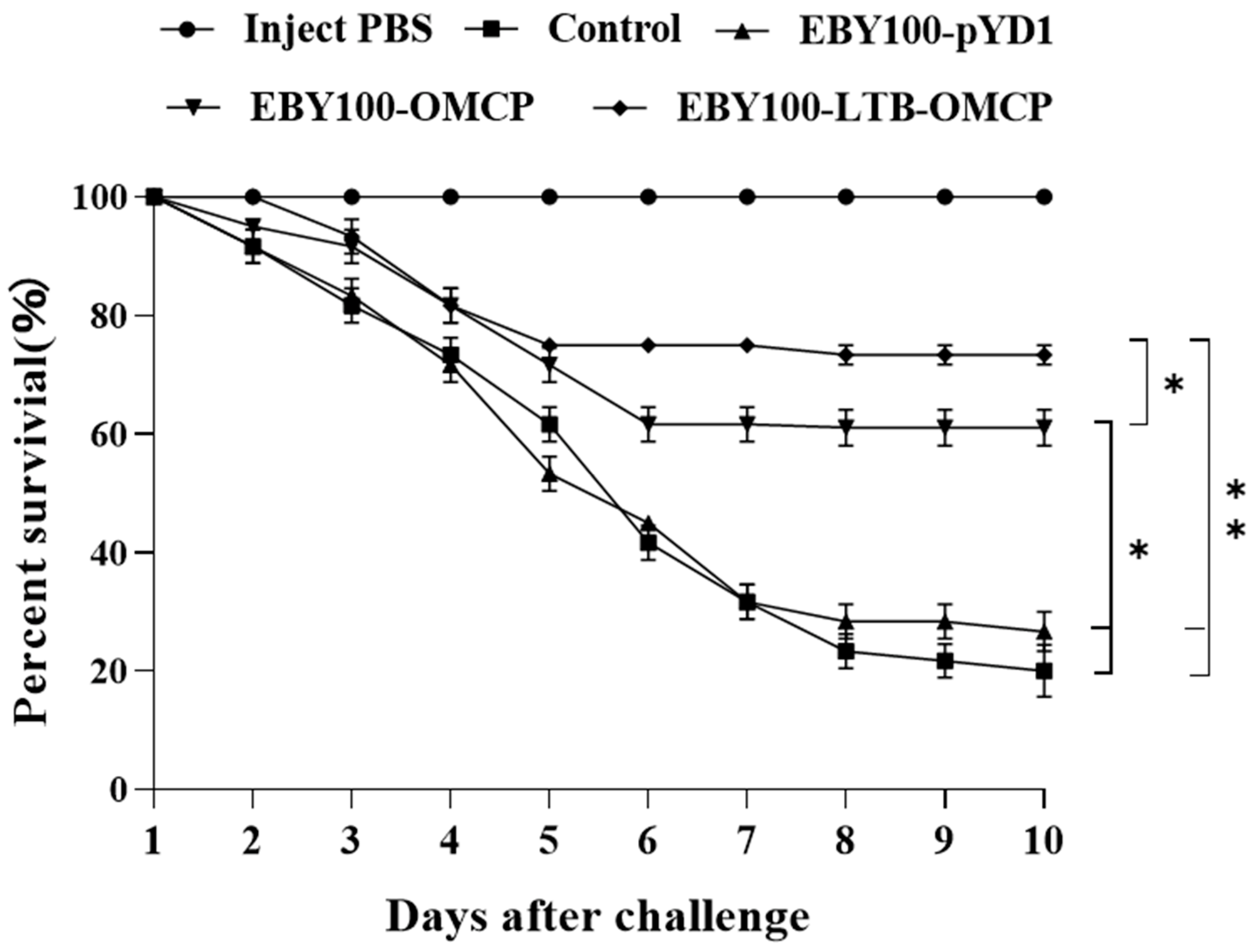
3.8. Oral Vaccine Protection against LMBV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bai, J.; Lutz-Carrillo, D.J.; Quan, Y.; Liang, S. Taxonomic status and genetic diversity of cultured largemouth bass Micropterus salmoides in China. Aquaculture 2008, 278, 27–30. [Google Scholar] [CrossRef]
- Hussein, G.H.G.; Chen, M.; Qi, P.-P.; Cui, Q.-K.; Yu, Y.; Hu, W.-H.; Tian, Y.; Fan, Q.-X.; Gao, Z.-X.; Feng, M.-W.; et al. Aquaculture industry development, annual price analysis and out-of-season spawning in largemouth bass Micropterus salmoides. Aquaculture 2020, 519, 734901. [Google Scholar] [CrossRef]
- Deng, G.; Li, S.; Xie, J.; Bai, J.; Chen, K.; Ma, D.; Jiang, X.; Lao, H.; Yu, L. Characterization of a ranavirus isolated from cultured largemouth bass (Micropterus salmoides) in China. Aquaculture 2011, 312, 198–204. [Google Scholar] [CrossRef]
- Fogelson, S.B.; Petty, B.D.; Reichley, S.R.; Ware, C.; Bowser, P.R.; Crim, M.J.; Getchell, R.G.; Sams, K.L.; Marquis, H.; Griffin, M.J. Histologic and molecular characterization of Edwardsiella piscicida infection in largemouth bass (Micropterus salmoides). J. Vet. Diagn. Investig. 2016, 28, 338–344. [Google Scholar] [CrossRef]
- Jiang, B.; Lu, G.; Du, J.; Wang, J.; Hu, Y.; Su, Y.; Li, A. First report of trypanosomiasis in farmed largemouth bass (Micropterus salmoides) from China: Pathological evaluation and taxonomic status. Parasitol. Res. 2019, 118, 1731–1739. [Google Scholar] [CrossRef]
- Mao, J.; Wang, J.; Chinchar, G.; Chinchar, V. Molecular characterization of a ranavirus isolated from largemouth bass Micropterus salmoides. Dis. Aquat. 1999, 37, 107–114. [Google Scholar] [CrossRef]
- Qin, Q.W.; Chang, S.F.; Ngoh-Lim, G.H.; Gibson-Kueh, S.; Shi, C.; Lam, T.J. Characterization of a novel ranavirus isolated from grouper Epinephelus tauvina. Dis. Aquat. Org. 2003, 53, 1–9. [Google Scholar] [CrossRef]
- Plumb, J.A.; Grizzle, J.M.; Young, H.E.; Noyes, A.D.; Lamprecht, S. An iridovirus isolated from wild largemouth bass. J. Aquat. Anim. Health 1996, 8, 265–270. [Google Scholar] [CrossRef]
- Groocock, G.H.; Grimmett, S.G.; Getchell, R.G.; Wooster, G.A.; Bowser, P.R. A survey to determine the presence and distribution of largemouth bass virus in wild freshwater bass in New York State. J. Aquat. Anim. Health 2008, 20, 158–164. [Google Scholar] [CrossRef]
- Grizzle, J.M.; Altinok, I.; Noyes, A.D. PCR method for detection of largemouth bass virus. Dis. Aquat. Org. 2003, 54, 29–33. [Google Scholar] [CrossRef]
- Southard, G.M.; Fries, L.T.; Terre, D.R. Largemouth bass virus in Texas: Distribution and management issues. J. Aquat. Anim. Health 2009, 21, 36–42. [Google Scholar] [CrossRef]
- Neal, J.W.; Eggleton, M.A.; Goodwin, A.E. The effects of largemouth bass virus on a quality largemouth bass population in Arkansas. J. Wildl. Dis. 2009, 45, 766–771. [Google Scholar] [CrossRef]
- Zhao, R.; Geng, Y.; Qin, Z.; Wang, K.; Ouyang, P.; Chen, D.; Huang, X.; Zuo, Z.; He, C.; Guo, H.; et al. A new ranavirus of the Santee-Cooper group invades largemouth bass (Micropterus salmoides) culture in southwest China. Aquaculture 2020, 526, 734988. [Google Scholar] [CrossRef]
- Jiang, N.; Shen, J.; Zhou, Y.; Liu, W.; Meng, Y.; Li, Y.; Xue, M.; Xu, C.; Fan, Y. Development of a droplet digital PCR method for the sensitive detection and quantification of largemouth bass ranavirus. J. Fish Dis. 2023, 46, 91–98. [Google Scholar] [CrossRef]
- Mondal, H.; Thomas, J. A review on the recent advances and application of vaccines against fish pathogens in aquaculture. Aquac. Int. 2022, 30, 1971–2000. [Google Scholar] [CrossRef]
- Kim, T.J.; Jang, E.J.; Lee, J.I. Vaccination of rock bream, Oplegnathus fasciatus (Temminck & Schlegel), using a recombinant major capsid protein of fish iridovirus. J. Fish Dis. 2008, 31, 547–551. [Google Scholar] [CrossRef]
- Liu, H.I.; Chiou, P.P.; Gong, H.Y.; Chou, H.Y. Cloning of the Major Capsid Protein (MCP) of Grouper Iridovirus of Taiwan (TGIV) and Preliminary Evaluation of a Recombinant MCP Vaccine against TGIV. Int. J. Mol. Sci. 2015, 16, 28647–28656. [Google Scholar] [CrossRef]
- Yi, W.; Zhang, X.; Zeng, K.; Xie, D.; Song, C.; Tam, K.; Liu, Z.; Zhou, T.; Li, W. Construction of a DNA vaccine and its protective effect on largemouth bass (Micropterus salmoides) challenged with largemouth bass virus (LMBV). Fish Shellfish Immunol. 2020, 106, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.Y.; Zhang, C.S.; Yuan, X.M.; Huang, L.; Hu, D.Y.; Yu, Z.; Yin, W.L.; Lin, L.Y.; Pan, X.Y.; Yang, G.L.; et al. Oral Vaccination with Recombinant Pichia pastoris Expressing Iridovirus Major Capsid Protein Elicits Protective Immunity in Largemouth Bass (Micropterus salmoides). Front. Immunol. 2022, 13, 852300. [Google Scholar] [CrossRef]
- Tan, Y.; Chen, L.; Li, K.; Lou, B.; Liu, Y.; Liu, Z. Yeast as carrier for drug delivery and vaccine construction. J. Control. Release 2022, 346, 358–379. [Google Scholar] [CrossRef]
- Porro, D.; Branduardi, P. Yeast cell factory: Fishing for the best one or engineering it? Microb. Cell Fact. 2009, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Schreuder, M.P.; Deen, C.; Boersma, W.J.; Pouwels, P.H.; Klis, F.M. Yeast expressing hepatitis B virus surface antigen determinants on its surface: Implications for a possible oral vaccine. Vaccine 1996, 14, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Stander, J.; Mbewana, S.; Meyers, A.E. Plant-Derived Human Vaccines: Recent Developments. BioDrugs 2022, 36, 573–589. [Google Scholar] [CrossRef]
- Hemmati, F.; Hemmati-Dinarvand, M.; Karimzade, M.; Rutkowska, D.; Eskandari, M.H.; Khanizadeh, S.; Afsharifar, A. Plantderived VLP: A worthy platform to produce vaccine against SARS-CoV-2. Biotechnol. Lett. 2022, 44, 45–57. [Google Scholar] [CrossRef]
- Di Pasquale, A.; Preiss, S.; Tavares Da Silva, F.; Garcon, N. Vaccine Adjuvants: From 1920 to 2015 and Beyond. Vaccines 2015, 3, 320–343. [Google Scholar] [CrossRef]
- Ma, Y. Recent advances in nontoxic Escherichia coli heat-labile toxin and its derivative adjuvants. Expert Rev. Vaccines 2016, 15, 1361–1371. [Google Scholar] [CrossRef]
- Li, W.; Han, D.; Liang, S.; Zhong, Z.; Li, X.; Wen, J.; Lin, H.; Wang, L.; Li, X.; Zhong, X.; et al. Molecular mechanism for the effects of E. coli heat-labile enterotoxin on mouse embryo survival. Reprod. Toxicol. 2014, 45, 31–38. [Google Scholar] [CrossRef]
- Huang, L.; Fan, Y.; Zhou, Y.; Jiang, N.; Liu, W.; Meng, Y.; Zeng, L. Cloning, sequence analysis and expression profiles of Tolllike receptor 7 from Chinese giant salamander Andrias davidianus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2015, 184, 52–57. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, L.B.; Fan, Y.D.; Luo, X.S.; Xu, J.; Xiao, Y. Plant fusion expression of grass carp reovirus VP6 protein and E.coli LTB subunit construction of body. J. Fish. Sci. China 2011, 18, 1–7. [Google Scholar]
- Cuscino, N.; Fatima, A.; Di Pilato, V.; Bulati, M.; Alfano, C.; Monaca, E.; Di Mento, G.; Di Carlo, D.; Cardinale, F.; Monaco, F.; et al. Computational design and characterization of a multiepitope vaccine against carbapenemase-producing Klebsiella pneumoniae strains, derived from antigens identified through reverse vaccinology. Comput. Struct. Biotechnol. J. 2022, 20, 4446–4463. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.J.; Guo, Z.R.; Ma, R.; Qiu, D.K.; Zhao, Z.; Wang, G.X.; Zhu, B. Immune efficacy of carbon nanotubes recombinant subunit vaccine against largemouth bass ulcerative syndrome virus. Fish Shellfish Immunol. 2020, 100, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Zhai, X.; Wu, Z.; Jiang, H.; Wu, S.; He, Y.; Qin, C.; Xu, Z. Molecular cloning of IgT, IgD and analysis the IgH locus in largemouth bass (Micropterus salmoides) and immune response upon bacterial infection. Aquaculture 2022, 546, 737291. [Google Scholar] [CrossRef]
- Andreu, C.; Del Olmo, M. Yeast arming by the Aga2p system: Effect of growth conditions in galactose on the efficiency of the display and influence of expressing leucine-containing peptides. Appl. Microbiol. Biotechnol. 2013, 97, 9055–9069. [Google Scholar] [CrossRef] [PubMed]
- Mai, W.; Yan, B.; Xin, J. Oral immunizations with Bacillus subtilis spores expressing MCP protein provide protection against red-spotted grouper nervous necrosis virus (RGNNV) infection in juvenile grouper, Epinephelus coioides. Aquaculture 2022, 552, 738008. [Google Scholar] [CrossRef]
- Gao, Y.; Huo, X.; Wang, Z.; Yuan, G.; Liu, X.; Ai, T.; Su, J. Oral Administration of Bacillus subtilis Subunit Vaccine Significantly Enhances the Immune Protection of Grass Carp against GCRV-II Infection. Viruses 2021, 14, 30. [Google Scholar] [CrossRef]
- Xue, M.; Wu, Y.; Hong, Y.; Meng, Y.; Xu, C.; Jiang, N.; Li, Y.; Liu, W.; Fan, Y.; Zhou, Y. Effects of dietary Bacillus amyloliquefaciens on the growth, immune responses, intestinal microbiota composition and disease resistance of yellow catfish, Pelteobagrus fulvidraco. Front. Cell. Infect. Microbiol. 2022, 12, 1047351. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Zhou, Y.; Jiang, N.; Fan, Y.; Lin, G.; Zeng, L. Characterization, expression pattern and antiviral activities of oligoadenylate synthetase in Chinese Giant Salamander, Andrias davidianus. Dev. Comp. Immunol. 2022, 129, 104347. [Google Scholar] [CrossRef]
- Zhou, Y.; Fan, Y.; LaPatra, S.E.; Ma, J.; Xu, J.; Meng, Y.; Jiang, N.; Zeng, L. Protective immunity of a Pichia pastoris expressed recombinant iridovirus major capsid protein in the Chinese giant salamander, Andrias davidianus. Vaccine 2015, 33, 5662–5669. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493497. [Google Scholar] [CrossRef]
- Yao, Y.Y.; Yang, Y.L.; Gao, C.C.; Zhang, F.L.; Xia, R.; Li, D.; Hu, J.; Ran, C.; Zhang, Z.; Liu-Clarke, J.; et al. Surface display system for probiotics and its application in aquaculture. Rev. Aquac. 2020, 12, 2333–2350. [Google Scholar] [CrossRef]
- Kondo, A.; Ueda, M. Yeast cell-surface display—applications of molecular display. Appl. Microbiol. Biotechnol. 2004, 64, 28–40. [Google Scholar] [CrossRef]
- Zhu, K.; Chi, Z.; Li, J.; Zhang, F.; Li, M.; Yasoda, H.N.; Wu, L. The surface display of haemolysin from Vibrio harveyi on yeast cells and their potential applications as live vaccine in marine fish. Vaccine 2006, 24, 6046–6052. [Google Scholar] [CrossRef]
- Gao, T.; Ren, Y.; Li, S.; Lu, X.; Lei, H. Immune response induced by oral administration with a Saccharomyces cerevisiae-based SARS-CoV-2 vaccine in mice. Microb. Cell Fact. 2021, 20, 95. [Google Scholar] [CrossRef]
- Companjen, A.R.; Florack, D.E.; Slootweg, T.; Borst, J.W.; Rombout, J.H. Improved uptake of plant-derived LTB-linked proteins in carp gut and induction of specific humoral immune responses upon infeed delivery. Fish Shellfish Immunol. 2006, 21, 251–260. [Google Scholar] [CrossRef]
- Wan, J.; Wang, T.; Xu, J.; Ouyang, T.; Wang, Q.; Zhang, Y.; Weng, S.; Li, Y.; Wang, Y.; Xin, X.; et al. Novel Japanese encephalitis virus NS1-based vaccine: Truncated NS1 fused with E. coli heat labile enterotoxin B subunit. EBioMedicine 2021, 67, 103353. [Google Scholar] [CrossRef] [PubMed]
- Rombout, J.H.; Abelli, L.; Picchietti, S.; Scapigliati, G.; Kiron, V. Teleost intestinal immunology. Fish Shellfish Immunol. 2011, 31, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Lokka, G.; Austbo, L.; Falk, K.; Bjerkas, I.; Koppang, E.O. Intestinal morphology of the wild Atlantic salmon (Salmo salar). J. Morphol. 2013, 274, 859–876. [Google Scholar] [CrossRef]
- Embregts, C.W.E.; Reyes-Lopez, F.; Pall, A.C.; Stratmann, A.; Tort, L.; Lorenzen, N.; Engell-Sorensen, K.; Wiegertjes, G.F.; Forlenza, M.; Sunyer, J.O.; et al. Pichia pastoris yeast as a vehicle for oral vaccination of larval and adult teleosts. Fish Shellfish Immunol. 2019, 85, 52–60. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, J.; Ma, Y.; Hao, L.; Liang, Z.; Ma, J.; Ke, H.; Li, Y.; Cao, J. Protective immunity against CyHV-3 infection via different prime-boost vaccination regimens using CyHV-3 ORF131-based DNA/protein subunit vaccines in carp Cyprinus carpio var. Jian. Fish Shellfish Immunol. 2020, 98, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Zhu, X.-C.; Liu, X.-Y.; Yuan, K.; Zhang, J.-J.; Gao, H.-H.; Li, J.-N. An oral double-targeted DNA vaccine induces systemic and intestinal mucosal immune responses and confers high protection against Vibrio mimicus in grass carps. Aquaculture 2019, 504, 248–259. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, Z.; Liu, G.Y.; Li, J.; Wang, G.X.; Zhu, B. Immune response and protective effect against spring viremia of carp virus induced by intramuscular vaccination with a SWCNTs-DNA vaccine encoding matrix protein. Fish Shellfish Immunol. 2018, 79, 256–264. [Google Scholar] [CrossRef]
- Biller, J.D.; Takahashi, L.S. Oxidative stress and fish immune system: Phagocytosis and leukocyte respiratory burst activity. An. Acad. Bras. Ciênc. 2018, 90, 3403–3414. [Google Scholar] [CrossRef]
- Xie, J.J.; Liu, Q.Q.; Liao, S.; Fang, H.H.; Yin, P.; Xie, S.W.; Tian, L.X.; Liu, Y.J.; Niu, J. Effects of dietary mixed probiotics on growth, non-specific immunity, intestinal morphology and microbiota of juvenile pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 90, 456–465. [Google Scholar] [CrossRef]
- Petit, J.; Wiegertjes, G.F. Long-lived effects of administering beta-glucans: Indications for trained immunity in fish. Dev. Comp. Immunol. 2016, 64, 93–102. [Google Scholar] [CrossRef]
- Castellheim, A.; Brekke, O.L.; Espevik, T.; Harboe, M.; Mollnes, T.E. Innate immune responses to danger signals in systemic inflammatory response syndrome and sepsis. Scand. J. Immunol. 2009, 69, 479–491. [Google Scholar] [CrossRef]
- De Jesus Andino, F.; Chen, G.; Li, Z.; Grayfer, L.; Robert, J. Susceptibility of Xenopus laevis tadpoles to infection by the ranavirus Frog-Virus 3 correlates with a reduced and delayed innate immune response in comparison with adult frogs. Virology 2012, 432, 435–443. [Google Scholar] [CrossRef]
- Salinas, I. The Mucosal Immune System of Teleost Fish. Biology 2015, 4, 525–539. [Google Scholar] [CrossRef]
- Gomez, D.; Sunyer, J.O.; Salinas, I. The mucosal immune system of fish: The evolution of tolerating commensals while fighting pathogens. Fish Shellfish Immunol. 2013, 35, 1729–1739. [Google Scholar] [CrossRef]
- Stosik, M.P.; Tokarz-Deptula, B.; Deptula, W. Specific humoral immunity in Osteichthyes. Cent. Eur. J. Immunol. 2018, 43, 335–340. [Google Scholar] [CrossRef]
- Dong, Z.R.; Mu, Q.J.; Kong, W.G.; Qin, D.C.; Zhou, Y.; Wang, X.Y.; Cheng, G.F.; Luo, Y.Z.; Ai, T.S.; Xu, Z. Gut mucosal immune responses and protective efficacy of oral yeast Cyprinid herpesvirus 2 (CyHV-2) vaccine in Carassius auratus gibelio. Front. Immunol. 2022, 13, 932722. [Google Scholar] [CrossRef]
- Zhao, J.Z.; Xu, L.M.; Liu, M.; Cao, Y.S.; LaPatra, S.E.; Yin, J.S.; Liu, H.B.; Lu, T.Y. Preliminary study of an oral vaccine against infectious hematopoietic necrosis virus using improved yeast surface display technology. Mol. Immunol. 2017, 85, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, X.; Weng, S.; Zhao, G.; He, J.; Dong, C. Virion-associated viral proteins of a Chinese giant salamander (Andrias davidianus) iridovirus (genus Ranavirus) and functional study of the major capsid protein (MCP). Vet. Microbiol. 2014, 172, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Qi, R.; Chen, C.; Yin, J.; Ma, S.; Shi, W.; Wu, Y.; Ge, J.; Jiang, Y.; Tang, L.; et al. Immunogenicity of recombinant Lactobacillus casei-expressing F4 (K88) fimbrial adhesin FaeG in conjunction with a heat-labile enterotoxin A (LTAK63) and heat-labile enterotoxin B (LTB) of enterotoxigenic Escherichia coli as an oral adjuvant in mice. J. Appl. Microbiol. 2017, 122, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Lei, X.Y.; Zhang, Q.Y. The antiviral defense mechanisms in mandarin fish induced by DNA vaccination against a rhabdovirus. Vet. Microbiol. 2012, 157, 264–275. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Size (bp) | Reference | Usage |
|---|---|---|---|---|
| pYD1 | F-AGTAACGTTTGTCAGTAATTGC | 399 | Designed | PCR |
| R-GTCGATTTTGTTACATCTACAC | ||||
| OMCP | F-GGATCCAGTGTGGTGGAATTCATGAGTAGCGTAACAGGC | 1389 | Designed | PCR |
| R-GCCCTCTAGACTCGAGCGGCCGCCACAAAATAGGAAAGCCC | ||||
| ltb | F-GGATCCAGTGTGGTGGGTACCCCTCAGTCTATTACAGAGC | 306 | Designed | PCR |
| R-TGAATTCCACCACACTGGATCCGTTTTCCATACTGATTGCCG | ||||
| TNF-α | F-ACTTCGTCTACAGCCAGGCA | 105 | [18] | qRT-PCR |
| R-AGTAACGCGAGACCCTGTGG | ||||
| IL-1β | F-TGGTGGAAAACAGCATGGAGC | 95 | [18] | qRT-PCR |
| R-AGGGTGCACGTAGTTCGACA | ||||
| IgM | F-GACTGGAGTGGCGGAAAGTGGAGG | 133 | [31] | qRT-PCR |
| R-TTTCATCTTCTACAAACGCAGACAACGG | ||||
| IgT | F-GAAGGTCAACAACGCTGAGTG | 248 | [32] | qRT-PCR |
| R-TGTTGCTGGTCACATCTAGTCC | ||||
| β-actin | F-CAGGATGCAGAAGGAGATCACA | 151 | [18] | qRT-PCR |
| R-CTCCTGCTTGCTGATCCACAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Chen, X.; Xue, M.; Jiang, N.; Li, Y.; Fan, Y.; Zhang, P.; Liu, N.; Xiao, Z.; Zhang, Q.; et al. Oral Vaccination of Largemouth Bass (Micropterus salmoides) against Largemouth Bass Ranavirus (LMBV) Using Yeast Surface Display Technology. Animals 2023, 13, 1183. https://doi.org/10.3390/ani13071183
Zhang M, Chen X, Xue M, Jiang N, Li Y, Fan Y, Zhang P, Liu N, Xiao Z, Zhang Q, et al. Oral Vaccination of Largemouth Bass (Micropterus salmoides) against Largemouth Bass Ranavirus (LMBV) Using Yeast Surface Display Technology. Animals. 2023; 13(7):1183. https://doi.org/10.3390/ani13071183
Chicago/Turabian StyleZhang, Mengjie, Xiaoyu Chen, Mingyang Xue, Nan Jiang, Yiqun Li, Yuding Fan, Peng Zhang, Naicheng Liu, Zidong Xiao, Qinghua Zhang, and et al. 2023. "Oral Vaccination of Largemouth Bass (Micropterus salmoides) against Largemouth Bass Ranavirus (LMBV) Using Yeast Surface Display Technology" Animals 13, no. 7: 1183. https://doi.org/10.3390/ani13071183