1. Introduction
As global demand for animal protein increases, the entire animal production system is gradually moving towards intensification [
1]. The aquaculture industry is growing rapidly, but it is vulnerable to the interactions between the animals themselves, diseases, and the environment. Therefore, a new model of aquaculture management strategy has emerged as a result of a growing understanding of animal nutrition and feed. The core objectives of this model are to minimize the effects of stressors by neutralizing free radicals, repairing oxidative damage to biological macromolecules and membrane systems, enhancing immunity, and maintaining normal physiological homeostasis. The key points of this model are antioxidant supplementation and increasing endogenous cellular antioxidants [
1]. Supplementation of diets with antioxidants can improve the resistance of fish to environmental stresses and is an essential measure to reduce losses in the aquaculture industry [
2]. Alpha-lipoic acid (α-LA), also known as 1,2-dithiolane-3-valeric acid, with the molecular formula C
8H
14O
2S
2, was first isolated from pig liver by Lester J. Reed in 1951 [
3]. α-LA is a naturally occurring compound found in microorganisms, plants, and animals, and is considered to be an “ideal antioxidant” or “universal antioxidant” because of its strong antioxidant capacity [
4,
5].
Studies have shown that α-LA can improve the survival rate, growth performance, and immunity of fish, and also improve the nutritional value of fish, which makes α-LA suitable for application in aquaculture [
6]. For example, dietary supplementation with an appropriate amount of α-LA could promote growth, fatty acid β-oxidation, and lipolysis of grass carp (
Ctenopharyngodon idellus; Cuvier et Valenciennes, 1844), increase protein deposition, enhance immunity and antioxidant capacity, alleviate the inflammatory response, and reduce lipid oxidative damage. It also could promote the expression of peripheral anorexia factor mRNA and reduce the expression of peripheral appetite factor mRNA, thus, reducing the intake and body weight of grass carp [
7,
8,
9,
10]. The enhancement of growth performance has also been found in other aquatic organisms with moderate amounts of α-LA in their diets, such as Nile tilapia (
Oreochromis niloticus; Linnaeus, 1758) [
11], African catfish (
Clarias gariepinus; Burchell, 1822) [
2], giant gourami (
Osphronemus goramy; Lacepède, 1801) [
12], Chinese mitten crab (
Eriocheir sinensis; H. Milne Edwards, 1853) [
13], and northern snakehead (
Channa argus; Cantor, 1842) [
14]. In addition, dietary supplementation with moderate amounts of α-LA could promote the expression of gluconeogenesis-related genes induced by a high-fat diet in fish, reduce lipid accumulation under high-fat conditions [
15], and enhance starch utilization in carp (
Cyprinus carpio; Linnaeus, 1758) [
16].
The hybrid grouper (
Epinephelus fuscoguttatus♀; Forsskål, 1775 ×
Epinephelus polyphekadion♂; Bleeker, 1849) is an important mariculture fish in southern China, with the characteristics of rapid growth and strong stress resistance, and it has a high economic value in China [
17,
18,
19,
20]. Although α-LA has been studied for over 70 years and there have been numerous studies on its addition as an antioxidant of aquatic animal diets, there have been few studies of α-LA in terms of the supplementation of marine fish diets. There are no reports of α-LA being added to the diet of groupers. Therefore, the purpose of this experiment was to research the effects of diet supplementation with α-LA on the growth performance, serum biochemical indexes, hepatic morphology, antioxidant capacity, and transcriptome of juvenile hybrid grouper fish, and to expand theoretical knowledge for the application of antioxidants in the hybrid grouper diet.
4. Discussion
α-LA is a multifunctional antioxidant that can promote growth performance as a feed additive for poultry animals [
24]. However, α-LA could also inhibit AMPKα in the hypothalamuses of chickens (
Gallus; Linnaeus, 1758) to reduce food intake [
25], and could activate AMPKα in the liver to inhibit the synthesis of glycogen synthase in the liver, resulting in a decrease in glycogen synthesis, thereby changing energy homeostasis and delaying the growth of chicken weight [
26]. Therefore, the growth-promoting effect of α-LA needs to be analyzed specifically in combination with the amount of α-LA added. In the study of α-LA as a diet supplement for aquatic animals, more studies have shown that with the increase in α-LA dose, the growth performance of aquatic animals exhibited a trend of increasing first and then decreasing, and high doses of α-LA were still able to improve the growth performance, such as in catfish [
2], giant gourami [
12], northern snakehead [
14], and Chinese mitten crab [
27]. However, some studies have suggested that high doses of α-LA had an inhibitory effect on the growth of aquatic animals, such as in Nile tilapia [
11] and oriental river prawn (
Macrobrachium nipponense; de Haan, 1849) [
28]. The recommended addition amounts were 439–528 mg/kg and 1354.8 mg/kg, respectively, but their growth performance was inhibited at 2400 mg/kg and 4000 mg/kg, respectively. In the present study, dietary supplementation with low doses of α-LA (0.4 and 0.6 g/kg) significantly reduced the WGR of juvenile hybrid groupers. Similar to the experimental results of Zhang et al. [
29], the addition of lower α-LA to the diet reduced the WGR of abalone (
Haliotis discus hannai; Ino, 1952), which may be the result of α-LA increasing energy consumption in juvenile hybrid groupers [
30]. However, the addition of 1.2 g/kg α-LA had no significant effect on the WGR of juvenile hybrid groupers, which may be the result of a high dose of α-LA promoting lipolysis to consume energy by activating the AMPKα-ATGL pathway without causing weight loss [
9]. In addition, Ding et al. [
31] found that α-LA in diets could reduce the growth rate of oriental river prawn fed with low carbohydrate diet but had no significant effect on the growth rate of oriental river prawn fed with high carbohydrate diet. This indicates that the composition of the diet may affect the mechanism of α-LA. Huang et al. [
7] discovered that dietary supplementation of 1.2 g/kg α-LA could inhibit the growth performance of grass carp. In this experiment, dietary supplementation of 1.2 g/kg α-LA had no significant effect on the WGR of juvenile hybrid groupers, indicating that different species had different sensitivities to α-LA. At present, the optimal α-LA addition amount for juvenile hybrid groupers with WGR as a reference still needs further study, and 1.2 g/kg α-LA has a certain reference value.
Serum biochemical indexes can reflect the overall health status, physiological stress response, and nutritional status of fish [
32,
33]. TG and TCHO are the main components of blood lipids [
34,
35]. The contents of TG and TCHO in serum are important indicators to measure lipid metabolism in fish [
36,
37]. Studies have shown that α-LA has the effect of lowering blood lipids and could reduce the content of TG and TCHO in mice and rats [
38,
39,
40]. Samuki et al. [
12] reported that dietary supplementation of 0.3, 0.6, and 0.9 g/kg α-LA reduced the content of TG in the serum of giant gourami; similarly, Siagian et al. [
2] also reported that 1.0 and 1.5 g/kg α-LA reduced the content of TG in the serum of African catfish. In this experiment, α-LA not only reduced the content of TG in L3 serum but also reduced the content of TCHO. Butler et al. [
41] suggested that α-LA could reduce the TG content in blood and the liver by inhibiting the expression of liver lipogenic genes, reducing hepatic TG secretion, and stimulating the clearance of TG-rich lipoproteins. Zulkhairi et al. [
42] believed that α-LA may reduce the TCHO content in the blood by cholesterol metabolism or lipoprotein lipase activity in the liver. TP and ALB are important indicators of protein synthesis and metabolism and immune function [
20,
43,
44]. Shi et al. [
9] discovered that α-LA regulates the AMPKα-CPT-1α pathway to reduce protein consumption in grass carp to increase protein deposition. In addition, Liu et al. [
8] found that α-LA could enhance the immune function of the grass carp skin, spleen, and head kidney. In this experiment, the contents of TP and ALB in the serum of L3 were significantly increased, but the growth performance of L3 did not change significantly. Therefore, the increase in TP and ALB may be the result of α-LA enhancing the immune function of juvenile hybrid groupers.
ALT and AST are low in serum and are mainly distributed in liver cells. When liver cells are damaged, they can release ALT and AST to increase their activity in serum, which is consistent with the extent of hepatic cell damage [
45]. In this experiment, the levels of ALT and AST in the serum of L3 decreased significantly, which was similar to the experimental results of adding α-LA in grass carp [
10] and African catfish [
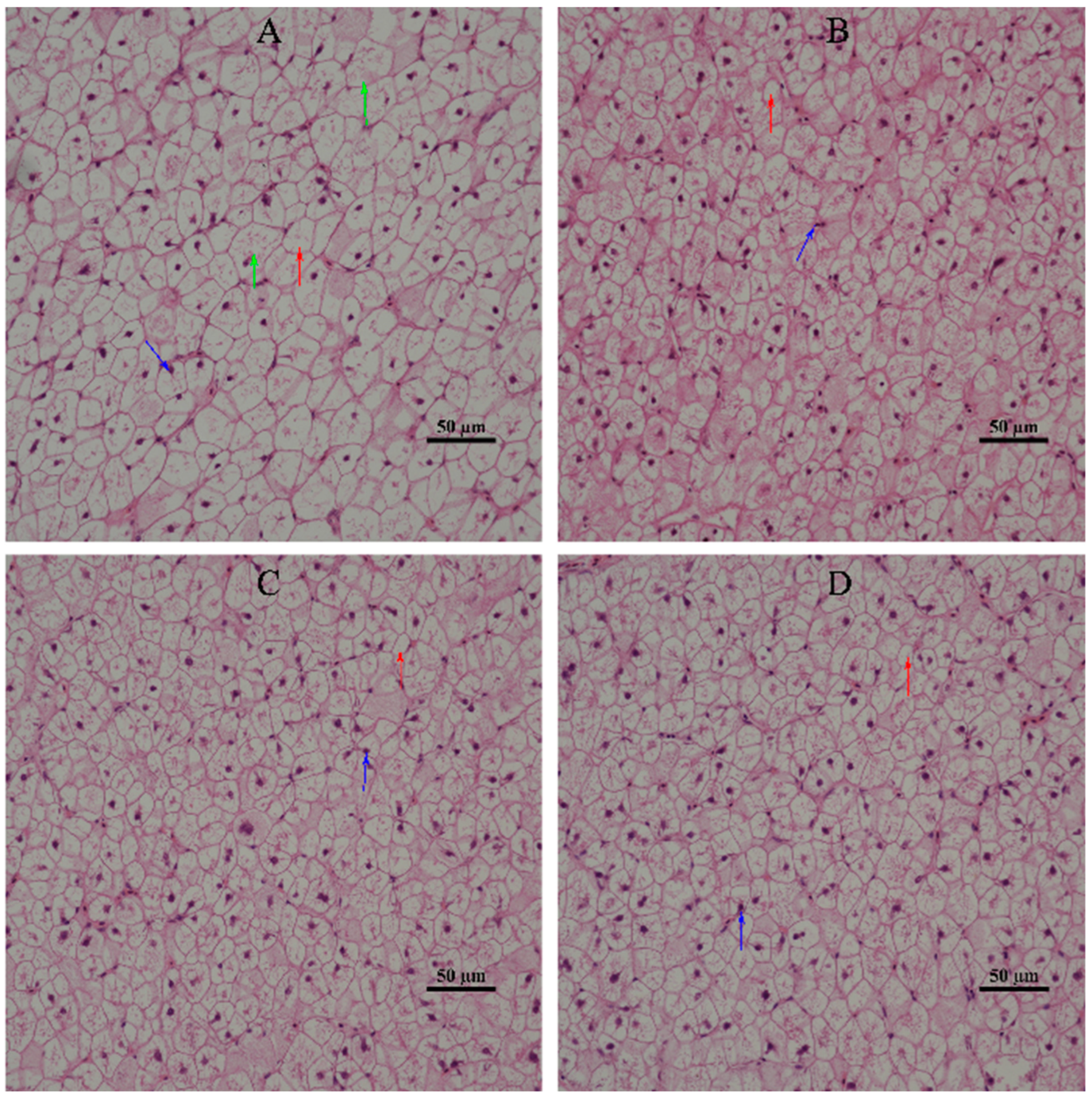
2]. This indicated that the degree of hepatic damage of L3 was lower than that of SL0, i.e., dietary supplementation of α-LA could improve the damage of liver cells in juvenile hybrid groupers. At the same time, by observing the morphology of hepatic tissue cells, it was found that compared with SL0, the morphology of hepatic tissue cells of juvenile hybrid groupers fed with α-LA was improved to varying degrees. This further confirmed that dietary supplementation of α-LA can improve hepatic cell damage.
The antioxidant system can protect fish from oxidative stress and is essential for fish health [
46]. Antioxidant enzymes (GSH-Px, CAT, and SOD) can scavenge free radicals to reduce oxidative stress, and their activities can reflect the health status of aquatic animals [
47]. GSH-Px can remove hydrogen peroxide and lipid peroxide in the body [
48]. SOD is a common antioxidant enzyme in the body and can remove superoxide anions [
49]. α-LA is considered to be an “ideal antioxidant” or “general antioxidant”, which can reduce oxidative damage and enhance the antioxidant defense systems of fish by scavenging excessive ROS and regenerating other antioxidants [
50,
51]. In this experiment, the activity of GSH-Px in the livers of juvenile hybrid groupers fed with α-LA was increased to varying degrees, and the activity of SOD in L2 and L3 was significantly increased. At present, many studies have reported similar results, and α-LA could improve the antioxidant capacity of aquatic animals. Xu et al. [
11] found that 0.3 g/kg α-LA significantly increased the activities of SOD and GSH-Px in the liver of Nile tilapia. Li et al. [
14] discovered that 600, 900, and 1200 mg/kg α-LA significantly increased the activities of SOD and GSH-Px in the liver, head kidney, and spleen of northern snakehead. Zhang et al. [
29] found that 800 mg/kg α-LA significantly increased the activities of SOD and GSH-Px in abalone. In summary, the results of this experiment showed that an appropriate amount of α-LA could increase the activity of antioxidant enzymes in the livers of juvenile hybrid groupers, thereby enhancing antioxidant capacity.
The transcriptome includes RNA transcripts expressed in a specific cell or tissue types under environmental conditions or specific developmental conditions [
52]. In recent years, transcriptome analysis has been widely used in aquaculture, and can be used for effective identification and expression analysis of candidate genes, such as growth, development, reproduction, disease, immunity, stress, and toxicology genes [
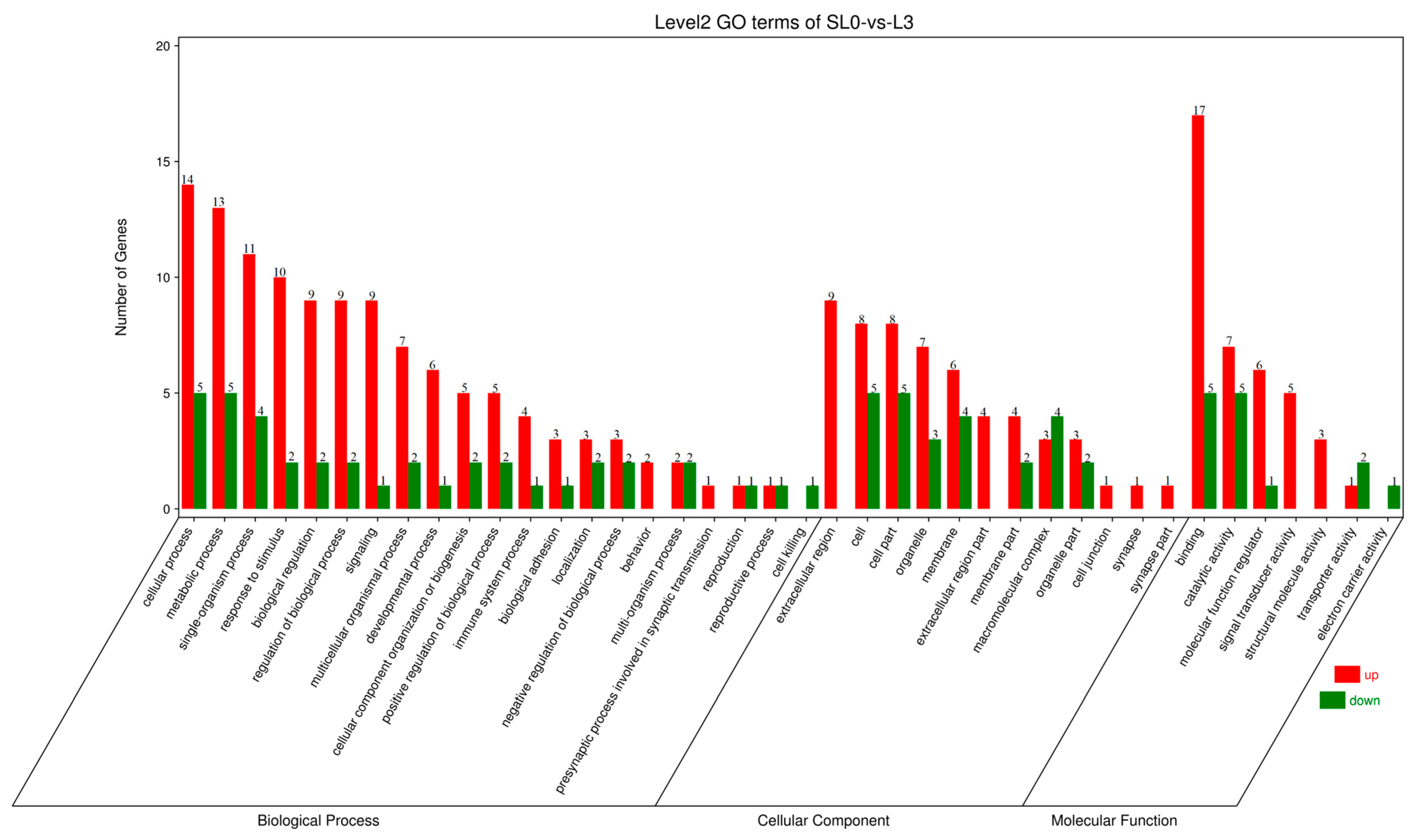
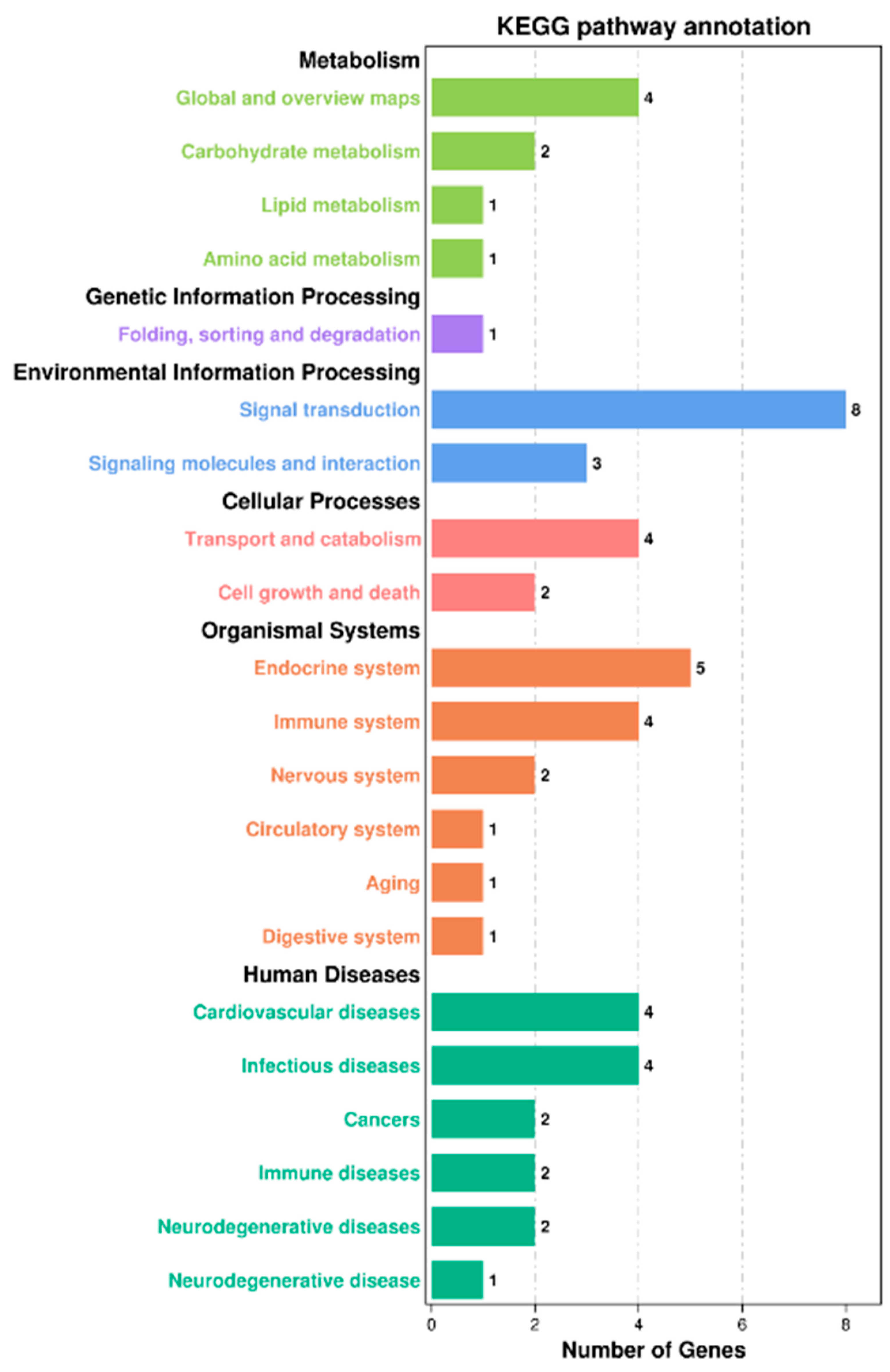
53]. In this experiment, according to serum biochemical indicators and liver antioxidant capacity, liver samples of SL0 and L3 were selected for transcriptome sequencing analysis. Functional analysis of the Kyoto Encyclopedia of Genes and Genomes (KEGG) showed that a total of 10,810 unigenes were annotated into 48 KEGG pathways, of which 12 pathways were significantly enriched, including the JAK/STAT signaling pathway, prolactin signaling pathway, antigen processing and presentation, glycolysis/gluconeogenesis, and so on.
The JAK/STAT signaling pathway is a common pathway for signal transduction of many cytokines, which is closely related to apoptosis, cell proliferation, inflammatory response, and differentiation. It is very important for coordinating adaptive immune mechanisms, initiating innate immunity, and inhibiting inflammatory responses [
54]. In this experiment, the JAK/STAT signaling pathway included three genes:
ifnk,
prl4a1, and
prl3b1. In addition, the prolactin signaling pathway also includes
prl4a1 and
prl3b1 genes. The
ifnk gene is a new type I interferon subclass [
55]. Interferons are proteins that are crucial to the human immune system. They are formed in various cells of fish, mammals, reptiles, and amphibians. IFN type I and IFN type II are found in ray-finned fish (Actinopterygii), and IFN type III is also found in phylogenetically older cartilaginous fishes. IFN type I in ray-finned fish (Actinopterygii) can activate the JAK/STAT signaling pathway and induce the expression of IFN-stimulated genes containing IFN-stimulated response elements complexes and, thus, possessing antiviral activity. In addition, in Perciformes, IFN I has been shown to exert antibacterial effects through macrophage phagocytosis [
56]. The grouper belongs to Osteichthyes, Actinopterygii, and Perciformes. Both
prl4a1 and
prl3b1 are members of the prolactin family, and prolactin is a multifunctional polypeptide hormone with immunomodulatory and protective effects [
57]. Studies have shown that prolactin can induce the expression of genes encoding major phagocytic NADPH oxidase components and ROS production in fish macrophages through the JAK2/Stat/IRF-1 signaling pathway [
58]. Antigen processing and presentation is the mechanism by which the entire antigen is degraded and loaded onto MHC molecules (class I and II) to display on the cell surface of T cells [
59]. Zhang and Chen [
60] found that a novel CC chemokine may be involved in the adaptive immune response by regulating MHC class I antigen processing and presentation in large yellow croaker (
Pseudosciaena crocea; Richardson, 1846). In this experiment, only the expression of the
ctsl gene was significantly up-regulated in antigen processing and presentation. Cathepsin L (
ctsl) is a member of the papain family of cysteine proteases [
61] which plays an important role in the biological activities of fish, including antigen processing [
62], antigen presentation [
63], protein degradation [
64], and anti-microbial invasion [
65]. Recently, the key role of
ctsl in the innate immune system of many fish species has been further revealed [
66]. In summary, combined with the significant increase in TP and ALB in the serum of L3 and the significant up-regulation in
ifnk,
prl4a1,
prl3b1, and
ctsl in the JAK/STAT signaling pathway, prolactin signaling pathway, and antigen processing and presentation in liver, it is speculated that dietary supplementation of α-LA can enhance the immune function of juvenile hybrid groupers by regulating the JAK/STAT signaling pathway, prolactin signaling pathway, and antigen processing and presentation.
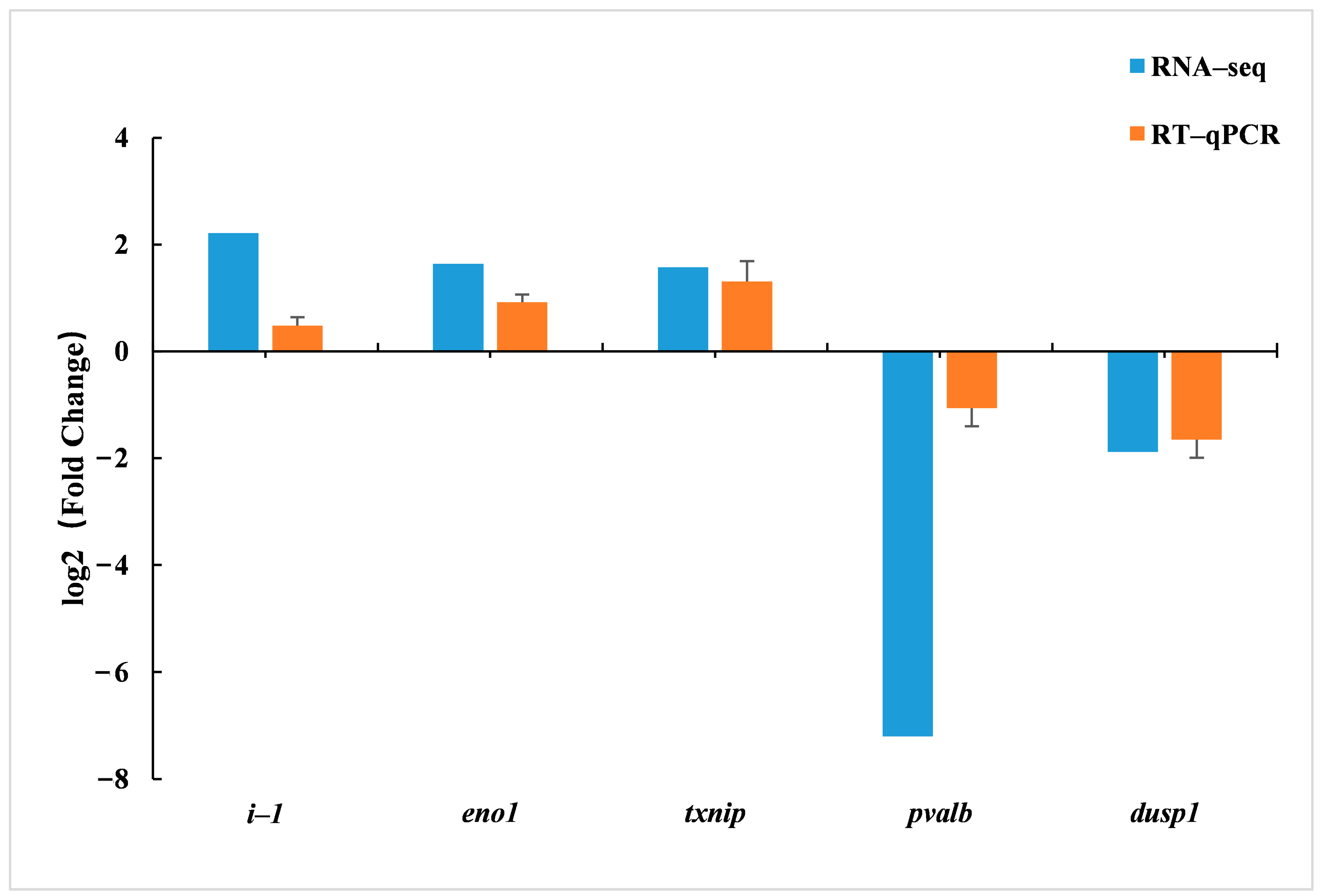
Glycolysis/gluconeogenesis is an opposing metabolic pathway involved in carbohydrate degradation and synthesis and plays an important role in maintaining glucose homeostasis [
67]. In this experiment, the glycolysis/gluconeogenesis pathway included two genes,
gapdh and
eno1. Glyceraldehyde-3-phosphate dehydrogenase (
gapdh) plays a key role in the glycolytic pathway. It can catalyze the formation of glyceraldehyde-3-phosphate to 1,3-bisphosphoglycerate, which produces NADH. NADH can synthesize ATP through the electron transport chain in mitochondria [
68]. α-enolase (
eno1) plays a functional role in glycolysis/gluconeogenesis. It can catalyze the conversion of 2-phosphate-D-glycerate to phosphoenolpyruvic acid during glycolysis and phosphoenolpyruvic acid to 2-phosphate-D-glycerate during glycogen synthesis [
69]. Huang et al. [
15] showed that the addition of α-LA to the diet could enhance the expression of glycolysis, gluconeogenesis, and glucose transport-related genes in zebrafish (
Danio rerio; Hamilton, 1822) livers. In this experiment, the expression of
gapdh gene was significantly down-regulated, and the expression of eno1 gene was significantly up-regulated. Therefore, it is speculated that α-LA can maintain the glucose homeostasis of juvenile hybrid groupers by regulating the expression of
gapdh and
eno1 genes in the glycolysis/gluconeogenesis pathway.
Therefore, the optimal addition of α-LA in the diet of hybrid groupers needs further study. However, this experiment showed that without affecting the growth of hybrid groupers, it could reduce the blood lipid level of hybrid groupers, improve the damage of liver cells, and increase the activity of antioxidant enzymes in the liver. This indicates that an appropriate amount of α-LA can be used as an additive to improve fish health in actual production. In addition, the transcriptome results provide some theoretical knowledge for the further study of α-LA in immune and glucose homeostasis.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}