
The Mechanisms of BDNF Promoting the Proliferation of Porcine Follicular Granulosa Cells: Role of miR-127 and Involvement of the MAPK-ERK1/2 Pathway
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Isolation and Culture
2.2. Immunofluorescence Staining
2.3. Cell Treatment
2.4. Cell Transfection
2.5. Cell Viability Assay
2.6. RNA Extraction and Reverse Transcripton-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.7. Western Blotting
2.8. Flow Cytometric Analysis (FACS)
2.9. Sequencing of miRNAs
2.10. Bioinformatics Analysis
2.11. Data Analysis
3. Results
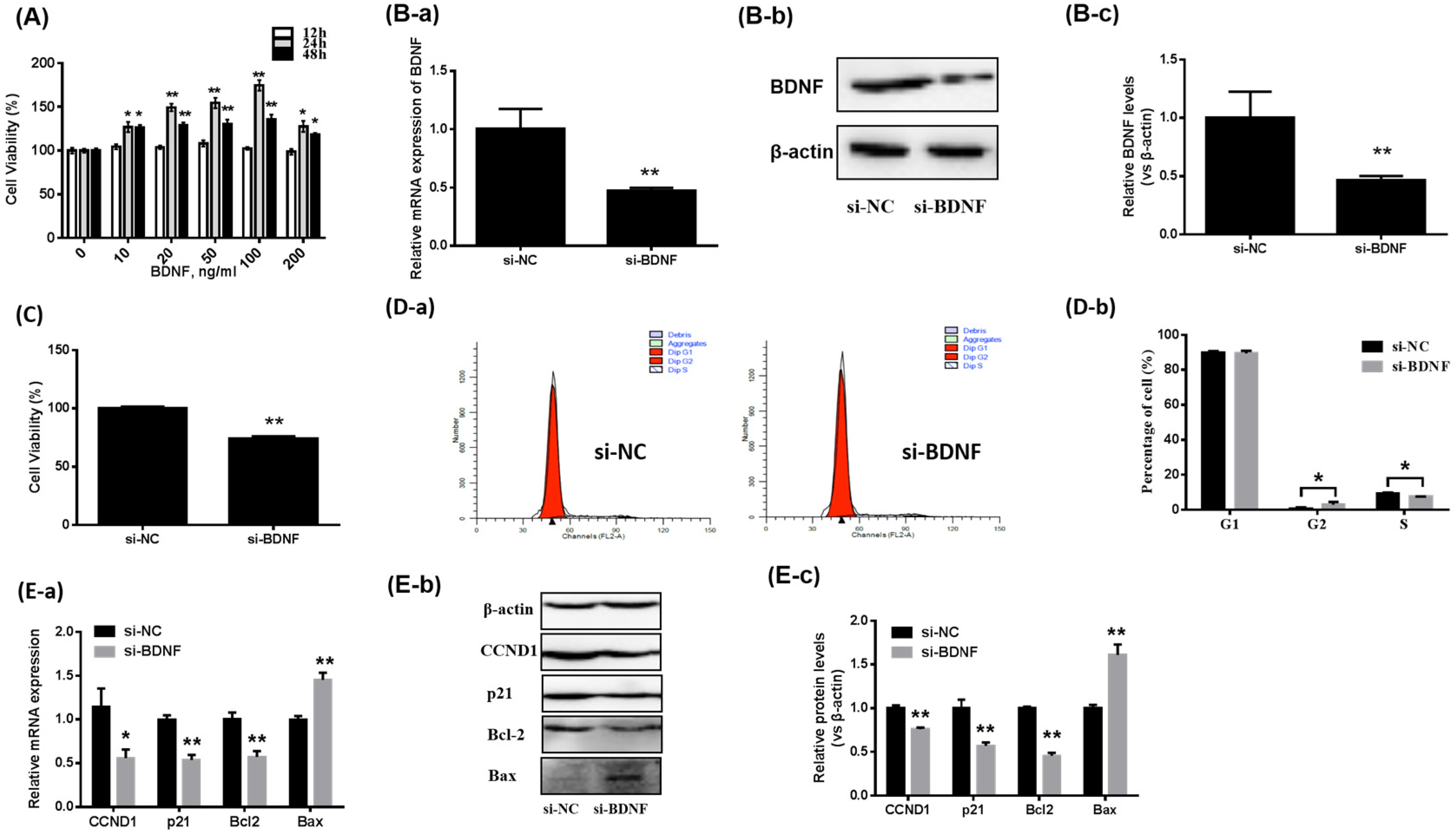
3.1. Effects of BDNF on the Proliferation of Porcine GCs
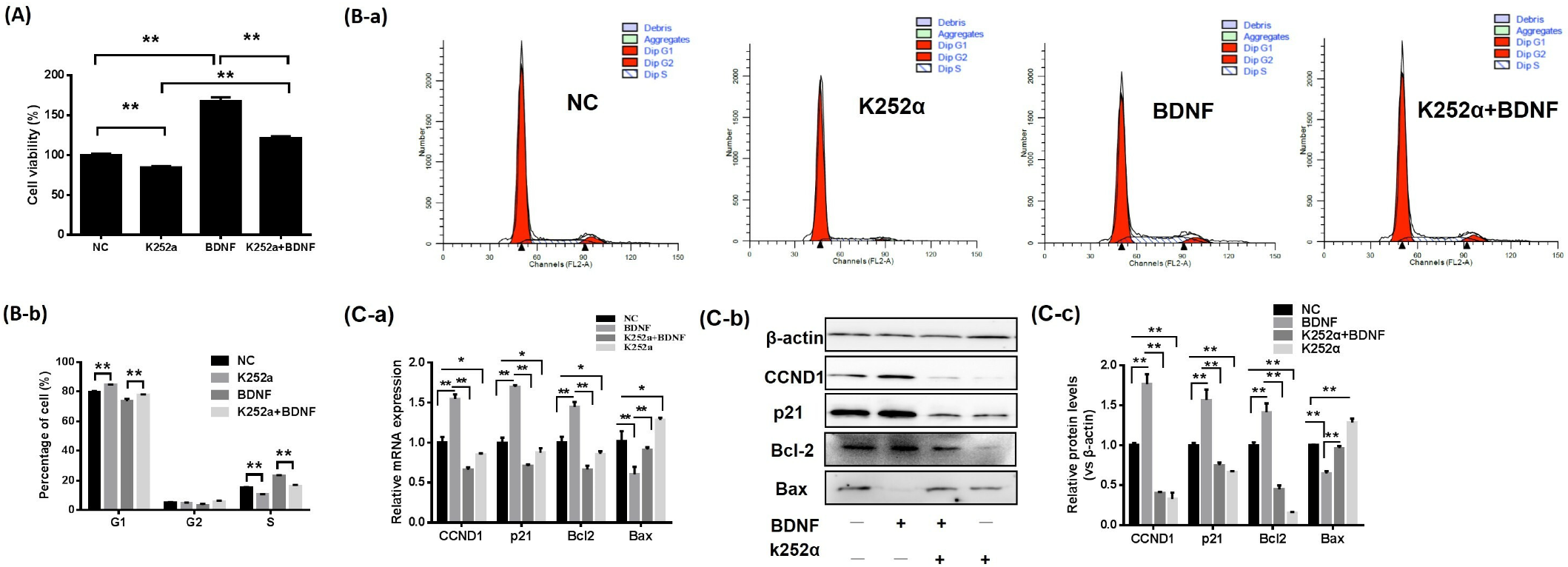
3.2. The BDNF/TrkB Pathway Affects the Proliferation of Porcine GCs
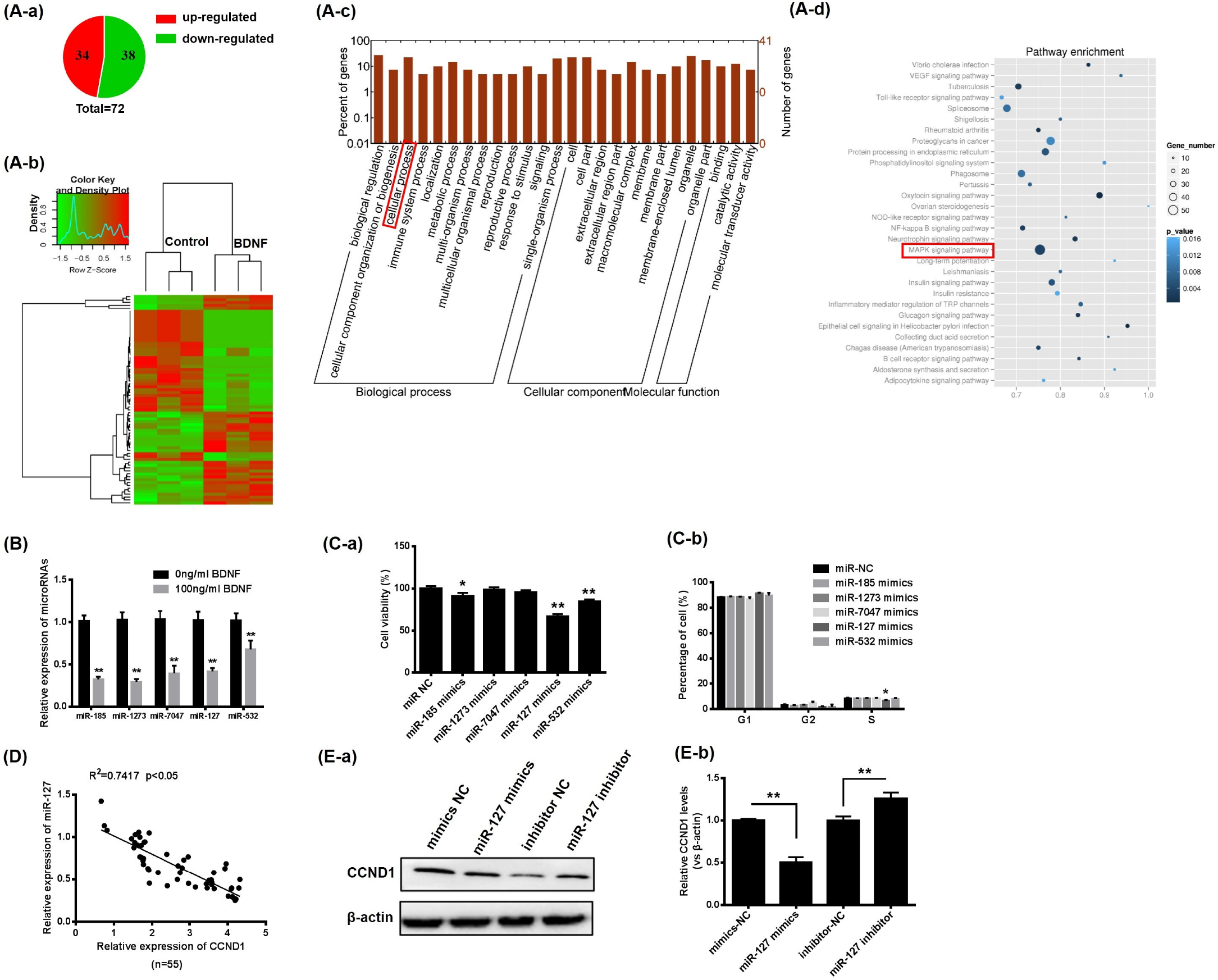
3.3. BDNF Promotes GCs Proliferation through Increase of CCND1 by Downregulating miR-127
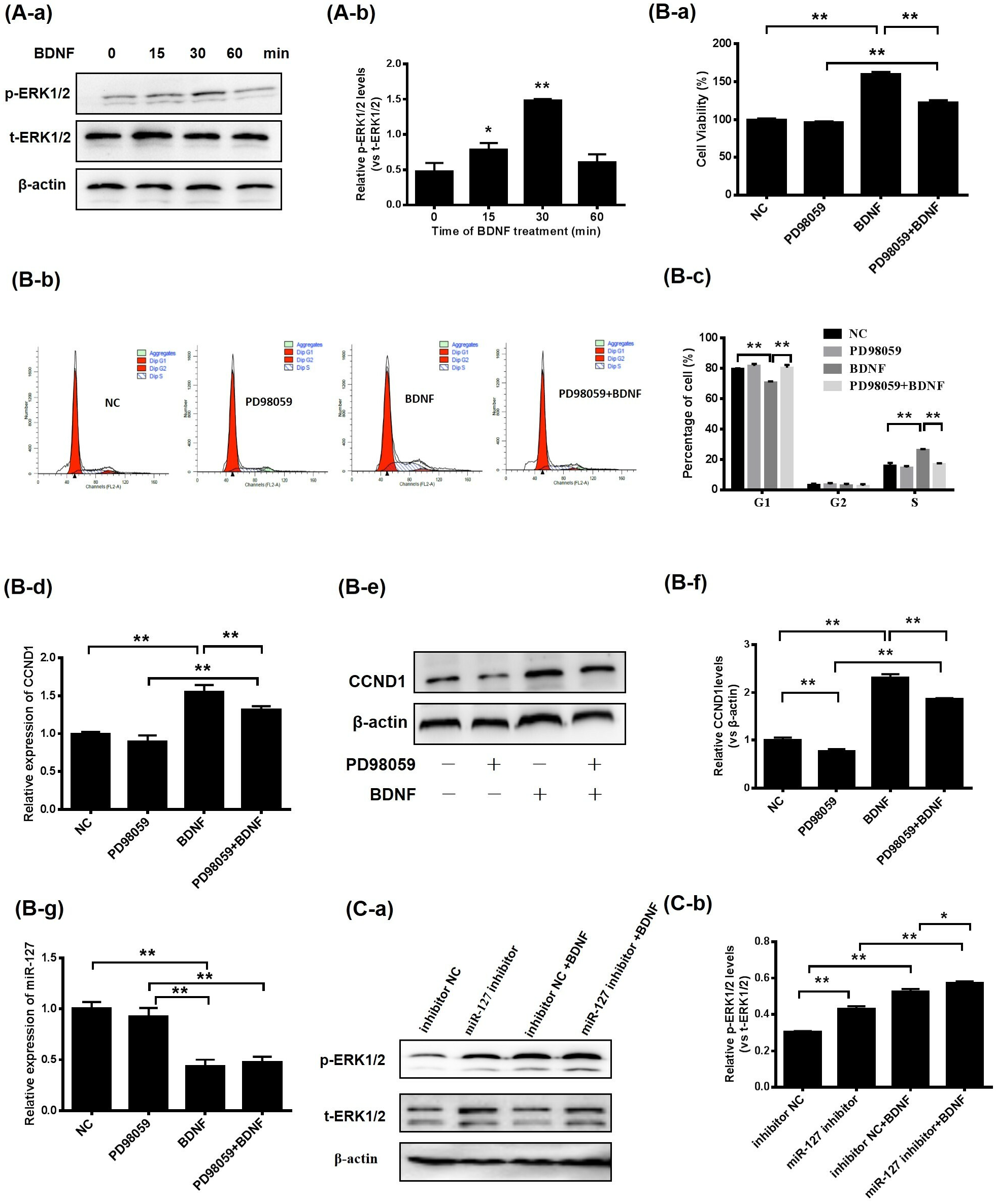
3.4. BDNF Promotes GC Proliferation via the ERK1/2 Signaling Pathway Mediated by miR-127
4. Discussion
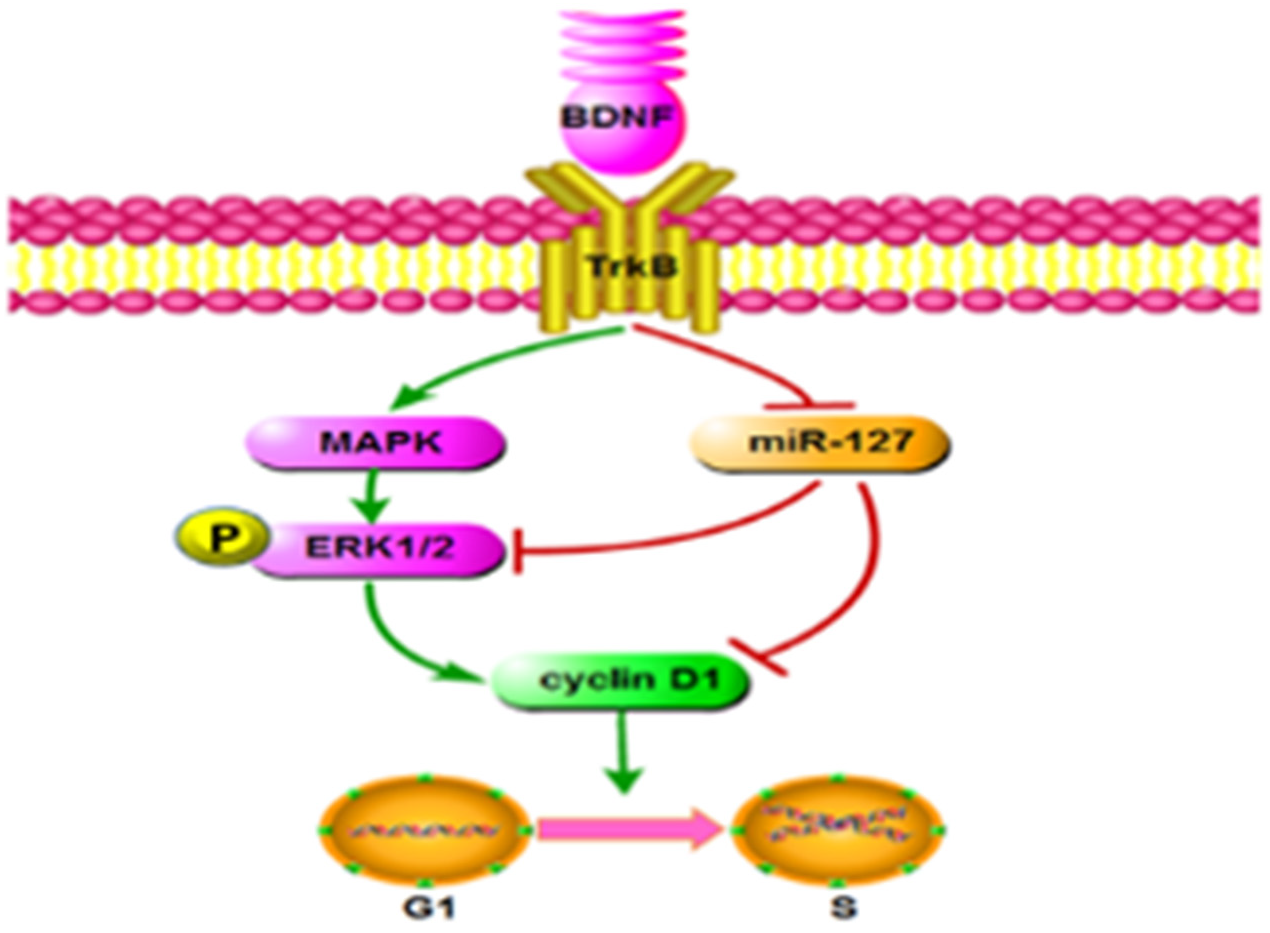
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pepling, M.E. Follicular assembly: Mechanisms of action. Reproduction 2012, 143, 139–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itami, S.; Yasuda, K.; Yoshida, Y.; Matsui, C.; Hashiura, S.; Sakai, A.; Tamotsu, S. Co-culturing of follicles with interstitial cells in collagen gel reproduce follicular development accompanied with theca cell layer formation. Reprod. Biol. Endocrinol. 2011, 9, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Martinowich, K.; Lee, F.S. BDNF at the synapse: Why location matters. Mol. Psychiatry 2017, 22, 1370–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohira, K.; Hayashi, M. A new aspect of the TrkB signaling pathway in neural plasticity. Curr. Neuropharmacol. 2009, 7, 276–285. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Nagappan, G.; Guan, X.; Nathan, P.J.; Wren, P. BDNF-based synaptic repair as a disease-modifying strategy for neurodegenerative diseases. Nat. Rev. Neurosci. 2013, 14, 401–416. [Google Scholar] [CrossRef]
- Dissen, G.A.; Romero, C.; Paredes, A.; Ojeda, S.R. Neurotrophic control of ovarian development. Microsc. Res. Tech. 2002, 59, 509–515. [Google Scholar] [CrossRef]
- Dissen, G.A.; Garcia-Rudaz, C.; Ojeda, S.R. Role of neurotrophic factors in early ovarian development. Semin. Reprod. Med. 2009, 27, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Levanti, M.B.; Germana, A.; Abbate, F.; Montalbano, G.; Vega, J.A.; Germana, G. TrkA and p75NTR in the ovary of adult cow and pig. J. Anat. 2005, 207, 93–96. [Google Scholar] [CrossRef]
- Jana, B.; Koszykowska, M.; Czarzasta, J. Expression of nerve growth factor and its receptors, TrkA and p75, in porcine ovaries. J. Reprod. Dev. 2011, 57, 468–474. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Jeong, Y.I.; Park, S.M.; Lee, J.Y.; Kim, J.H.; Park, S.W.; Hossein, M.S.; Jeong, Y.W.; Kim, S.; Hyun, S.H.; et al. Beneficial effects of brain-derived neurotropic factor on in vitro maturation of porcine oocytes. Reproduction 2007, 134, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Chen, C.; Chen, L.; Chen, S.; Li, H.; Zhao, Y.; Rao, J.; Zhou, X. BDNF-induced expansion of cumulus-oocyte complexes in pigs was mediated by microRNA-205. Theriogenology 2016, 85, 1476–1482. [Google Scholar] [CrossRef]
- Chow, R.; Wessels, J.M.; Foster, W.G. Brain-derived neurotrophic factor (BDNF) expression and function in the mammalian reproductive Tract. Hum. Reprod. Update 2020, 26, 545–564. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yi, K.; Sun, Y.; Sun, Y.; Tang, L.; Zhou, X. Expression of BDNF mRNA in Porcine Reproductive Tissues During Follicular Phase and Luteal Phase and Oocytes in GV and in vitro Matured Mil Stage. J. Anim. Veterniary Adv. 2011, 10, 2571–2574. [Google Scholar]
- Yi, K.L.; Zhou, X.; Shi, D.S.; Chen, H.H.; Qin, Q.L.; Chen, Y.; Li, C.J.; Zhao, Z.H.; Xing, S.Y. The mRNA expression of brain-derived neurotrophic factor in oocytes and embryos and its effects on the development of early embryos in cattle. Animal 2008, 2, 1786–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wang, F.; Liu, Z.; Zhao, Y.; Jiang, Y.; Chen, L.; Li, C.; Zhou, X. Brain-derived neurotrophic factor promotes proliferation and progesterone synthesis in bovine granulosa cells. J. Cell. Physiol. 2019, 234, 8776–8787. [Google Scholar] [CrossRef] [PubMed]
- Harel, S.; Jin, S.; Fisch, B.; Feldberg, D.; Krissi, H.; Felz, C.; Freimann, S.; Tan, S.L.; Ao, A.; Abir, R. Tyrosine kinase B receptor and its activated neurotrophins in ovaries from human fetuses and adults. Mol. Hum. Reprod. 2006, 12, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, S.R.; Romero, C.; Tapia, V.; Dissen, G.A. Neurotrophic and cell-cell dependent control of early follicular development. Mol. Cell. Endocrinol. 2000, 163, 67–71. [Google Scholar] [CrossRef]
- Spears, N.; Molinek, M.D.; Robinson, L.L.; Fulton, N.; Cameron, H.; Shimoda, K.; Telfer, E.E.; Anderson, R.A.; Price, D.J. The role of neurotrophin receptors in female germ-cell survival in mouse and human. Development 2003, 130, 5481–5491. [Google Scholar] [CrossRef] [Green Version]
- Paredes, A.; Romero, C.; Dissen, G.A.; DeChiara, T.M.; Reichardt, L.; Cornea, A.; Ojeda, S.R.; Xu, B. TrkB receptors are required for follicular growth and oocyte survival in the mammalian ovary. Dev. Biol. 2004, 267, 430–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, W.H. Regulation of mammalian spermatogenesis by miRNAs. Semin. Cell Dev. Biol. 2022, 121, 24–31. [Google Scholar] [CrossRef]
- Liang, G.; Wang, Q.; Zhang, G.; Li, Z.; Wang, Q. Differentially expressed miRNAs and potential therapeutic targets for asthenospermia. Andrologia 2022, 54, e14265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ji, X.; Zhou, D.; Li, Y.; Lin, J.; Liu, J.; Luo, H.; Cui, S. miR-143 is critical for the formation of primordial follicles in mice. Front. Biosci. 2013, 18, 588–597. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Du, X.; Zhou, J.; Pan, Z.; Liu, H.; Li, Q. MicroRNA-26b functions as a proapoptotic factor in porcine follicular Granulosa cells by targeting Sma-and Mad-related protein 4. Biol. Reprod. 2014, 91, 146. [Google Scholar] [CrossRef]
- Zhang, L.; Mao, B.; Zhao, X.; Yuan, Y.; Wang, W.; Lin, S. Translation regulatory long non-coding RNA 1 (TRERNA1) sponges microRNA-23a to suppress granulosa cell apoptosis in premature ovarian failure. Bioengineered 2022, 13, 2173–2180. [Google Scholar] [CrossRef] [PubMed]
- Vendrell-Flotats, M.; Garcia-Martinez, T.; Martinez-Rodero, I.; Lopez-Bejar, M.; LaMarre, J.; Yeste, M.; Mogas, T. In vitro maturation in the presence of Leukemia Inhibitory Factor modulates gene and miRNA expression in bovine oocytes and embryos. Sci. Rep. 2020, 10, 17777. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Salilew-Wondim, D.; Schellander, K.; Tesfaye, D. The role of microRNAs in mammalian oocytes and embryos. Anim. Reprod. Sci. 2012, 134, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Lyu, X.; Ding, L.; Ke, L.; Yang, D.; Pirouz, M.; Qi, Y.; Ong, J.; Gao, G.; Du, P.; et al. Global miRNA dosage control of embryonic germ layer specification. Nature 2021, 593, 602–606. [Google Scholar] [CrossRef] [PubMed]
- Keifer, J.; Zheng, Z.; Ambigapathy, G. A MicroRNA-BDNF Negative Feedback Signaling Loop in Brain: Implications for Alzheimer’s Disease. Microrna 2015, 4, 101–108. [Google Scholar] [CrossRef]
- Caputo, V.; Sinibaldi, L.; Fiorentino, A.; Parisi, C.; Catalanotto, C.; Pasini, A.; Cogoni, C.; Pizzuti, A. Brain derived neurotrophic factor (BDNF) expression is regulated by microRNAs miR-26a and miR-26b allele-specific binding. PLoS ONE 2011, 6, e28656. [Google Scholar] [CrossRef] [Green Version]
- Maurel, O.M.; Torrisi, S.A.; Barbagallo, C.; Purrello, M.; Salomone, S.; Drago, F.; Ragusa, M.; Leggio, G.M. Dysregulation of miR-15a-5p, miR-497a-5p and miR-511-5p Is Associated with Modulation of BDNF and FKBP5 in Brain Areas of PTSD-Related Susceptible and Resilient Mice. Int. J. Mol. Sci. 2021, 22, 5157. [Google Scholar] [CrossRef]
- Shang, C.; Chen, Q.; Zu, F.; Ren, W. Integrated analysis identified prognostic microRNAs in breast cancer. BMC Cancer 2022, 22, 1170. [Google Scholar] [CrossRef]
- Chang, S.; Sun, L.; Feng, G. SP1-mediated long noncoding RNA POU3F3 accelerates the cervical cancer through miR-127-5p/FOXD1. Biomed. Pharm. 2019, 117, 109133. [Google Scholar] [CrossRef]
- Simpson, L.O. Renal function and kidney morphology in biopsies from patients with IgA nephropathy. Clin. Nephrol. 1993, 40, 122–124. [Google Scholar]
- Heavner, J.E.; de Jong, R.H. Lidocaine blocking concentrations for B- and C-nerve fibers. Anesthesiology 1974, 40, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Du, S.Y.; Huang, X.X.; Li, N.M.; Lv, C.Y.; Lv, C.H.; Wei, M.L.; Gao, Z.; Zhang, Y.P. MiR-127-3p inhibits proliferation of ovarian cancer in rats through down-regulating MAPK4. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10383–10390. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.; You, S.; Cao, Y.; Li, C.; Wei, J.; Wang, D.; Qiao, J.; Zhao, X.; Hu, S.; Quan, R. Aberrant expression of miR-127, miR-21 and miR-16 in placentas of deceased cloned sheep. Res. Vet. Sci. 2016, 105, 200–204. [Google Scholar] [CrossRef]
- Ito, M.; Sferruzzi-Perri, A.N.; Edwards, C.A.; Adalsteinsson, B.T.; Allen, S.E.; Loo, T.H.; Kitazawa, M.; Kaneko-Ishino, T.; Ishino, F.; Stewart, C.L.; et al. A trans-homologue interaction between reciprocally imprinted miR-127 and Rtl1 regulates placenta development. Development 2015, 142, 2425–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.S.; Zhang, D.X.; Ko, Y.G.; Kim, N.H. Aberrant epigenetic reprogramming of imprinted microRNA-127 and Rtl1 in cloned mouse embryos. Biochem. Biophys. Res. Commun. 2009, 379, 390–394. [Google Scholar] [CrossRef]
- Butler, A.E.; Ramachandran, V.; Hayat, S.; Dargham, S.R.; Cunningham, T.K.; Benurwar, M.; Sathyapalan, T.; Najafi-Shoushtari, S.H.; Atkin, S.L. Expression of microRNA in follicular fluid in women with and without PCOS. Sci. Rep. 2019, 9, 16306. [Google Scholar] [CrossRef] [Green Version]
- Nothnick, W.B. The role of micro-RNAs in the female reproductive tract. Reproduction 2012, 143, 559–576. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.Y.; An, X.P.; Fang, F.; Gao, K.X.; Xin, H.Y.; Han, P.; Bao, L.J.; Ma, H.D.; Cao, B.Y. MicroRNA-10b suppresses goat granulosa cell proliferation by targeting brain-derived neurotropic factor. Domest. Anim. Endocrinol. 2016, 54, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Yang, J.; Bai, S.; Wei, S. MiR-15a inhibits the levels of porcine ovarian granulosa cell’s BDNF. Anim. Husb. Vet. Med. 2018, 50, 14–19. [Google Scholar]
- Seifer, D.B.; Lambert-Messerlian, G.; Schneyer, A.L. Ovarian brain-derived neurotrophic factor is present in follicular fluid from normally cycling women. Fertil. Steril. 2003, 79, 451–452. [Google Scholar] [CrossRef]
- Wang, X.; Sun, Z.; Zhen, J.; Yu, Q. Brain-derived neurotrophic factor from follicular fluid is positively associated with rate of mature ooocytes collected and cleavage rate in intracytoplasmic sperm injection patients. J. Assist. Reprod. Genet. 2011, 28, 1053–1058. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Eldredge, J.B.; Xu, Y.; Puett, D. MicroRNA expression and regulation in human ovarian carcinoma cells by luteinizing hormone. PLoS ONE 2011, 6, e21730. [Google Scholar] [CrossRef] [Green Version]
- Donadeu, F.X.; Schauer, S.N.; Sontakke, S.D. Involvement of miRNAs in ovarian follicular and luteal development. J. Endocrinol. 2012, 215, 323–334. [Google Scholar] [CrossRef]
- Pan, B.; Toms, D.; Shen, W.; Li, J. MicroRNA-378 regulates oocyte maturation via the suppression of aromatase in porcine cumulus cells. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E525–E534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wang, M.; Guo, M.; Xie, Y.; Cong, Y.S. miR-127 regulates cell proliferation and senescence by targeting BCL6. PLoS ONE 2013, 8, e80266. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Lin, Y. Hsa-mir-127 impairs survival of patients with glioma and promotes proliferation, migration and invasion of cancerous cells by modulating replication initiator 1. Neuroreport 2018, 29, 1166–1173. [Google Scholar] [CrossRef]
- Wang, Y.; Kong, D. Knockdown of lncRNA MEG3 inhibits viability, migration, and invasion and promotes apoptosis by sponging miR-127 in osteosarcoma cell. J. Cell. Biochem. 2018, 119, 669–679. [Google Scholar] [CrossRef]
- Ma, H.; Lin, Y.; Zhao, Z.A.; Lu, X.; Yu, Y.; Zhang, X.; Wang, Q.; Li, L. MicroRNA-127 Promotes Mesendoderm Differentiation of Mouse Embryonic Stem Cells by Targeting Left-Right Determination Factor 2. J. Biol. Chem. 2016, 291, 12126–12135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohoutek, J.; Dvorak, P.; Hampl, A. Temporal distribution of CDK4, CDK6, D-type cyclins, and p27 in developing mouse oocytes. Biol. Reprod. 2004, 70, 139–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hume, A.J.; Finkel, J.S.; Kamil, J.P.; Coen, D.M.; Culbertson, M.R.; Kalejta, R.F. Phosphorylation of retinoblastoma protein by viral protein with cyclin-dependent kinase function. Science 2008, 320, 797–799. [Google Scholar] [CrossRef] [PubMed]
- Sherr, C.J.; Roberts, J.M. CDK inhibitors: Positive and negative regulators of G1-phase progression. Genes Dev. 1999, 13, 1501–1512. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, R.L.; Musgrove, E.A. CDK inhibitors as potential breast cancer therapeutics: New evidence for enhanced efficacy in ER+ disease. Breast Cancer Res. 2009, 11, 112. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Wu, W.; Ge, H.; Li, P.; Wang, Z. Up-Regulation of miR-204 Enhances Anoikis Sensitivity in Epithelial Ovarian Cancer Cell Line Via Brain-Derived Neurotrophic Factor Pathway In Vitro. Int. J. Gynecol. Cancer 2015, 25, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Woitach, J.T.; Zhang, M.; Niu, C.H.; Thorgeirsson, S.S. A retinoblastoma-binding protein that affects cell-cycle control and confers transforming ability. Nat. Genet. 1998, 19, 371–374. [Google Scholar] [CrossRef]
- Hydbring, P.; Malumbres, M.; Sicinski, P. Non-canonical functions of cell cycle cyclins and cyclin-dependent kinases. Nat. Rev. Mol. Cell Biol. 2016, 17, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Filippi, S.; Barnes, C.P.; Kirk, P.D.; Kudo, T.; Kunida, K.; McMahon, S.S.; Tsuchiya, T.; Wada, T.; Kuroda, S.; Stumpf, M.P. Robustness of MEK-ERK Dynamics and Origins of Cell-to-Cell Variability in MAPK Signaling. Cell Rep. 2016, 15, 2524–2535. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Ju, Q.; Zhu, J.; Jiang, Y.; Yang, X.; Liu, X.; Ma, J.; Sun, C.; Shi, J. RASSF9 promotes NSCLC cell proliferation by activating the MEK/ERK axis. Cell Death Discov. 2021, 7, 199. [Google Scholar] [CrossRef]
- Yuen, H.F.; Chan, K.K.; Grills, C.; Murray, J.T.; Platt-Higgins, A.; Eldin, O.S.; O′Byrne, K.; Janne, P.; Fennell, D.A.; Johnston, P.G.; et al. Ran is a potential therapeutic target for cancer cells with molecular changes associated with activation of the PI3K/Akt/mTORC1 and Ras/MEK/ERK pathways. Clin. Cancer Res. 2012, 18, 380–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, J.V.; Wurtzel, J.G.; Goldfinger, L.E. Regulation of H-Ras-driven MAPK signaling, transformation and tumorigenesis, but not PI3K signaling and tumor progression, by plasma membrane microdomains. Oncogenesis 2016, 5, e228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durruthy-Durruthy, J.; Sebastiano, V.; Wossidlo, M.; Cepeda, D.; Cui, J.; Grow, E.J.; Davila, J.; Mall, M.; Wong, W.H.; Wysocka, J.; et al. The primate-specific noncoding RNA HPAT5 regulates pluripotency during human preimplantation development and nuclear reprogramming. Nat. Genet. 2016, 48, 44–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, C.; Cawthorn, W.P.; Li, Y.; Zhao, G.; Macdougald, O.A.; Franceschi, R.T. Reciprocal Control of Osteogenic and Adipogenic Differentiation by ERK/MAP Kinase Phosphorylation of Runx2 and PPARgamma Transcription Factors. J. Cell. Physiol. 2016, 231, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Li, Y.; Li, Z.; Liu, C.; Xue, P. Sal B targets TAZ to facilitate osteogenesis and reduce adipogenesis through MEK-ERK pathway. J. Cell. Mol. Med. 2019, 23, 3683–3695. [Google Scholar] [CrossRef]
- Zhao, H.; Bo, Q.; Wu, Z.; Liu, Q.; Li, Y.; Zhang, N.; Guo, H.; Shi, B. KIF15 promotes bladder cancer proliferation via the MEK-ERK signaling pathway. Cancer Manag. Res. 2019, 11, 1857–1868. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Gao, Z.L.; Zhang, Z.P.; Jiang, H.B.; Yang, C.Q.; Yang, J.; Xia, X.B. Downregulation of CKS1B restrains the proliferation, migration, invasion and angiogenesis of retinoblastoma cells through the MEK/ERK signaling pathway. Int. J. Mol. Med. 2019, 44, 103–114. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.; Chen, L.; Chen, T.; Cao, M.; Zhang, B.; Yuan, C.; Zhao, Z.; Li, C.; Zhou, X. The Mechanisms of BDNF Promoting the Proliferation of Porcine Follicular Granulosa Cells: Role of miR-127 and Involvement of the MAPK-ERK1/2 Pathway. Animals 2023, 13, 1115. https://doi.org/10.3390/ani13061115
Zheng X, Chen L, Chen T, Cao M, Zhang B, Yuan C, Zhao Z, Li C, Zhou X. The Mechanisms of BDNF Promoting the Proliferation of Porcine Follicular Granulosa Cells: Role of miR-127 and Involvement of the MAPK-ERK1/2 Pathway. Animals. 2023; 13(6):1115. https://doi.org/10.3390/ani13061115
Chicago/Turabian StyleZheng, Xue, Lu Chen, Tong Chen, Maosheng Cao, Boqi Zhang, Chenfeng Yuan, Zijiao Zhao, Chunjin Li, and Xu Zhou. 2023. "The Mechanisms of BDNF Promoting the Proliferation of Porcine Follicular Granulosa Cells: Role of miR-127 and Involvement of the MAPK-ERK1/2 Pathway" Animals 13, no. 6: 1115. https://doi.org/10.3390/ani13061115