

Occurrence of Giardia duodenalis in Cats from Queretaro and the Risk to Public Health
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Setting and Sampling
2.2. Coproparasitoscopic Analysis
2.3. Sample Pooling
2.4. DNA Extraction
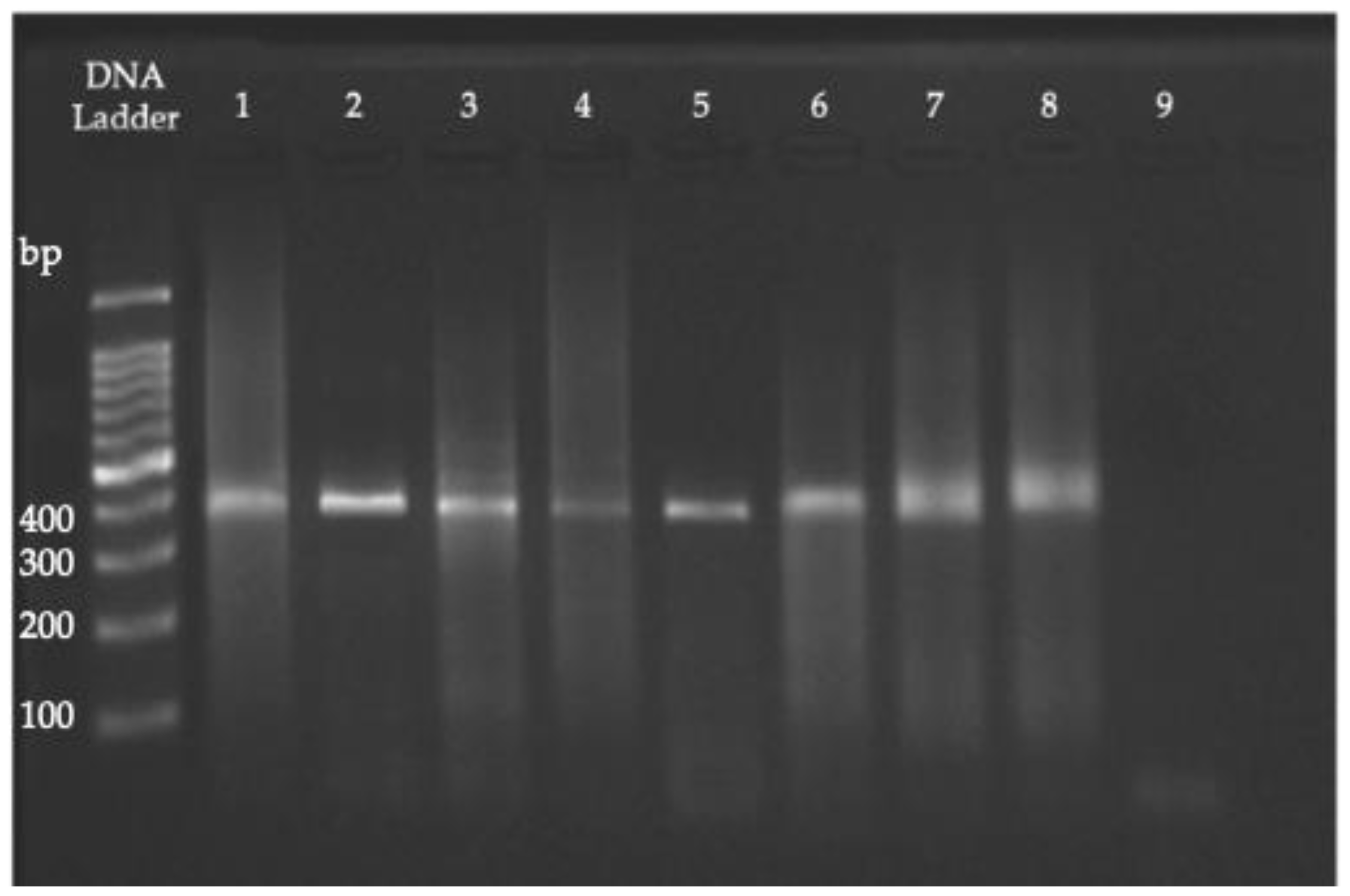
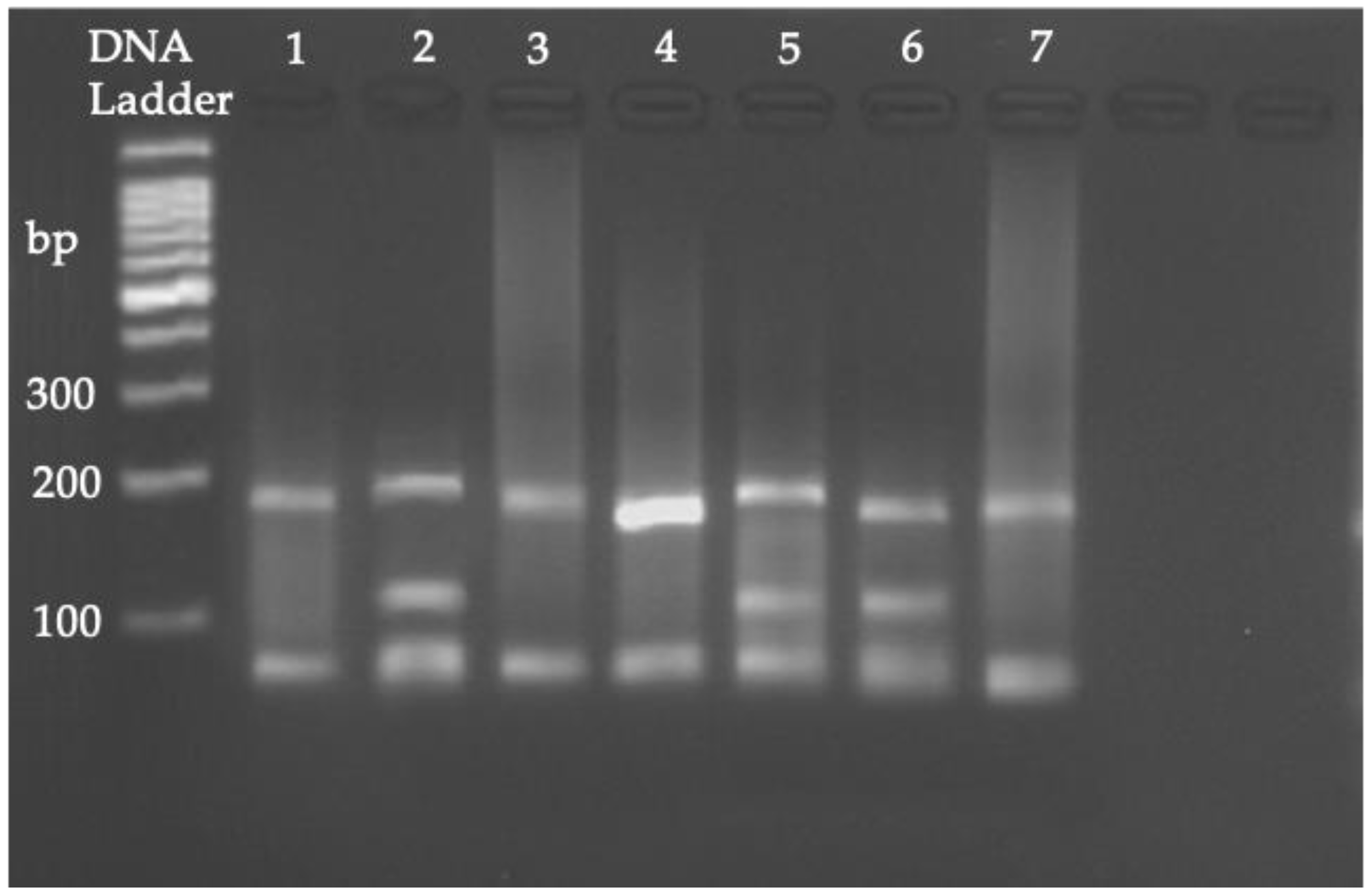
2.5. PCR, RFLP, and Sequencing
2.6. Statistical Analysis
3. Results
3.1. Coproparasitoscopic Analysis
3.2. PCR, RFLP, and Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Sequencing Analysis for the forward Primer G736 > B_Giardin_G376_Partial Sequence

{kind=link}
{kind=link}
| Description | Max Score | Total Score | Query Cover | E Value | Ident. | Accession |
|---|---|---|---|---|---|---|
| Giardia intestinalis isolate CAT clone 2 beta-giardin gene, complete cds | 496 | 496 | 99% | 1 × 10−135 | 99.63% | EU014386.1 |
| Giardia intestinalis isolate B7V_oct’11 note assemblage AI beta-giardin gene, partial cds | 497 | 496 | 100% | 3 × 10−136 | 99.63% | KF963547.1 |
| Giardia intestinalis isolate 34H beta-giardin gene, partial cds | 497 | 497 | 100% | 3 × 10−136 | 99.63% | DQ466765.1 |
Appendix A.2. Sequencing Analysis for the forward Primer G759 > B_Giardin_G759_Partial Sequence
| Description | Max Score | Total Score | Query Cover | E Value | Ident. | Accession |
|---|---|---|---|---|---|---|
| Giardia intestinalis isolate 9C beta-giardin gene, partial cds | 508 | 508 | 100% | 1 × 10−139 | 99.64% | DQ466729.1 |
| Giardia intestinalis isolate Z1 beta-giardin gene, partial cds | 507 | 507 | 99% | 5 × 10−139 | 99.64% | MN629930.1 |
| Giardia intestinalis strain VANC/96/UBC/126 beta-giardin (bg) gene, partial cds | 507 | 507 | 99% | 5 × 10−139 | 99.64% | KP687765.1 |
References
- Savioli, L.; Smith, H.; Thompson, A. Giardia and Cryptosporidium join the ‘Neglected Diseases Initiative’. Trends Parasitol. 2006, 22, 203–208. [Google Scholar] [CrossRef]
- Aw, J.Y.H.; Clarke, N.E.; McCarthy, J.S.; Traub, R.J.; Amaral, S.; Huque, H.; Andrews, R.M.; Gray, D.J.; Clements, A.C.A.; Nery, S.V. Giardia duodenalis infection in the context of a community-based deworming and water, sanitation and hygiene trial in Timor-Leste. Parasites Vectors 2019, 12, 491–510. [Google Scholar] [CrossRef]
- Rendtorff, R.C. The experimental transmission of human intestinal protozoan parasites: II. Giardia lamblia Cysts Gwen in Capsules. Am. J. Epidemiol. 1954, 59, 209–222. [Google Scholar] [CrossRef]
- Thompson, R. Giardiasis as a re-emerging infectious disease and its zoonotic potential. Int. J. Parasitol. 2000, 30, 1259–1267. [Google Scholar] [CrossRef] [PubMed]
- Balderrama-Carmona, A.P.; Gortares-Moroyoqui, P.; Moran-Palacio, E.F.; Ulloa-Mercado, R.G.; Diaz-Tenorio, L.M.; Leyva-Soto, L.A. Risk assesmet for Giardia in enviromental samples. In Current Topics in Giardiasis, 1st ed.; Alfonso Rodriguez Morales; IntechOpen: Ryjeca, Croacia, 2017; pp. 147–164. [Google Scholar] [CrossRef] [Green Version]
- Lujan, H.D. Giardia y Giardiasis. Medicina 2006, 66, 70–74. [Google Scholar] [PubMed]
- Leung, A.K.; Leung, A.A.; Wong, A.H.; Sergi, C.M.; Kam, J.K. Giardiasis: An Overview. Recent Pat. Inflamm. Allergy Drug Discov. 2019, 13, 134–143. [Google Scholar] [CrossRef]
- Adam, R.D. Biology of Giardia lamblia. Clin. Microbiol. Rev. 2001, 14, 447–475. [Google Scholar] [CrossRef] [Green Version]
- Paget, T.A.; Macechko, P.T.; Jarroll, E.L. Metabolic Changes in Giardia intestinalis during Differentiation. J. Parasitol. 1998, 84, 222–226. [Google Scholar] [CrossRef]
- Heresi, G.P.; Murphy, J.R.; Cleary, T.G. Giardiasis. Semin. Pediatr. Infect. Dis. 2000, 11, 189–195. [Google Scholar] [CrossRef]
- Thompson, R.; Reynoldson, J.; Mendis, A. Giardia and Giardiasis. Adv. Parasitol. 1993, 32, 71–160. [Google Scholar] [CrossRef] [PubMed]
- Williamson, A.; O’Donoghue, P.; Upcroft, J.; Upcroft, P. Immune and pathophysiological responses to different strains of Giardia duodenalis in neonatal mice. Int. J. Parasitol. 2000, 30, 129–136. [Google Scholar] [CrossRef] [PubMed]
- El-Basha, N.R.; Zaki, M.M.; Hassanin, O.M.; Rehan, M.K.; Omran, D. Giardia Assemblages A and B in Diarrheic Patients: A Comparative Study in Egyptian Children and Adults. J. Parasitol. 2016, 102, 69–74. [Google Scholar] [CrossRef]
- Puebla, L.E.J.; Núñez, F.A.; Silva, I.M.; Rivero, L.R.; González, M.M.; Sutil, Y.M.; Valdés, L.A.; Millán, I.A.; Müller, N. Molecular Characterization and Risk Factors of Giardia duodenalisamong School Children from La Habana, Cuba. J. Parasitol. Res. 2015, 2015, 378643. [Google Scholar] [CrossRef] [Green Version]
- Nash, T.E.; Herrington, D.A.; Losonsky, G.A.; Levine, M.M. Experimental Human Infections with Giardia lamblia. J. Infect. Dis. 1987, 156, 974–984. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Cardoso, E.; Eligio-García, L.; Cortés-Campos, A.; Estrada, A.C.; Pinto-Sagahón, M.; Noguera-Estrada, C. The frequency of intestinal parasites in puppies from Mexican kennels. Health 2010, 02, 1316–1319. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Ocampo, S.; Cotte-Alzate, J.D.; Escobedo, Á.A.; Rodríguez-Morales, A.J. Prevalence of zoonotic and non-zoonotic genotypes of Giardia intestinalis in cats: A systematic review and meta-analysis. Infez. Med. 2017, 25, 326–338. [Google Scholar] [CrossRef]
- Cedillo, R.; Leal, Y.; Yepez, L.; Gomez, A.; Ortega, G.; Tapia, R.; Muñoz, O. Seroepidemiology of Giardiasis in Mexico. Am. J. Trop. Med. Hyg. 2009, 80, 6–10. [Google Scholar] [CrossRef]
- García-Cervantes, P.C.; Báez-Flores, M.E.; Delgado-Vargas, F.; Ponce-Macotela, M.; Nawa, Y.; De-La-Cruz-Otero, M.D.-C.; Martínez-Gordillo, M.N.; Díaz-Camacho, S.P. Giardia duodenalis genotypes among schoolchildren and their families and pets in urban and rural areas of Sinaloa, Mexico. J. Infect. Dev. Ctries. 2017, 11, 180–187. [Google Scholar] [CrossRef]
- Sommer, M.; Rupp, P.; Pietsch, M.; Kaspar, A.; Beelitz, P. Giardia in a selected population of dogs and cats in Germany—diagnostics, coinfections and assemblages. Vet. Parasitol. 2018, 249, 49–56. [Google Scholar] [CrossRef]
- Paoletti, B.; Otranto, D.; Weigl, S.; Giangaspero, A.; Di Cesare, A.; Traversa, D. Prevalence and genetic characterization of Giardia and Cryptosporidium in cats from Italy. Res. Vet. Sci. 2010, 91, 397–399. [Google Scholar] [CrossRef]
- Mircean, V.; Györke, A.; Jarca, A.; Cozma, V. Prevalence of Giardia species in stool samples by ELISA in household cats from Romania and risk factors. J. Feline Med. Surg. 2011, 13, 479–482. [Google Scholar] [CrossRef]
- Jaros, D.; Zygner, W.; Jaros, S.; Wędrychowicz, H. Detection of Giardia intestinalis Assemblages A, B and D in Domestic Cats from Warsaw, Poland. Pol. J. Microbiol. 2011, 60, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Vasilopulos, R.J.; Rickard, L.G.; Mackin, A.J.; Pharr, G.T.; Huston, C.L. Genotypic Analysis of Giardia duodenalis in Domestic Cats. J. Vet. Intern. Med. 2007, 21, 352–355. [Google Scholar] [CrossRef] [PubMed]
- De Santis-Kerr, A.C.; Raghavan, M.; Glickman, N.W.; Caldanaro, R.J.; Moore, G.E.; Lewis, H.B.; Schantz, P.M.; Glickman, L.T. Prevalence and risk factors for Giardia and coccidia species of pet cats in 2003–2004. J. Feline Med. Surg. 2006, 8, 292–301. [Google Scholar] [CrossRef]
- Itoh, N.; Muraoka, N.; Saeki, H.; Aoki, M.; Itagaki, T. Prevalence of Giardia intestinalis Infection in Dogs of Breeding Kennels in Japan. J. Vet. Med. Sci. 2005, 67, 717–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santín, M.; Trout, J.M.; Vecino, J.A.C.; Dubey, J.; Fayer, R. Cryptosporidium, Giardia and Enterocytozoon bieneusi in cats from Bogota (Colombia) and genotyping of isolates. Vet. Parasitol. 2006, 141, 334–339. [Google Scholar] [CrossRef]
- López-Arias, Á.; Villar, D.; López-Osorio, S.; Calle-Vélez, D.; Chaparro-Gutiérrez, J.J. Giardia is the most prevalent parasitic infection in dogs and cats with diarrhea in the city of Medellín, Colombia. Vet. Parasitol. Reg. Stud. Rep. 2019, 18, 100335. [Google Scholar] [CrossRef]
- Souza, S.L.; Gennari, S.M.; Richtzenhain, L.J.; Pena, H.F.; Funada, M.R.; Cortez, A.; Gregori, F.; Soares, R.M. Molecular identification of Giardia duodenalis isolates from humans, dogs, cats and cattle from the state of São Paulo, Brazil, by sequence analysis of fragments of glutamate dehydrogenase (gdh) coding gene. Vet. Parasitol. 2007, 149, 258–264. [Google Scholar] [CrossRef]
- López, J.; Abarca, K.; Paredes, P.; Inzunza, E. Parásitos intestinales en caninos y felinos con cuadros digestivos en Santiago, Chile: Consideraciones en Salud Pública. Revista Médica Chile 2006, 134, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Scorza, A.V.; Duncan, C.; Miles, L.; Lappin, M.R. Prevalence of selected zoonotic and vector-borne agents in dogs and cats in Costa Rica. Vet. Parasitol. 2011, 183, 178–183. [Google Scholar] [CrossRef]
- Thompson, R.A.; Palmer, C.S.; O’Handley, R. The public health and clinical significance of Giardia and Cryptosporidium in domestic animals. Vet. J. 2008, 177, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Gerhold, R.W.; Jessup, D.A. Zoonotic Diseases Associated with Free-Roaming Cats. Zoonoses Public Health 2012, 60, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Abe, N.; Kimata, I.; Tokoro, M. Genotyping of Giardia isolates from humans in Japan using the small subunit ribosomal RNA and glutamate dehydrogenase gene sequences. Jpn. J. Infect. Dis. 2005, 58, 57–58. [Google Scholar]
- Eligio-García, L.; Cortes-Campos, A.; Jiménez-Cardoso, E. Genotype of Giardia intestinalis isolates from children and dogs and its relationship to host origin. Parasitol. Res. 2005, 97, 1–6. [Google Scholar] [CrossRef]
- Monis, P.; Caccio, S.M.; Thompson, R.A. Variation in Giardia: Towards a taxonomic revision of the genus. Trends Parasitol. 2009, 25, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Otero-Negrete, J.J.; Martínez-Godillo, M.N.; Ibarra-Velarde, F.; Ponce-Macotela, M. Prevalence of Giardia Intestinalis and Zo-onotic Genotype Predominance in Small Scale Sheep and Cattle Farms in Five States of Mexican Republic. Veterinaria Mexico 2011, 42, 219–226. [Google Scholar]
- Godínez-Galaz, E.M.; Veyna-Salazar, N.P.; Olvera-Ramírez, A.M.; Milián-Suazo, F.; Perea-Razo, C.A.; Bernal-Reynaga, R.; Cantó-Alarcón, G.J. Prevalence and Zoonotic Potential of Giardia intestinalis in Dogs of the Central Region of Mexico. Animals 2019, 9, 325. [Google Scholar] [CrossRef] [Green Version]
- Dryden, M.W.; Payne, P.A.; Smith, V. Accurate diagnosis of Giardia spp and proper fecal examination procedures. Vet Ther. 2006, 7, 4–14. [Google Scholar]
- Minvielle, M.C.; Molina, N.B.; Polverino, D.; Basualdo, J. First genotyping of Giardia lamblia from human and animal feces in Argentina, South America. Mem. Inst. Oswaldo Cruz 2008, 103, 98–103. [Google Scholar] [CrossRef]
- Ghosh, S.; Debnath, A.; Sil, A.; De, S.; Chattopadhyay, D.; Das, P. PCR detection of Giardia lamblia in stool: Targeting intergenic spacer region of multicopy rRNA gene. Mol. Cell. Probes 2000, 14, 181–189. [Google Scholar] [CrossRef]
- Babaei, Z.; Oormazdi, H.; Rezaie, S.; Rezaeian, M.; Razmjou, E. Giardia intestinalis: DNA extraction approaches to improve PCR results. Exp. Parasitol. 2011, 128, 159–162. [Google Scholar] [CrossRef]
- de Almeida, I.N.; Carvalho, W.D.S.; Rossetti, M.L.; Costa, E.R.D.; de Miranda, S.S. Evaluation of six different DNA extraction methods for detection of Mycobacterium tuberculosis by means of PCR-IS6110: Preliminary study. BMC Res. Notes 2013, 6, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacciò, S.M.; De Giacomo, M.; Pozio, E. Sequence analysis of the β-giardin gene and development of a polymerase chain reaction–restriction fragment length polymorphism assay to genotype Giardiaduodenalis cysts from human faecal samples. Int. J. Parasitol. 2002, 32, 1023–1030. [Google Scholar] [CrossRef]
- Nikolic, A.; Dimitrijevic, S.; Djurkovic-Djarkovic, O.; Bobic, B.; Maksimovic-Mihajlovic, O. Giardiasis in Dogs and Cats in the Belgrade Area. Acta Vet. Brno. 2002, 52, 43–48. [Google Scholar]
- Bouzid, M.; Halai, K.; Jeffreys, D.; Hunter, P.R. The prevalence of Giardia infection in dogs and cats, a systematic review and meta-analysis of prevalence studies from stool samples. Vet. Parasitol. 2015, 207, 181–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall´agnol, L.P.; Otto, M.A.; da Silva, A.S.; Gonzalez Monteiro, S. Gastrointestinal Parasites in Naturally Infected Cats in the Municipality of Santa Maria in Rio Grande Do Sul, Brazil. Vet. Bras. 2010, 4, 181–184. [Google Scholar]
- Fernandes de Mendoça, F.; Pitella, A.; Emmerick, S.; Pereira, N. Assesment of the diagnostic performances of four methods for detection of Giardia duodenalis in fecal samplesfrom human, canineand feline carriers. J. Microbiol. Methods 2018, 145, 73–78. [Google Scholar] [CrossRef]
- Decock, C.; Cadiergues, M.C.; Larcher, M.; Vermot, S.; Franc, M. Comparison of two techniques for diagnosis of giardiasis in dogs. Parasite 2003, 10, 69–72. [Google Scholar] [CrossRef]
- Chalkowski, K.; Wilson, A.E.; Lepczyk, C.A.; Zohdy, S. Who let the cats out? A global meta-analysis on risk of parasitic infection in indoor versus outdoor domestic cats (Felis catus). Biol. Lett. 2019, 15, 20180840. [Google Scholar] [CrossRef] [Green Version]
- Singer, S.M.; Nash, T.E. The Role of Normal Flora in Giardia lamblia Infections in Mice. J. Infect. Dis. 2000, 181, 1510–1512. [Google Scholar] [CrossRef] [Green Version]
- Nydam, D.V.; Wade, S.E.; Schaaf, S.L.; Mohammed, H.O. Number of Cryptosporidium parvum oocysts or Giardia spp cysts shed by dairy calves after natural infection. Am. J. Vet. Res. 2001, 62, 1612–1615. [Google Scholar] [CrossRef]
- Mosallanejad, B.; Avizeh, R.; Jalali, M.R.; Alborzi, A. Prevalence of Giardia duodenalis Infection in Household Cats of Ahvaz District, South-West of Iran. Iran. J. Parasitol. 2010, 5, 27–34. [Google Scholar] [PubMed]
- Mekaru, S.R.; Marks, S.L.; Felley, A.J.; Chouicha, N.; Kass, P.H. Comparison of Direct Immunofluorescence, Immunoassays, and Fecal Flotation for Detection of Cryptosporidium spp. and Giardia spp. in Naturally Exposed Cats in 4 Northern California Animal Shelters. J. Vet. Intern. Med. 2007, 21, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Sursal, N.; Simsek, E.; Yildiz, K. Feline Giardiasis in Turkey: Prevalence and Genetic and Haplotype Diversity of Giardia duodenalis Based on the β-Giardin Gene Sequence in Symptomatic Cats. J. Parasitol. 2020, 106, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.S.; Johansen, M.; Jorsal, S.E.; Nielsen, J.P.; Bækbo, P.; Angen, Ø. Pooling of porcine fecal samples for quantification of Lawsonia intracellularis by real-time polymerase chain reaction. J. Vet. Diagn. Investig. 2014, 26, 342–345. [Google Scholar] [CrossRef] [Green Version]
- Eysker, M.; Bakker, J.; Berg, M.V.D.; van Doorn, D.; Ploeger, H. The use of age-clustered pooled faecal samples for monitoring worm control in horses. Vet. Parasitol. 2008, 151, 249–255. [Google Scholar] [CrossRef]
- Papaiakovou, M.; Wright, J.; Pilotte, N.; Chooneea, D.; Schär, F.; Truscott, J.E.; Dunn, J.C.; Gardiner, I.; Walson, J.L.; Williams, S.A.; et al. Pooling as a strategy for the timely diagnosis of soil-transmitted helminths in stool: Value and reproducibility. Parasites Vectors 2019, 12, 443. [Google Scholar] [CrossRef] [Green Version]
- Singer, R.S.; Cooke, C.L.; Maddox, C.W.; Isaacson, R.E.; Wallace, R.L. Use of Pooled Samples for the Detection of Salmonella in Feces by Polymerase Chain Reaction. J. Vet. Diagn. Investig. 2006, 18, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Clasen, J.; Mellerup, A.; Olsen, J.E.; Angen, Ø.; Folkesson, A.; Halasa, T.; Toft, N.; Birkegård, A.C. Determining the optimal number of individual samples to pool for quantification of average herd levels of antimicrobial resistance genes in Danish pig herds using high-throughput qPCR. Vet. Microbiol. 2016, 189, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Covacin, C.; Aucoin, D.; Elliot, A.; Thompson, R. Genotypic characterisation of Giardia from domestic dogs in the USA. Vet. Parasitol. 2011, 177, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Torres-Romero, J.; Euan-Canto, A.D.J.; Benito-González, N.; Padilla-Montaño, N.; Huchin-Chan, C.; Lara-Riegos, J.; Cedillo-Rivera, R. Intestinal parasites and genotyping of Giardia duodenalis in children: First report of genotype B in isolates from human clinical samples in Mexico. Mem. Inst. Oswaldo Cruz 2014, 109, 388–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Procesi, I.G.; Carnio, A.; Berrilli, F.; Di Filippo, M.M.; Scarito, A.; Amoruso, C.; Barni, M.; Ruffini, M.; Barlozzari, G.; Scarpulla, M.; et al. Giardia duodenalis in colony stray cats from Italy. Zoonoses Public Health 2021, 69, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Zanzani, S.; Gazzonis, A.L.; Scarpa, P.; Berrilli, F.; Manfredi, M.T. Intestinal Parasites of Owned Dogs and Cats from Metropolitan and Micropolitan Areas: Prevalence, Zoonotic Risks, and Pet Owner Awareness in Northern Italy. BioMed. Res. Int. 2014, 2014, 696508. [Google Scholar] [CrossRef] [PubMed]
- Papini, R.; Cardini, G.; Paoletti, B.; Giangaspero, A. Detection of Giardia assemblage A in cats in Florence, Italy. Parasitol. Res. 2006, 100, 653–656. [Google Scholar] [CrossRef]
- Palmer, C.S.; Traub, R.; Robertson, I.; Devlin, G.; Rees, R.; Thompson, R.A. Determining the zoonotic significance of Giardia and Cryptosporidium in Australian dogs and cats. Vet. Parasitol. 2008, 154, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dan, X.; Zhu, K.; Li, N.; Guo, Y.; Zheng, Z.; Feng, Y.; Xiao, L. Genetic characterization of Cryptosporidium spp. and Giardia duodenalis in dogs and cats in Guangdong, China. Parasites Vectors 2019, 12, 571. [Google Scholar] [CrossRef]
- Suzuki, J.; Murata, R.; Kobayashi, S.; Sadamasu, K.; Kai, A.; Takeuchi, T. Risk of human infection with Giardia duodenalis from cats in Japan and genotyping of the isolates to assess the route of infection in cats. Parasitology 2010, 138, 493–500. [Google Scholar] [CrossRef]
- Sahagún, J.; Clavel, A.; Goñi, P.; Seral, C.; Llorente, M.T.; Castillo, F.J.; Capilla, S.; Arias, A.; Gómez-Lus, R. Correlation between the presence of symptoms and the Giardia duodenalis genotype. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 27, 81–83. [Google Scholar] [CrossRef]
- Ballweber, L.R.; Xiao, L.; Bowman, D.D.; Kahn, G.; Cama, V.A. Giardiasis in dogs and cats: Update on epidemiology and public health significance. Trends Parasitol. 2010, 26, 180–189. [Google Scholar] [CrossRef]
- Read, C.M.; Monis, P.T.; Thompson, R.A. Discrimination of all genotypes of Giardia duodenalis at the glutamate dehydrogenase locus using PCR-RFLP. Infect. Genet. Evol. 2004, 4, 125–130. [Google Scholar] [CrossRef]
- van Keulen, H.; Macechko, P.; Wade, S.; Schaaf, S.; Wallis, P.M.; Erlandsen, S.L. Presence of human Giardia in domestic, farm and wild animals, and environmental samples suggests a zoonotic potential for giardiasis. Vet. Parasitol. 2002, 108, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Instituto Nacional de Estadística, Geografía e informática. Primera Encuesta Nacional de Bienestar Autorreportado; INEGI: Aguascalientes, Mexico, 2021.


| Location | Age and Origin | Total | Occurrence (%) | Assemblage | Reference |
|---|---|---|---|---|---|
| Germany | Owned, stray, Shelter (Adults and kittens) | 145 | 17.9% | D | 2018 [20] |
| Italy | Owned, Stray (Kittens, Adults) | 181 | 6.1% | A, F | 2011 [21] |
| Romania | Owned (Kittens, Adults) | 181 | 28% | Not reported | 2011 [22] |
| Poland | Owned (No age) | 160 | 3.75% | A, B, D | 2011 [23] |
| USA | Owned (No age) | 250 | 13% | AI, AII, F | 2007 [24] |
| USA | Owned (Kittens, Adults) | 211,105 | 0.58% | Not reported | 2006 [25] |
| Japan | Kennels, Owned (Kittens, Adults) | 600 | 40% | Not reported | 2005 [26] |
| Colombia | Stray (Kittens, Adults) | 46 | 6.5% | F | 2006 [27] |
| Colombia | Owned (Kittens, Adults) | 203 | 20% | Not reported | 2019 [28] |
| Brazil | Owned, shelters (No age) | 19 | Not reported | AI, F | 2007 [29] |
| Chile | Owned (No age) | 230 | 19% | Not reported | 2006 [30] |
| Costa Rica | Owned (No age) | 9 | 57.1% | Not reported | 2011 [31] |
| Positive | Negative | Occurrence (%) | p Value | Odds Ratio | 95% CI | |
|---|---|---|---|---|---|---|
| Total | 50 | 150 | 25% | |||
| Strata of origin | ||||||
| Owned | 29 | 74 | 28 | 0.28827 | 1.41 | 0.731–2.7 |
| Unowned | 21 | 76 | 21 | |||
| Age | ||||||
| <6 Months | 24 | 48 | 33% | 0.041227 | 1.96 | 1.02–3.76 |
| >6 Months | 26 | 102 | 20% | |||
| Stool consistency | ||||||
| Pasty | 27 | 34 | 44% | 0.000031 | 4.00 | 2.03–7.8 |
| Firm | 23 | 116 | 16% | |||
| Sex | ||||||
| Male | 22 | 69 | 24% | 0.805723 | 0.92 | 0.48–1.75 |
| Female | 28 | 81 | 25% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veyna-Salazar, N.P.; Cantó-Alarcón, G.J.; Olvera-Ramírez, A.M.; Ruiz-López, F.J.; Bernal-Reynaga, R.; Bárcenas-Reyes, I.; Durán-Aguilar, M. Occurrence of Giardia duodenalis in Cats from Queretaro and the Risk to Public Health. Animals 2023, 13, 1098. https://doi.org/10.3390/ani13061098
Veyna-Salazar NP, Cantó-Alarcón GJ, Olvera-Ramírez AM, Ruiz-López FJ, Bernal-Reynaga R, Bárcenas-Reyes I, Durán-Aguilar M. Occurrence of Giardia duodenalis in Cats from Queretaro and the Risk to Public Health. Animals. 2023; 13(6):1098. https://doi.org/10.3390/ani13061098
Chicago/Turabian StyleVeyna-Salazar, Nerina P., Germinal J. Cantó-Alarcón, Andrea M. Olvera-Ramírez, Felipe J. Ruiz-López, Rodolfo Bernal-Reynaga, Isabel Bárcenas-Reyes, and Marina Durán-Aguilar. 2023. "Occurrence of Giardia duodenalis in Cats from Queretaro and the Risk to Public Health" Animals 13, no. 6: 1098. https://doi.org/10.3390/ani13061098