
Effects of the Replacement of Dietary Fish Meal with Poultry By-Product Meal on Growth and Intestinal Health of Chinese Soft-Shelled Turtle (Pelodiscus sinensis)

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals, Diets, and Feeding Trial
2.2. Sample Collection
2.3. Biochemical Analysis
2.4. Intestinal Histological Analysis
2.5. Analysis of Intestinal and Liver Gene Expression
2.6. Statistical Analysis
3. Results
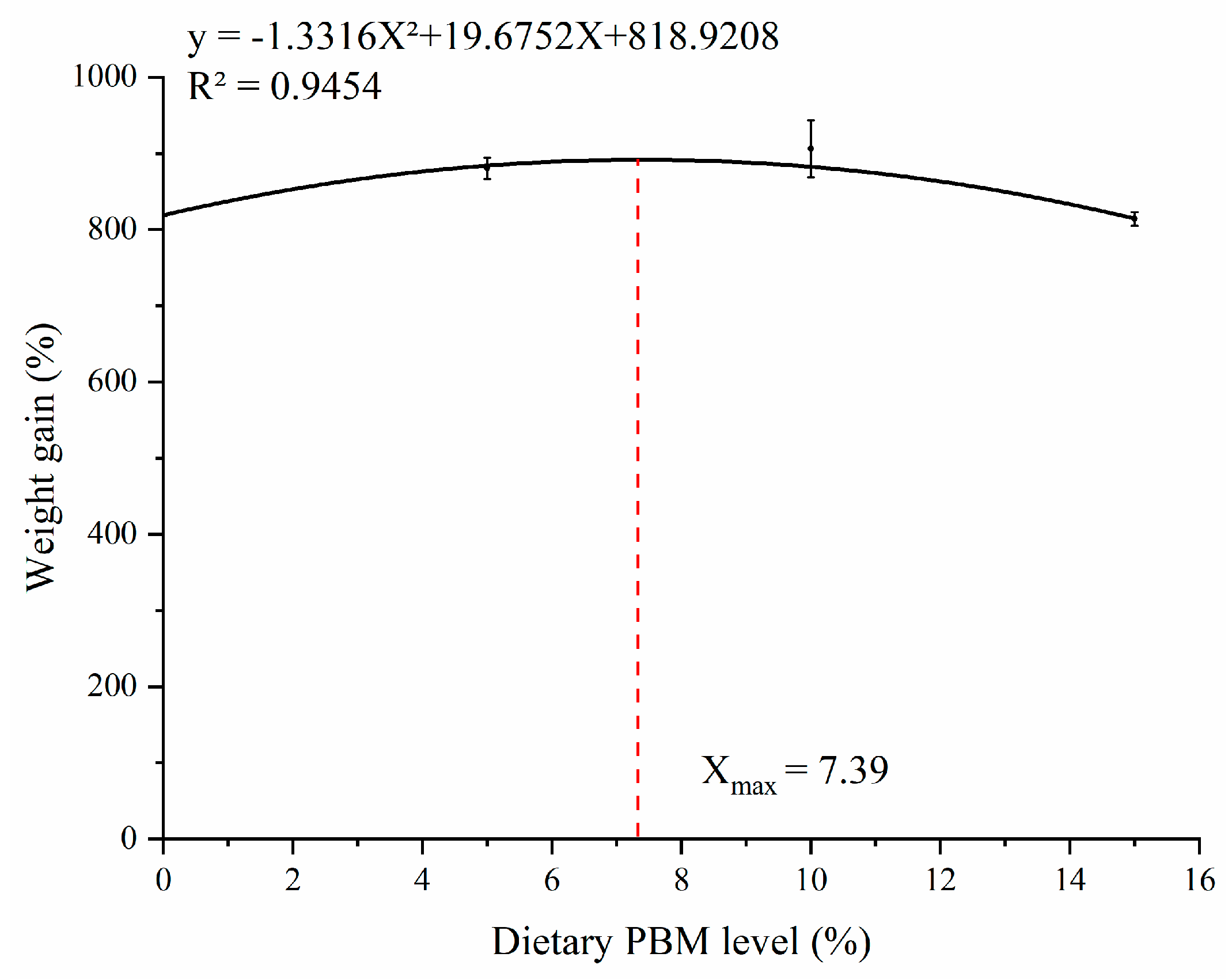
3.1. Growth Performance
3.2. whole-body Composition
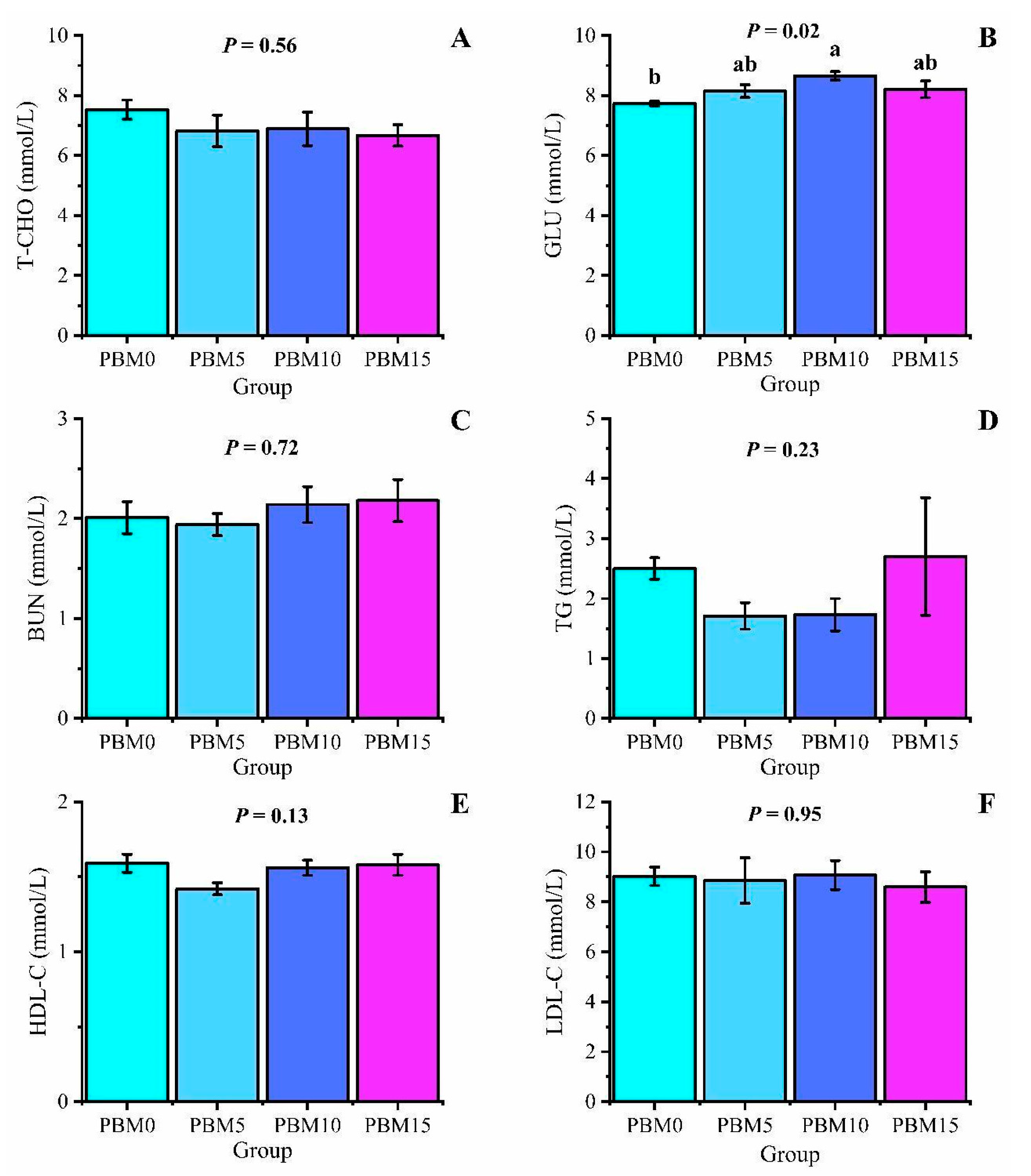
3.3. Serum Biochemical Indicators
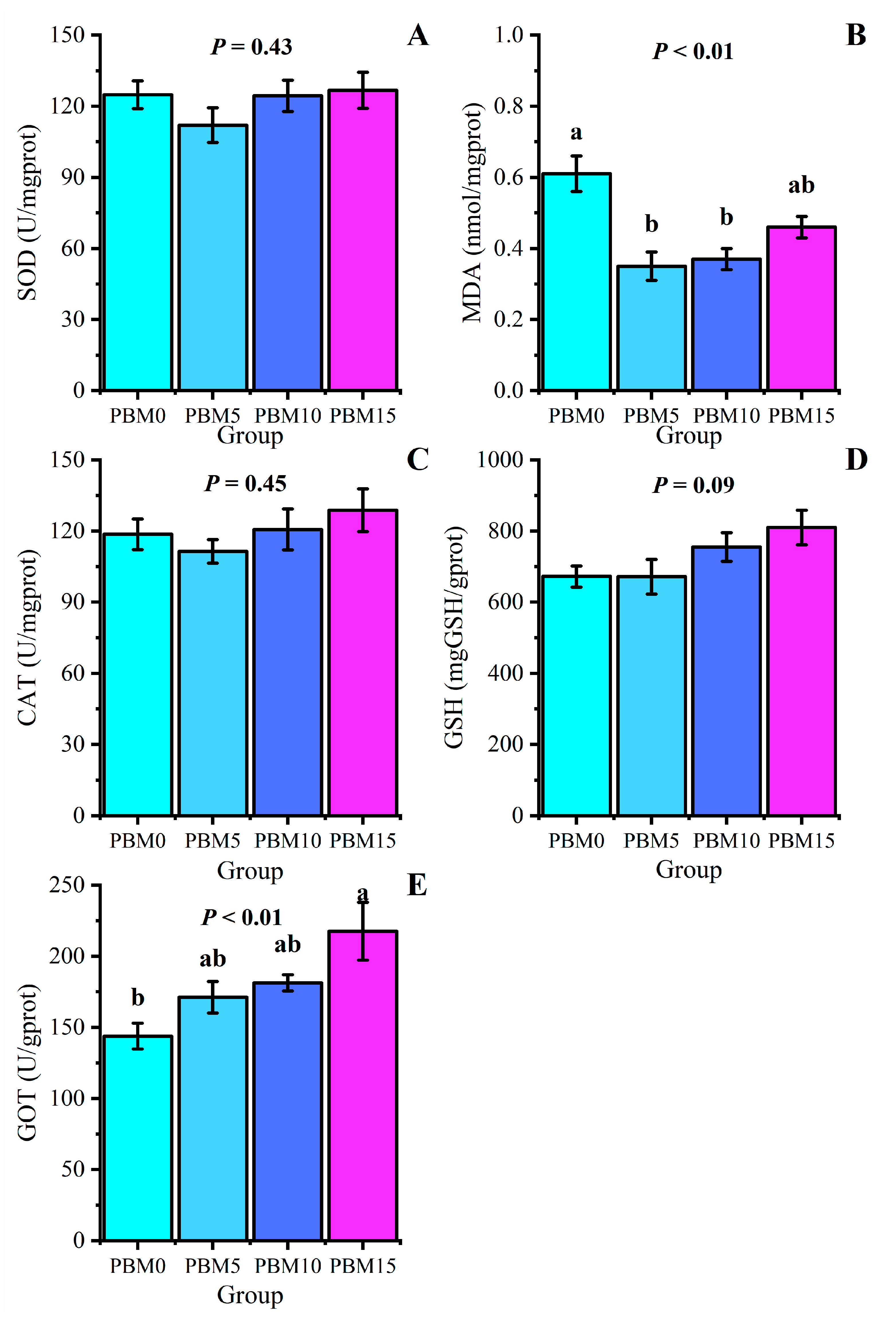
3.4. Antioxidant Parameters of the Liver
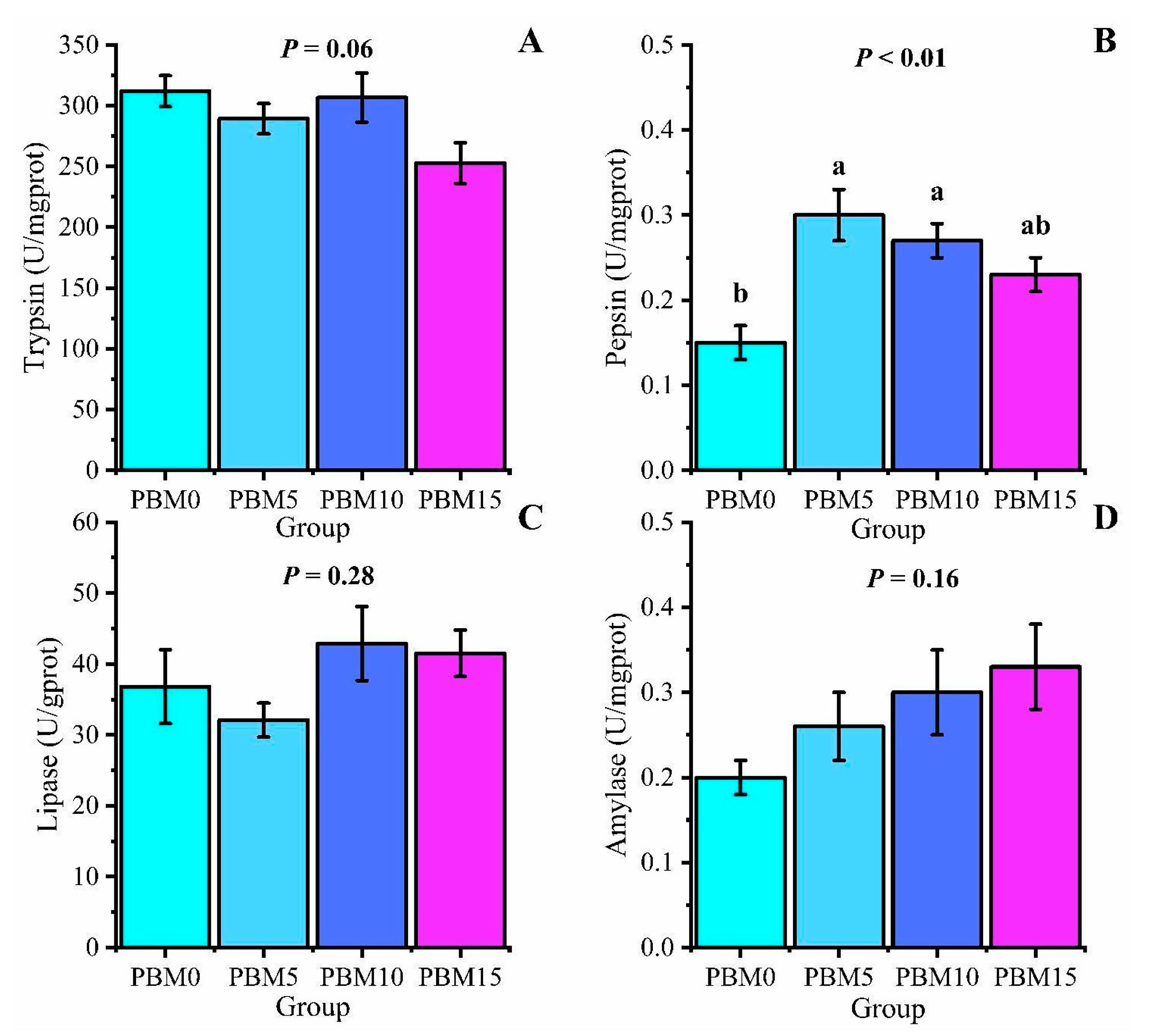
3.5. Intestinal Digestive Enzyme Activity
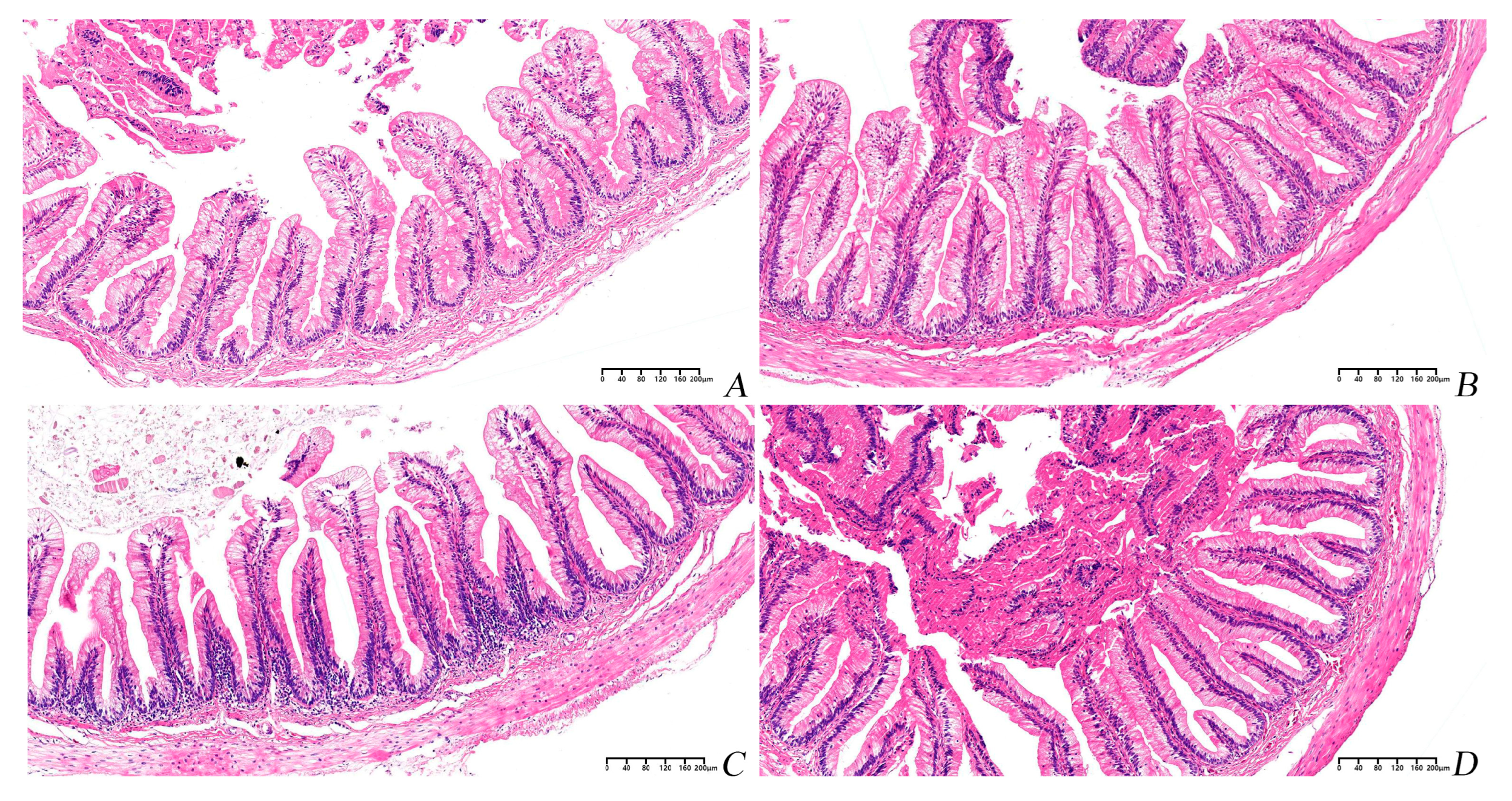
3.6. Histological Study of the Intestine
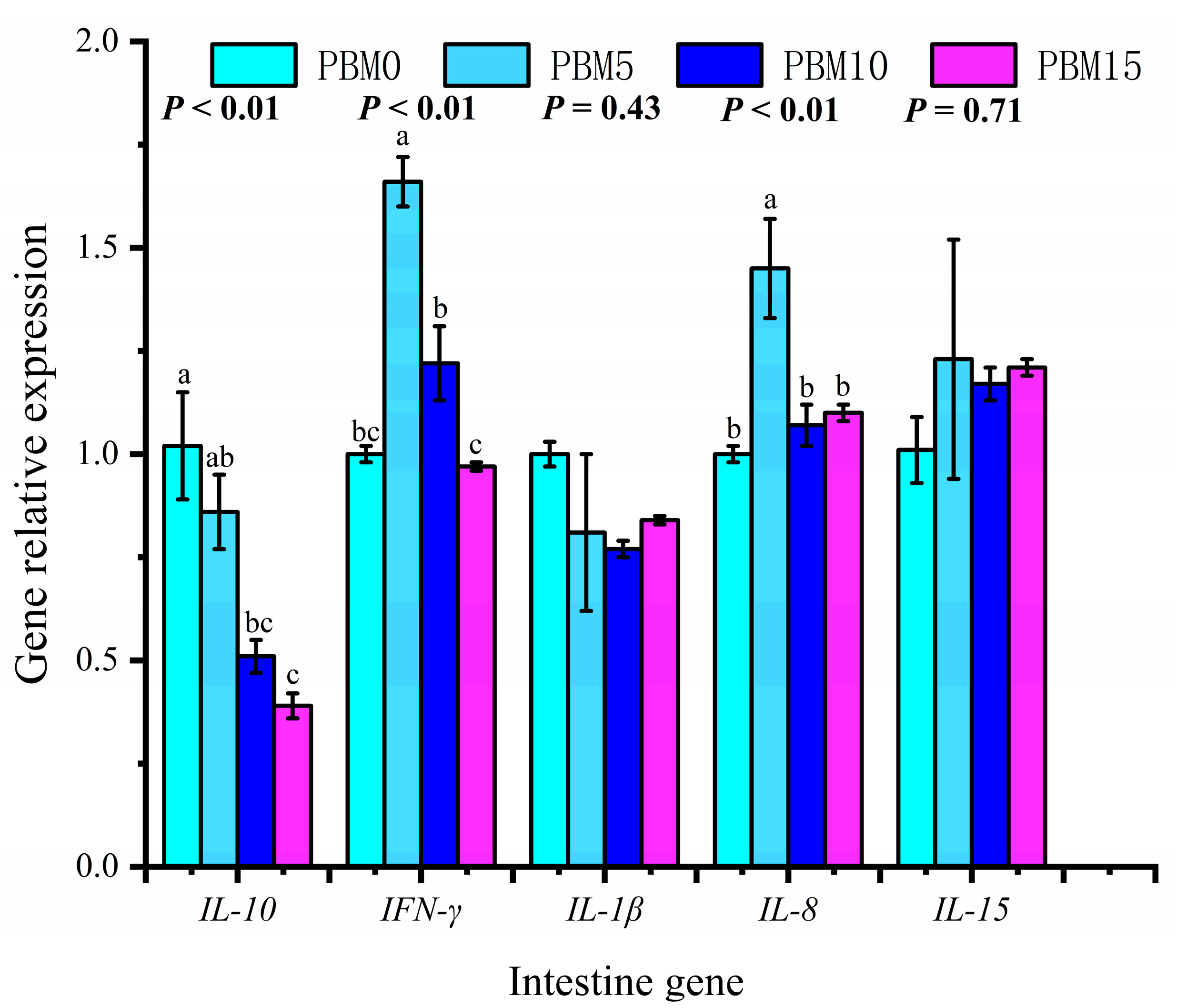
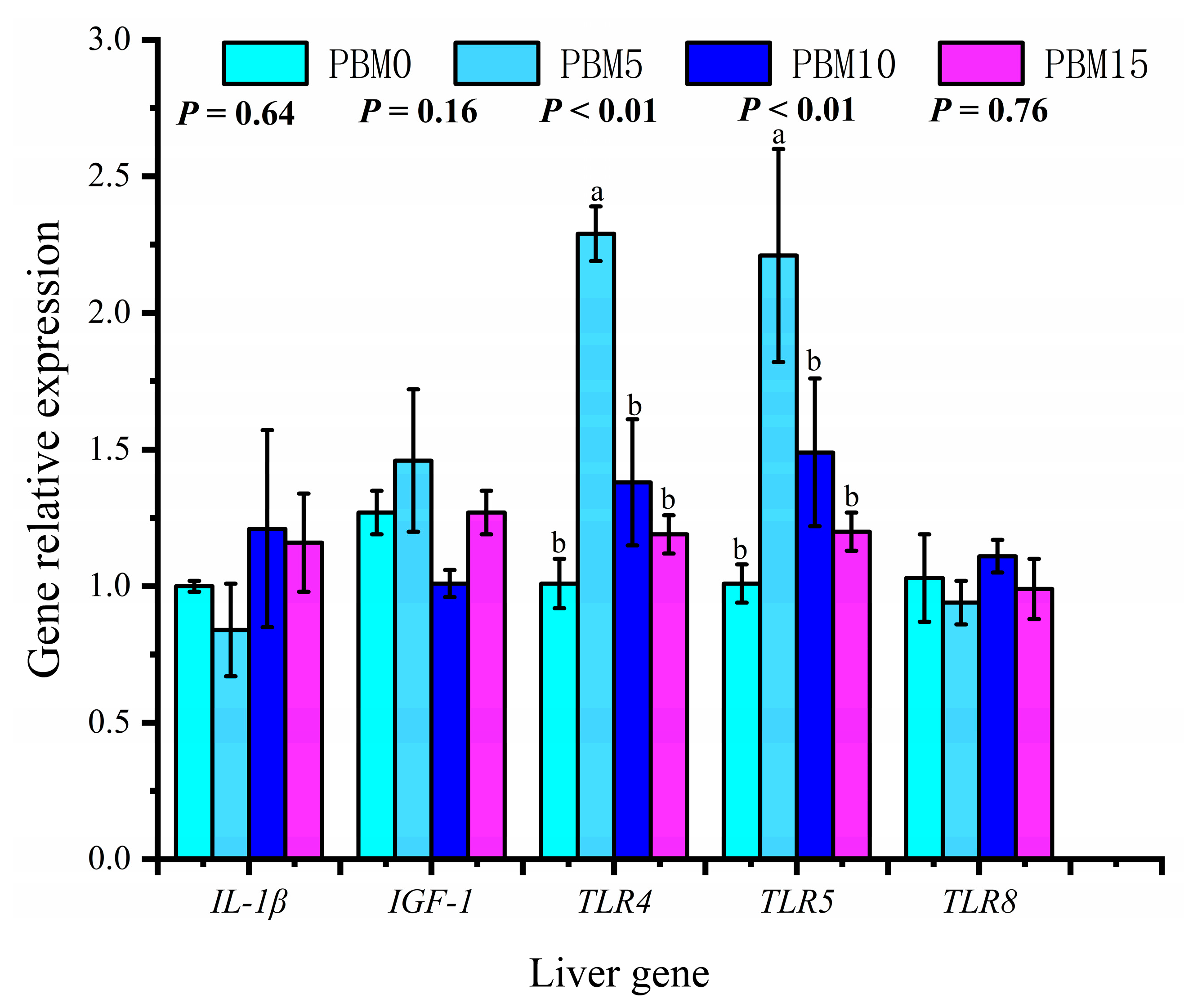
3.7. Expression of Intestinal and Liver Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cerqueira, M.; Schrama, D.; Silva, T.S.; Colen, R.; Engrola, S.A.D.; Conceição, L.E.C.; Rodrigues, P.M.L.; Farinha, A.P. How tryptophan levels in plant-based aquafeeds affect fish physiology, metabolism and proteome. J. Proteom. 2020, 221, 103782. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Yao, W.; Ye, B.; Wu, X.; Li, X.; Dong, Y. Effects of replacing fishmeal protein with poultry by-product meal protein and soybean meal protein on growth, feed intake, feed utilization, gut and liver histology of hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂) juveniles. Aquaculture 2020, 516, 734503. [Google Scholar] [CrossRef]
- Ma, Y.; Li, M.; Xie, D.; Chen, S.; Dong, Y.; Wang, M.; Zhang, G.; Zhang, M.; Chen, H.; Ye, R.; et al. Fishmeal can be replaced with a high proportion of terrestrial protein in the diet of the carnivorous marine teleost (Trachinotus ovatus). Aquaculture 2020, 519, 734910. [Google Scholar] [CrossRef]
- Attivi, K.; Mlaga, K.G.; Agboka, K.; Tona, K.; Kouame, Y.; Lin, H. Effect of fish meal replacement by black Soldier Fly (Hermetia illucens) larvae meal on serum biochemical indices, thyroid hormone and zootechnical performance of laying chickens. J. Appl. Poult. Res. 2022, 31, 100275. [Google Scholar] [CrossRef]
- Kim, J.; Baek, S.I.; Cho, S.H.; Kim, T. Evaluating the efficacy of partially substituting fish meal with unfermented tuna by-product meal in diets on the growth, feed utilization, chemical composition and non-specific immune responses of olive flounder (Paralichthys olivaceus). Aquacult. Rep. 2022, 24, 101150. [Google Scholar] [CrossRef]
- Diógenes, A.F.; Castro, C.; Miranda, A.C.; Oliva-Teles, A.; Peres, H. Dietary replacement of fishmeal by corn distillers dried grains with solubles (DDGS) in diets for turbot (Scophthalmus maximus, Linneaus, 1758) Juveniles. Aquaculture 2018, 492, 113–122. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, L.; Brown, P.B.; Feng, Y.; Zhu, Z.; Lei, A.; Zhang, Y.; Ding, X.; Bai, Y.; Jia, F. Effect of replacement of fish meal with cricket meal on growth performance, proximate composition, digestive enzyme activities, serum biochemical indices, and antioxidant capacity in largemouth bass (Micropterus salmoides). Aquac. Res. 2022, 53, 5354–5364. [Google Scholar] [CrossRef]
- Kim, K.D.; Jang, J.W.; Lee, K.W.; Kim, K.W.; Lee, B.J.; Hur, S.W.; Han, H.S. The effect of a long-term dietary replacement of fishmeal with a mixture of alternative protein sources in a formulated diet on growth performance, body composition and economic efficiency of young Korean rockfish (Sebastes schlegeli). Aquac. Res. 2021, 52, 2004–2013. [Google Scholar] [CrossRef]
- Bao, P.; Li, X.; Xu, Y. An Evaluation on the Ratio of Plant to Animal Protein in the Diet of Juvenile Sea Cucumber (Apostichopus japonicus): Growth, Nutrient Digestibility and Nonspecific Immunity. J. Ocean Univ. China 2018, 17, 1479–1486. [Google Scholar] [CrossRef]
- Yu, X.; Luo, K.; Rao, W.; Chen, P.; Lei, K.; Liu, C.; Cui, Z.; Zhang, W.; Mai, K. Effects of replacing dietary fish meal with enzyme-treated soybean meal on growth performance, intestinal microbiota, immunity and mTOR pathway in abalone Haliotis discus hannai. Fish Shellfish. Immun. 2022, 130, 9–21. [Google Scholar] [CrossRef]
- Fries, E.M.; Oxford, J.H.; Godoy, A.C.; Hassamer, M.Z.; Correia, A.F.; Boscolo, W.R.; Signor, A. Phytase on the digestibility of plant protein feed for silver catfish, Rhamdia voulezi. Aquaculture 2020, 528, 735528. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, J.; Bureau, D.P.; Cui, Z. Replacement of fish meal by rendered animal protein ingredients in feeds for cuneate drum (Nibea miichthioides). Aquaculture 2006, 252, 476–483. [Google Scholar] [CrossRef]
- McLean, E.; Barrows, F.T.; Craig, S.R.; Alfrey, K.; Tran, L. Complete replacement of fishmeal by soybean and poultry meals in Pacific whiteleg shrimp feeds: Growth and tolerance to EMS/AHPND and WSSV challenge. Aquaculture 2020, 527, 735383. [Google Scholar] [CrossRef]
- Abdul-Halim, H.H.; Aliyu-Paiko, M.; Hashim, R. Partial Replacement of Fish Meal with Poultry By-product Meal in Diets for Snakehead, Channa striata (Bloch, 1793), Fingerlings. J. World Aquacult. Soc. 2014, 45, 233–241. [Google Scholar] [CrossRef]
- Irm, M.; Taj, S.; Jin, M.; Luo, J.; Andriamialinirina, H.J.T.; Zhou, Q. Effects of replacement of fish meal by poultry by-product meal on growth performance and gene expression involved in protein metabolism for juvenile black sea bream (Acanthoparus schlegelii). Aquaculture 2020, 528, 735544. [Google Scholar] [CrossRef]
- Palupi, E.T.; Setiawati, M.; Lumlertdacha, S.; Suprayudi, M.A. Growth performance, digestibility, and blood biochemical parameters of Nile tilapia (Oreochromis niloticus) reared in floating cages and fed poultry by-product meal. J. Appl. Aquacult. 2020, 32, 16–33. [Google Scholar] [CrossRef]
- Yigit, M.; Erdem, M.; Koshio, S.; Ergün, S.; Türker, A.; Karaali, B. Substituting fish meal with poultry by-product meal in diets for black Sea turbot Psetta maeotica. Aquacult. Nutr. 2006, 12, 340–347. [Google Scholar] [CrossRef]
- Yang, M.; Guo, X.; Chen, T.; Li, P.; Xiao, T.; Dai, Z.; Hu, Y. Effect of dietary replacement of fish meal by poultry by-product meal on the growth performance, immunity, and intestinal health of juvenile red swamp crayfish, procambarus clarkia. Fish Shellfish. Immun. 2022, 131, 381–390. [Google Scholar] [CrossRef]
- Goda, A.M.; El-Haroun, E.R.; Kabir Chowdhury, M.A. Effect of totally or partially replacing fish meal by alternative protein sources on growth of African catfish Clarias gariepinus (Burchell, 1822) reared in concrete tanks. Aquac. Res. 2007, 38, 279–287. [Google Scholar] [CrossRef]
- Rossi, W.; Davis, D.A. Replacement of fishmeal with poultry by-product meal in the diet of Florida pompano Trachinotus carolinus L. Aquaculture 2012, 338–341, 160–166. [Google Scholar] [CrossRef]
- Turker, A.; Yigit, M.; Ergun, S.; Karaali, B.; Erteken, A. Potential of poultry by-product meal as a substitute for fishmeal in diets for Black Sea turbot Scophthalmus maeoticus: Growth and nutrient utilization in winter. Isr. J. Aquac. -Bamidgeh 2005, 57, 49–61. [Google Scholar]
- Aoac, I. Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Maryland, MD, USA, 2010. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cardinaletti, G.; Di Marco, P.; Daniso, E.; Messina, M.; Donadelli, V.; Finoia, M.G.; Petochi, T.; Fava, F.; Faccenda, F.; Contò, M. Growth and Welfare of Rainbow Trout (Oncorhynchus mykiss) in Response to Graded Levels of Insect and Poultry By-Product Meals in Fishmeal-Free Diets. Animals 2022, 12, 1698. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Jung, W.; Myung, S.H.; Cho, S.H.; Kim, D.S. Substitution effects of fishmeal with tuna byproduct meal in the diet on growth, body composition, plasma chemistry and amino acid profiles of juvenile olive flounder (Paralichthys olivaceus). Aquaculture 2014, 431, 92–98. [Google Scholar] [CrossRef]
- Sayed Hassani, M.H.; Banavreh, A.; Yousefi Jourdehi, A.; Mohseni, M.; Monsef Shokri, M.; Yeganeh Rastekenari, H. The feasibility of partial replacement fish meal with poultry by-products in practical diets of juvenile great sturgeon, Huso huso: Effects on growth performance, body composition, physiometabolic indices, digestibility and digestive enzymes. Aquac. Res. 2021, 52, 3605–3616. [Google Scholar] [CrossRef]
- Wang, Z.; Qian, X.; Xie, S.; Yun, B. Changes of growth performance and plasma biochemical parameters of hybrid grouper (Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀) in response to substitution of dietary fishmeal with poultry by-product meal. Aquacult. Rep. 2020, 18, 100516. [Google Scholar] [CrossRef]
- Ji, Y.; Gu, Y.; Liu, H.; Yang, Z.; Li, C. The effects of partial replacement of white fish meal by poultry by-product meal and addition of bile acid in feed on growth, digestibility, and serum enzyme activities of the Chinese soft-shelled turtle. Fish. Sci. 2017, 83, 83–88. [Google Scholar] [CrossRef]
- Shang, R.; Man, L.; Wang, G.; Li, M.; Liu, C.; Li, L. Influences of Partial Substitution of Fish Meal with Defatted Black Soldier Fly (Hermetia illucens) Larvae Meal in Diets on Growth Performance, Biochemical Parameters, and Body Composition of Juvenile Chinese Soft-Shelled Turtles (Pelodiscus sinensis). Aquacult. Nutr. 2022, 2022, 1–10. [Google Scholar] [CrossRef]
- Yang, Y.; Xie, S.; Cui, Y.; Zhu, X.; Lei, W.; Yang, Y. Partial and total replacement of fishmeal with poultry by-product meal in diets for gibel carp, Carassius auratus gibelio Bloch. Aquac. Res. 2006, 37, 40–48. [Google Scholar] [CrossRef]
- Yones, A.; Metwalli, A.A. Effects of fish meal substitution with poultry by-product meal on growth performance, nutrients utilization and blood contents of juvenile Nile Tilapia (Oreochromis niloticus). J. Aquac. Res. Dev. 2015, 7, 1000389. [Google Scholar]
- Zhou, Q.; Zhao, J.; Li, P.; Wang, H.; Wang, L. Evaluation of poultry by-product meal in commercial diets for juvenile cobia (Rachycentron canadum). Aquaculture 2011, 322–323, 122–127. [Google Scholar] [CrossRef]
- Jiang, M.; Wen, H.; Gou, G.W.; Liu, T.L.; Lu, X.; Deng, D.F. Preliminary study to evaluate the effects of dietary bile acids on growth performance and lipid metabolism of juvenile genetically improved farmed tilapia (Oreochromis niloticus) fed plant ingredient-based diets. Aquacult. Nutr. 2018, 24, 1175–1183. [Google Scholar] [CrossRef]
- Guo, J.; Zhou, Y.; Zhao, H.; Chen, W.; Chen, Y.; Lin, S. Effect of dietary lipid level on growth, lipid metabolism and oxidative status of largemouth bass, Micropterus salmoides. Aquaculture 2019, 506, 394–400. [Google Scholar] [CrossRef]
- Tian, J.; Li, Y.; Xia, Y.; Zhang, K.; Li, Z.; Gong, W.; Yu, E.; Wang, G.; Xie, J. Dietary Creatine Reduces Lipid Accumulation by Improving Lipid Catabolism in the Herbivorous Grass Carp, Ctenopharyngodon idella. Aquacult. Nutr. 2022, 2022, 1–13. [Google Scholar] [CrossRef]
- Luo, W.; Chen, P.; Zhang, X.; Zhang, Y.; Zhang, S.; Sun, K.; He, F.; Li, L.; Zhang, N.; Xiong, Y. Effect of Adding L-carnitine to High-Fat/Low-Protein Diets of Common Carp (Cyprinus carpio) and the Mechanism of Regulation of Fat and Protein Metabolism. Aquacult. Nutr. 2022, 2022, 1–13. [Google Scholar] [CrossRef]
- Sugita, T.; Gavile, A.B.; Sumbing, J.G. Efficacy of poultry by-product meal as an effective alternative to fish meal in aquaculture feed for milkfish Chanos chanos. Jpn. Agric. Res. Q. JARQ 2020, 54, 277–284. [Google Scholar] [CrossRef]
- Li, P.; Hou, D.; Zhao, H.; Huang, W.; Peng, K.; Cao, J. Dietary pyridoxine effect on growth performance, physiological metabolic parameters, intestinal enzymatic activities and antioxidant status of juvenile yellow catfish (Pelteobagrus fulvidraco). Aquacult. Rep. 2022, 24, 101153. [Google Scholar] [CrossRef]
- Xu, Z.; Regenstein, J.M.; Xie, D.; Lu, W.; Ren, X.; Yuan, J.; Mao, L. The oxidative stress and antioxidant responses of Litopenaeus vannamei to low temperature and air exposure. Fish Shellfish. Immun. 2018, 72, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; He, S.; Tan, B.; Zhang, X.; Lin, Y.; Yang, Q. Effect of Rice Protein Meal Replacement of Fish Meal on Growth, Anti-Oxidation Capacity, and Non-Specific Immunity for Juvenile Shrimp Litopenaeus vannamei. Animals 2022, 12, 3579. [Google Scholar] [CrossRef]
- Liu, Q.; Wen, B.; Li, X.; Jiang, Y.; Liang, Z.; Zuo, R. An investigation on the effects of dietary protein level in juvenile Chinese mitten crab (Eriocheir sinensis) reared at three salinities: Survival, growth performance, digestive enzyme activities, antioxidant capacity and body composition. Aquac. Res. 2021, 52, 2580–2592. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, W.; Dan, Z.; Cao, X.; Cui, K.; Zhu, S.; Zhuang, Y.; Mai, K.; Ai, Q. Effects of fish meal replaced by methanotroph bacteria meal (Methylococcus capsulatus) on growth, body composition, antioxidant capacity, amino acids transporters and protein metabolism of turbot juveniles (Scophthalmus maximus L.). Aquaculture 2023, 562, 738782. [Google Scholar] [CrossRef]
- Yuan, X.; Liu, W.; Wang, C.; Huang, Y.; Dai, Y.; Cheng, H.; Jiang, G. Evaluation of antioxidant capacity and immunomodulatory effects of cottonseed meal protein hydrolysate and its derivative peptides for hepatocytes of blunt snout bream (Megalobrama amblycephala). Fish Shellfish. Immun. 2020, 98, 10–18. [Google Scholar] [CrossRef]
- Lu, F.; Haga, Y.; Satoh, S. Effects of replacing fish meal with rendered animal protein and plant protein sources on growth response, biological indices, and amino acid availability for rainbow trout Oncorhynchus mykiss. Fish. Sci. 2015, 81, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Zambonino Infante, J.L.; Cahu, C.L.; Peres, A. Partial substitution of di-and tripeptides for native proteins in sea bass diet improves Dicentrarchus labrax larval development. J. Nutr. 1997, 127, 608–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Dong, Y.; Song, K.; Wang, L.; Li, X.; Lu, K.; Tan, B.; Zhang, C. Substituting Fish Meal with a Bacteria Protein (Clostridium autoethanogenum Protein) Derived from Industrial-Scale Gas Fermentation: Effects on Growth and Gut Health of Juvenile Large Yellow Croakers (Larimichthys crocea). Fishes 2022, 7, 228. [Google Scholar] [CrossRef]
- Xuquan, X.; Weilan, Z.; Ruixue, D.; Jie, M.; Zhuojun, W.; Bin, L.; Haoming, Y.; Yuantu, Y.; Zhijun, H. Study of enzyme-hydrolyzed soybean replacing fish meal and/or chicken meal on the growth of channel catfish (Ictalurus punctatus). Aquacult. Rep. 2022, 27, 101344. [Google Scholar] [CrossRef]
- Liu, Y.; Chang, H.; Han, D.; Lu, S.; Lv, W.; Guo, K.; Wang, C.; Li, S.; Han, S.; Liu, H. Effects of dietary chrysophyte (Poterioochromonas malhamensis) rich in beta-glucan on the growth performance, intestinal health, lipid metabolism, immune gene expression, and disease resistance against Aeromonas salmonicida in juvenile rainbow trout (Oncorhynchus mykiss). Aquaculture 2022, 561, 738589. [Google Scholar]
- Meng, X.; Li, H.; Yang, G.; Zhang, Y.; Cao, H.; Hu, W.; Zhang, X.; Zhang, J.; Li, K.; Chang, X. Safflower Promotes the Immune Functions of the Common Carp (Cyprinus carpio L.) by Producing the Short-Chain Fatty Acids and Regulating the Intestinal Microflora. Aquacult. Nutr. 2022, 2022, 1–16. [Google Scholar] [CrossRef]
- Fehrmann-Cartes, K.; Coronado, M.; Hernández, A.J.; Allende, M.L.; Feijoo, C.G. Anti-inflammatory effects of aloe vera on soy meal-induced intestinal inflammation in zebrafish. Fish Shellfish. Immun. 2019, 95, 564–573. [Google Scholar] [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Cardinaletti, G.; Cerri, R.; Giorgini, E.; Belloni, A.; Contò, M.; Tibaldi, E.; Olivotto, I. Hermetia illucens and poultry by-product meals as alternatives to plant protein sources in gilthead seabream (Sparus aurata) diet: A multidisciplinary study on fish gut status. Animals 2021, 11, 677. [Google Scholar] [CrossRef]
- Zhou, Z.; Lin, Z.; Pang, X.; Shan, P.; Wang, J. MicroRNA regulation of Toll-like receptor signaling pathways in teleost fish. Fish Shellfish. Immun. 2018, 75, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Y.; Guo, H.; Ouyang, K.; Wang, X.; Li, D.; Li, L. Nano-TiO2 aggravates immunotoxic effects of chronic ammonia stress in zebrafish (Danio rerio) intestine. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2023, 266, 109548. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | IBW (g) | FBW (g) | SR (%) | WG (%) | SGR (%/d) | FCR |
|---|---|---|---|---|---|---|
| PBM0 | 3.47 ± 0.00 | 31.89 ± 0.06 b | 91.67 ± 2.15 | 818.93 ± 1.27 b | 3.96 ± 0.00 b | 0.81 ± 0.02 ab |
| PBM5 | 3.48 ± 0.00 | 34.10 ± 0.48 ab | 92.50 ± 2.85 | 880.71 ± 13.90 ab | 4.08 ± 0.03 ab | 0.80 ± 0.01 b |
| PBM10 | 3.47 ± 0.00 | 34.93 ± 1.30 a | 91.67 ± 3.19 | 906.13 ± 37.20 a | 4.12 ± 0.07 a | 0.77 ± 0.03 b |
| PBM15 | 3.48 ± 0.00 | 31.81 ± 0.30 b | 92.50 ± 2.85 | 813.99 ± 8.92 b | 3.95 ± 0.02 b | 0.88 ± 0.02 a |
| p−Value | 0.31 | 0.02 | 0.99 | 0.02 | 0.02 | 0.02 |
| Groups | Moisture (%) | Crude Protein (%) | Crude Lipid (%) | Ash (%) |
|---|---|---|---|---|
| PBM0 | 69.46 ± 0.76 bc | 19.59 ± 0.58 ab | 4.24 ± 0.15 a | 5.24 ± 0.08 a |
| PBM5 | 71.39 ± 0.61 ab | 18.25 ± 0.49 ab | 3.54 ± 0.10 b | 4.80 ± 0.09 ab |
| PBM10 | 68.67 ± 0.69 c | 20.46 ± 0.60 a | 4.16 ± 0.17 a | 5.10 ± 0.07 a |
| PBM15 | 73.90 ± 0.43 a | 17.55 ± 0.51 b | 3.42 ± 0.10 b | 4.53 ± 0.20 b |
| p−Value | <0.01 | 0.01 | <0.01 | <0.01 |
| Groups | Villus Height (μm) | Villus Width (μm) | Muscle Layer Thickness (μm) |
|---|---|---|---|
| PBM0 | 373.62 ± 9.11 | 109.21 ± 4.87 | 72.98 ± 2.58 ab |
| PBM5 | 414.65 ± 18.98 | 111.64 ± 6.17 | 80.39 ± 8.99 a |
| PBM10 | 409.46 ± 16.65 | 110.40 ± 6.62 | 71.84 ± 5.36 ab |
| PBM15 | 354.38 ± 19.19 | 97.52 ± 6.95 | 55.17 ± 2.92 b |
| p−Value | 0.05 | 0.37 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Z.; Xu, Q.; Xie, D.; Zhao, J.; Yamamoto, F.Y.; Xu, H.; Zhao, J. Effects of the Replacement of Dietary Fish Meal with Poultry By-Product Meal on Growth and Intestinal Health of Chinese Soft-Shelled Turtle (Pelodiscus sinensis). Animals 2023, 13, 865. https://doi.org/10.3390/ani13050865
Qiu Z, Xu Q, Xie D, Zhao J, Yamamoto FY, Xu H, Zhao J. Effects of the Replacement of Dietary Fish Meal with Poultry By-Product Meal on Growth and Intestinal Health of Chinese Soft-Shelled Turtle (Pelodiscus sinensis). Animals. 2023; 13(5):865. https://doi.org/10.3390/ani13050865
Chicago/Turabian StyleQiu, Zongsheng, Qiyou Xu, Dazhang Xie, Jiantao Zhao, Fernando Y. Yamamoto, Hong Xu, and Jianhua Zhao. 2023. "Effects of the Replacement of Dietary Fish Meal with Poultry By-Product Meal on Growth and Intestinal Health of Chinese Soft-Shelled Turtle (Pelodiscus sinensis)" Animals 13, no. 5: 865. https://doi.org/10.3390/ani13050865