

In Vivo and In Vitro Expression of iC1, a Methylation-Controlled J Protein (MCJ) in Bovine Liver, and Response to In Vitro Bovine Fatty Liver Disease Model
 ,
,
Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
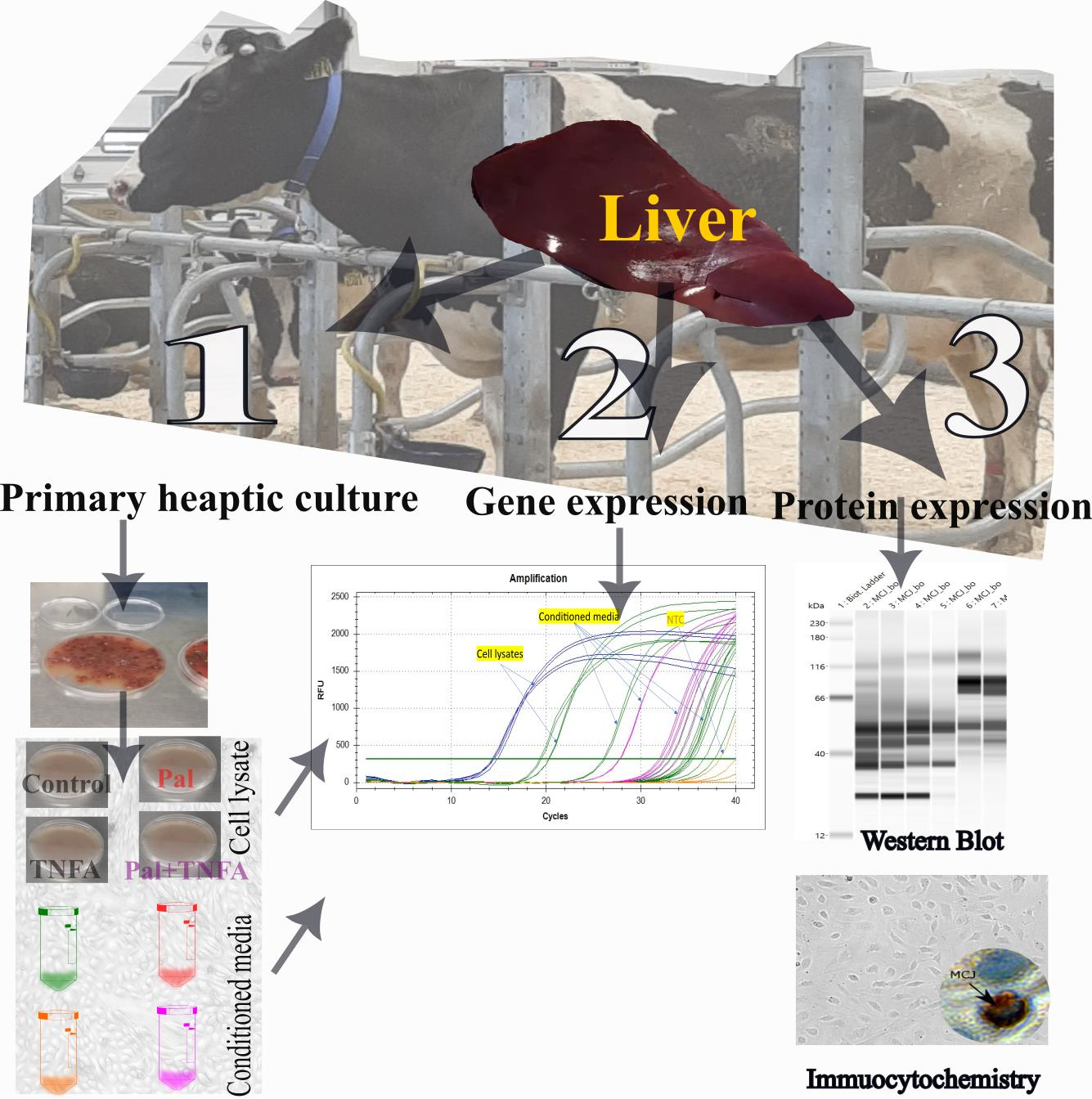
2. Materials and Methods
2.1. Isolation of Primary Hepatocytes of Bovine
2.2. Preparation of TNFA and Palmitic Acid Solutions
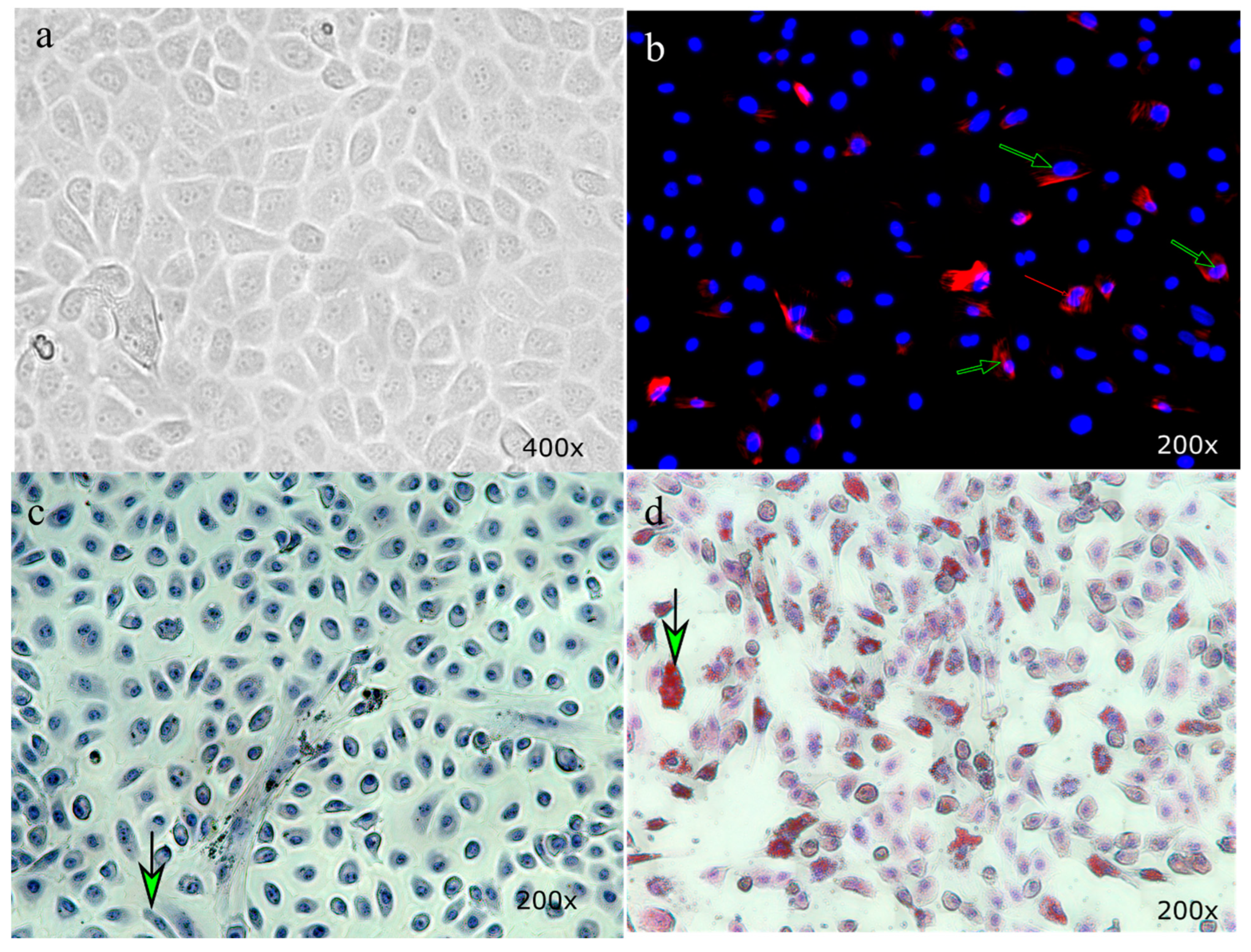
2.3. Oil Red O Staining of Hepatic cells
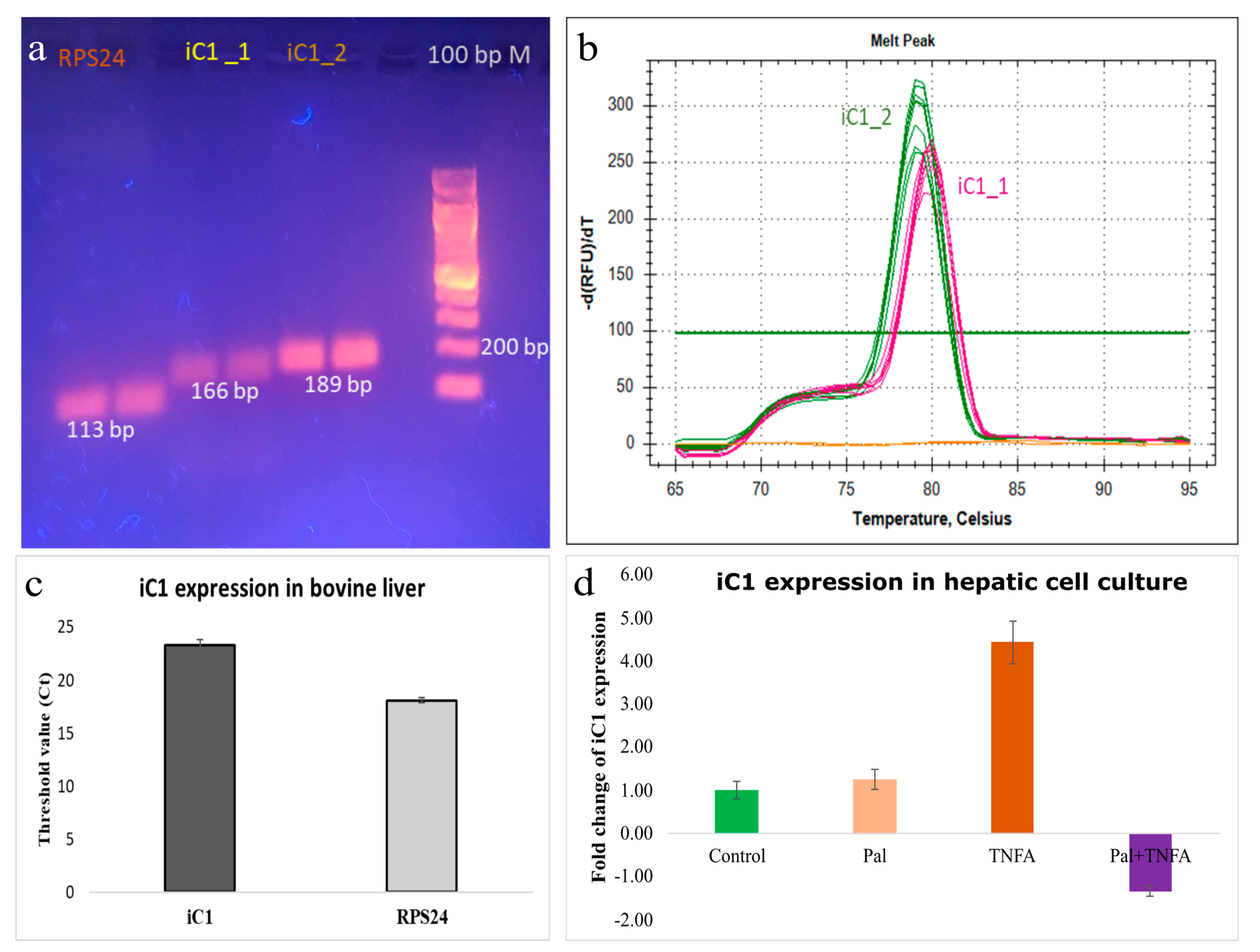
2.4. Gene Expression
2.5. Protein Isolation and Western Blot
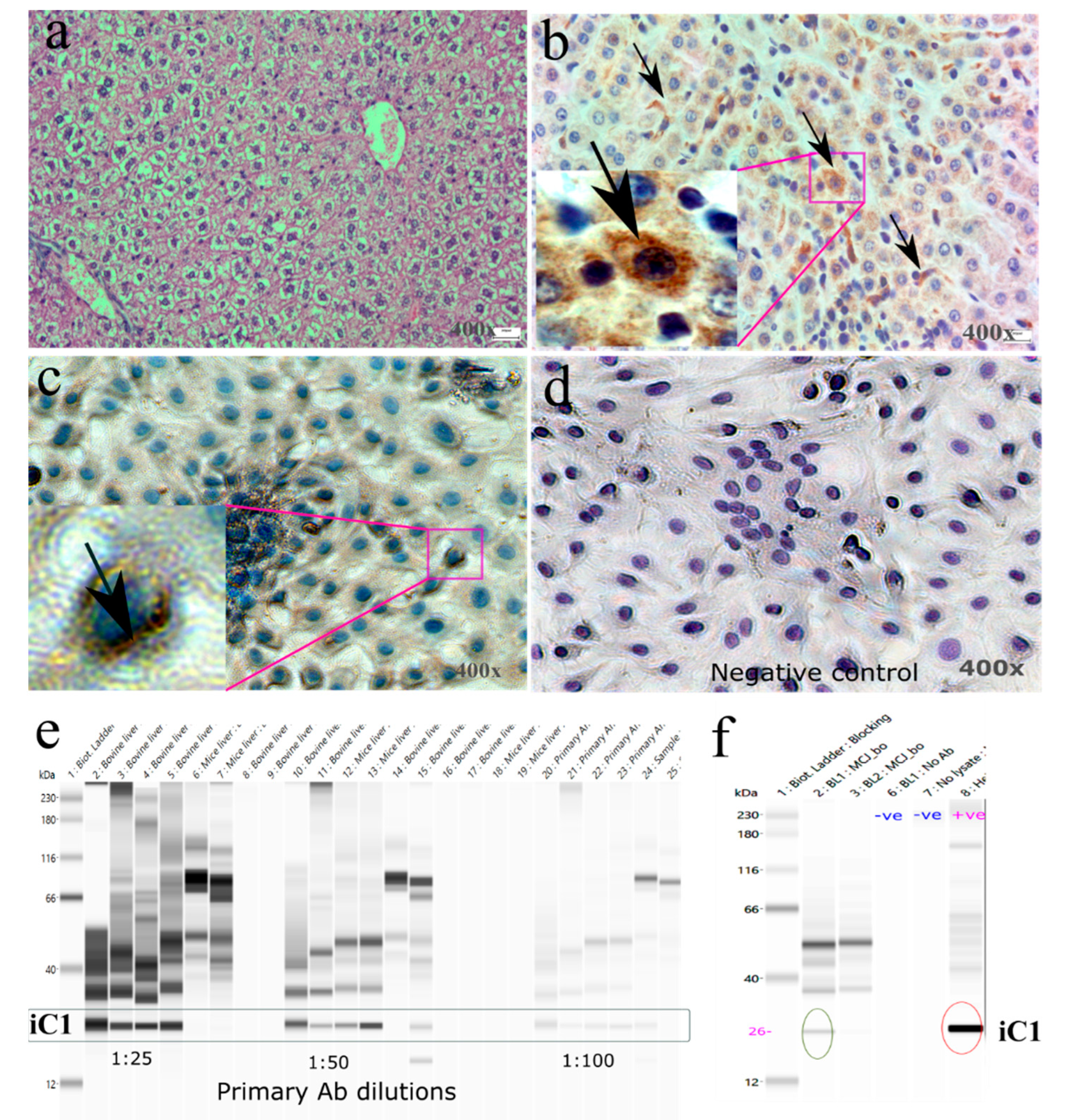
2.6. Immunostaining
3. Results and Discussion
3.1. Primary Hepatocyte Culture of Bovine
3.2. Gene Expression of iC1 in Bovine Liver
3.3. Protein Expression of iC1 in Bovine Liver
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Trevisi, E.; Amadori, M.; Cogrossi, S.; Razzuoli, E.; Bertoni, G. Metabolic Stress and Inflammatory Response in High-Yielding, Periparturient Dairy Cows. Res. Vet. Sci. 2012, 93, 695–704. [Google Scholar] [CrossRef]
- Du, X.; Shen, T.; Wang, H.; Qin, X.; Xing, D.; Ye, Q.; Shi, Z.; Fang, Z.; Zhu, Y.; Yang, Y.; et al. Adaptations of Hepatic Lipid Metabolism and Mitochondria in Dairy Cows with Mild Fatty Liver. J. Dairy Sci. 2018, 101, 9544–9558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobe, G.; Young, J.W.; Beitz, D.C. Invited Review: Pathology, Etiology, Prevention, and Treatment of Fatty Liver in Dairy Cows. J. Dairy Sci. 2004, 87, 3105–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezzetti, M.; Cattaneo, L.; Passamonti, M.M.; Lopreiato, V.; Minuti, A.; Trevisi, E.; Dänicke, S. The Transition Period Updated: A Review of the New Insights into the Adaptation of Dairy Cows to the New Lactation. Dairy 2021, 2, 617–636. [Google Scholar] [CrossRef]
- Overton, T.R.; Waldron, M.R. Nutritional Management of Transition Dairy Cows: Strategies to Optimize Metabolic Health. J. Dairy Sci. 2004, 87, E105–E119. [Google Scholar] [CrossRef] [Green Version]
- Buonaiuto, G.; Cavallini, D.; Mammi, L.M.E.; Ghiaccio, F.; Palmonari, A.; Formigoni, A.; Visentin, G. The Accuracy of NIRS in Predicting Chemical Composition and Fibre Digestibility of Hay-Based Total Mixed Rations. Ital. J. Anim. Sci. 2021, 20, 1730–1739. [Google Scholar] [CrossRef]
- Ingvartsen, K.L. Feeding- and Management-Related Diseases in the Transition Cow: Physiological Adaptations around Calving and Strategies to Reduce Feeding-Related Diseases. Anim. Feed Sci. Technol. 2006, 126, 175–213. [Google Scholar] [CrossRef]
- Ferguson, J.D.; Sklan, D.; Chalupa, W.V.; Kronfeld, D.S. Effects of Hard Fats on In Vitro and In Vivo Rumen Fermentation Milk Production, and Reproduction in Dairy Cows. J. Dairy Sci. 1990, 73, 2864–2879. [Google Scholar] [CrossRef]
- Horst, E.A.; Kvidera, S.K.; Baumgard, L.H. Invited Review: The Influence of Immune Activation on Transition Cow Health and Performance-A Critical Evaluation of Traditional Dogmas. J. Dairy Sci. 2021, 104, 8380–8410. [Google Scholar] [CrossRef]
- Buonaiuto, G.; Lopez-Villalobos, N.; Niero, G.; Degano, L.; Dadati, E.; Formigoni, A.; Visentin, G. The Application of Legendre Polynomials to Model Muscularity and Body Condition Score in Primiparous Italian Simmental Cattle. Ital. J. Anim. Sci. 2022, 21, 350–360. [Google Scholar] [CrossRef]
- Shao, T.; McCann, J.C.; Shike, D.W. Effects of Late Gestation Supplements Differing in Fatty Acid Amount and Profile to Beef Cows on Cow Performance, Steer Progeny Growth Performance through Weaning, and Relative MRNA Expression of Genes Associated with Muscle and Adipose Tissue Development. Animals 2023, 13, 437. [Google Scholar] [CrossRef]
- Rukkwamsuk, T.; Geelen, M.J.H.; Kruip, T.A.M.; Wensing, T. Interrelation of Fatty Acid Composition in Adipose Tissue, Serum, and Liver of Dairy Cows During the Development of Fatty Liver Postpartum. J. Dairy Sci. 2000, 83, 52–59. [Google Scholar] [CrossRef]
- Malhi, H.; Bronk, S.F.; Werneburg, N.W.; Gores, G.J. Free Fatty Acids Induce JNK-Dependent Hepatocyte Lipoapoptosis. J. Biol. Chem. 2006, 281, 12093–12101. [Google Scholar] [CrossRef] [Green Version]
- Kakino, S.; Ohki, T.; Nakayama, H.; Yuan, X.; Otabe, S.; Hashinaga, T.; Wada, N.; Kurita, Y.; Tanaka, K.; Hara, K.; et al. Pivotal Role of TNF-α in the Development and Progression of Nonalcoholic Fatty Liver Disease in a Murine Model. Horm. Metab. Res. 2018, 50, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Hatle, K.M.; Gummadidala, P.; Navasa, N.; Bernardo, E.; Dodge, J.; Silverstrim, B.; Fortner, K.; Burg, E.; Suratt, B.T.; Hammer, J.; et al. MCJ/DnaJC15, an Endogenous Mitochondrial Repressor of the Respiratory Chain That Controls Metabolic Alterations. Mol. Cell Biol. 2013, 33, 2302–2314. [Google Scholar] [CrossRef] [Green Version]
- Barbier-Torres, L.; Iruzubieta, P.; Fernández-Ramos, D.; Delgado, T.C.; Taibo, D.; Guitiérrez-De-Juan, V.; Varela-Rey, M.; Azkargorta, M.; Navasa, N.; Fernández-Tussy, P.; et al. The Mitochondrial Negative Regulator MCJ Is a Therapeutic Target for Acetaminophen-Induced Liver Injury. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; LaCasse, M.; Beitz, D.C.; Testroet, E.D. Fatty Liver Disease and Utility of Stem Cells in Developing the Disease Model. In Stem Cells in Veterinary Science; Springer: Singapore, 2021; pp. 79–96. [Google Scholar] [CrossRef]
- Testroet, E.D.; Choudhary, S.; Choudhary, R.K.; Beitz, D.C.; Du, M. Tumor Necrosis Factor Alpha and Palmitate Simulate Bovine Fatty Liver Disease in Vitro When Using Abattoir-Derived Primary Bovine Hepatocytes Isolated by a Novel Nonperfusion Method. JDS Commun. 2022, 3, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Tapia, N.C.; Rosso, N.; Tiribelli, C. In Vitro Models for the Study of Non-Alcoholic Fatty Liver Disease. Curr. Med. Chem. 2011, 18, 1079–1084. [Google Scholar] [CrossRef]
- MacParland, S.A.; Ma, X.-Z.; Chen, L.; Khattar, R.; Cherepanov, V.; Selzner, M.; Feld, J.J.; Selzner, N.; McGilvray, I.D. Lipopolysaccharide and Tumor Necrosis Factor Alpha Inhibit Interferon Signaling in Hepatocytes by Increasing Ubiquitin-like Protease 18 (USP18) Expression. J. Virol. 2016, 90, 5549–5560. [Google Scholar] [CrossRef] [Green Version]
- Wobser, H.; Dorn, C.; Weiss, T.S.; Amann, T.; Bollheimer, C.; Büttner, R.; Schölmerich, J.; Hellerbrand, C. Lipid Accumulation in Hepatocytes Induces Fibrogenic Activation of Hepatic Stellate Cells. Cell Res. 2009, 19, 996–1005. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, R.K.; Choudhary, S.; Mukhopadhyay, C.S.; Pathak, D.; Verma, R. Deciphering the Transcriptome of Prepubertal Buffalo Mammary Glands Using RNA Sequencing. Funct. Integr. Genom. 2019, 19, 349–362. [Google Scholar] [CrossRef]
- Chu, C.M.; Shyu, W.C.; Liaw, Y.F. Comparative Studies on Expression of Alpha-Smooth Muscle Actin in Hepatic Stellate Cells in Chronic Hepatitis B and C. Dig. Dis. Sci. 2007, 53, 1364–1369. [Google Scholar] [CrossRef] [PubMed]
- Carpino, G.; Morini, S.; Ginanni Corradini, S.; Franchitto, A.; Merli, M.; Siciliano, M.; Gentili, F.; Onetti Muda, A.; Berloco, P.; Rossi, M.; et al. Alpha-SMA Expression in Hepatic Stellate Cells and Quantitative Analysis of Hepatic Fibrosis in Cirrhosis and in Recurrent Chronic Hepatitis after Liver Transplantation. Dig. Liver Dis. 2005, 37, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Wang, J.; Lv, Q.; Liu, M.; Xu, H.; Zhang, H.; Jin, L.; Yu, J.; Wang, X. Three-Dimensional Coculture of Primary Hepatocytes and Stellate Cells in Silk Scaffold Improves Hepatic Morphology and Functionality In Vitro. J. Biomed. Mater. Res. A 2018, 106, 2171–2180. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, M.R.; Vaandrager, A.B.; Helms, J.B. Some Lipid Droplets Are More Equal Than Others: Different Metabolic Lipid Droplet Pools in Hepatic Stellate Cells. Lipid Insights 2017, 10, 117863531774728. [Google Scholar] [CrossRef] [Green Version]
- Beekman, C.; Janson, A.A.; Baghat, A.; van Deutekom, J.C.; Datson, N.A. Use of Capillary Western Immunoassay (Wes) for Quantification of Dystrophin Levels in Skeletal Muscle of Healthy Controls and Individuals with Becker and Duchenne Muscular Dystrophy. PLoS ONE 2018, 13, e0195850. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choudhary, S.; LaCasse, M.; Choudhary, R.K.; Rincon, M.; Beitz, D.C.; Testroet, E.D. In Vivo and In Vitro Expression of iC1, a Methylation-Controlled J Protein (MCJ) in Bovine Liver, and Response to In Vitro Bovine Fatty Liver Disease Model. Animals 2023, 13, 1101. https://doi.org/10.3390/ani13061101
Choudhary S, LaCasse M, Choudhary RK, Rincon M, Beitz DC, Testroet ED. In Vivo and In Vitro Expression of iC1, a Methylation-Controlled J Protein (MCJ) in Bovine Liver, and Response to In Vitro Bovine Fatty Liver Disease Model. Animals. 2023; 13(6):1101. https://doi.org/10.3390/ani13061101
Chicago/Turabian StyleChoudhary, Shanti, Michelle LaCasse, Ratan Kumar Choudhary, Mercedes Rincon, Donald C. Beitz, and Eric D. Testroet. 2023. "In Vivo and In Vitro Expression of iC1, a Methylation-Controlled J Protein (MCJ) in Bovine Liver, and Response to In Vitro Bovine Fatty Liver Disease Model" Animals 13, no. 6: 1101. https://doi.org/10.3390/ani13061101