The two treatments showed significant differences in terms of chemical composition (
Table 1). In particular, while sucrose was not statistically different across samples, glucose and fructose were higher in cane compared to beet molasses (glucose, 4.57 vs. 0.07%, and fructose, 7.65 vs. 0.15%, on average for cane and beet, respectively;
p < 0.01). Raffinose showed opposite results, being higher on average in beet compared to cane (0.89 vs. 0.03% in beet and cane, respectively;
p < 0.01). Even other components resulted differently: starch was higher in cane compared to beet, while crude protein was the opposite. Interestingly, sulfates, phosphates, and nitrates were also higher in cane compared to beet molasses (
p < 0.05).
3.1. VFAs
The total VFA production is reported in
Table 2, expressed as mmol/L.
Net average values were similar among groups after up to 3 h of in vitro fermentation (T3), while for T4, T6, T8, and T24, VFAs produced in the beet or cane group were significantly higher than in the CTR (beet: 13.7, 23.1, and 33.0 mmol/L; cane: 13.6, 24.3, and 34.0 mmol/L; CTR: 8.7, 9.4, and 24.8 mmol/L at T6, T8, and T24, respectively;
p < 0.01). In addition to a higher concentration of VFAs produced at longer time points, the mol % composition also showed significant differences among treatments and for the treatment x time interaction (
Table 3). Acetic acid was higher in CTR than beet and cane (beet: 61.8, 63.5, and 58.2%; cane: 59.2, 60.0, and 57.0%; CTR: 70.7, 71.8, and 73.5% at T1, T4, and T24, respectively;
p < 0.01), while propionic acid significantly increased in treatments with molasses addition compared to CTR (beet: 23.2, 22.2, and 19.6%; cane: 21.3, 21.4, and 18.6%; CTR: 17.8, 17.5, and 14.2% at T1, T4, and T24, respectively;
p < 0.01). Butyric acid also displayed higher values in the molasses groups, cane in particular (beet: 12.5, 11.7, and 21.9%; cane: 15.7, 15.7, and 23.2%; CTR: 8.4, 8.2, and 8.6% at T1, T4, and T24, respectively;
p < 0.01). Moreover, a tendency was observed considering T24 against all the other time points. In particular, T24 values were lower for both acetic and propionic acid and higher for butyric acid. Iso-butyric, iso-valeric, and valeric showed no differences among treatments, remaining similar even at different time points.
3.2. Rumen Microbiota Composition
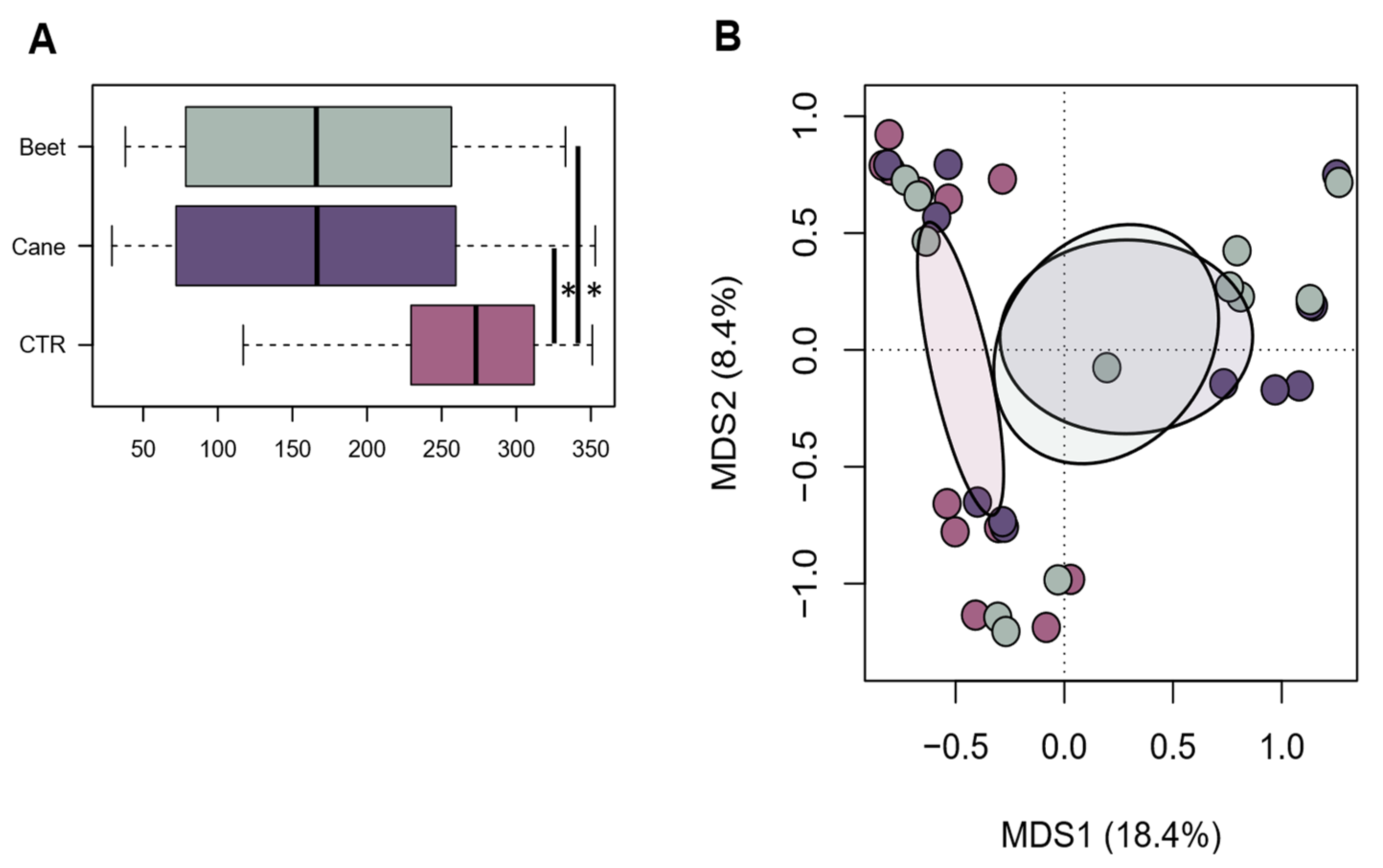
The 16S rRNA amplicon sequencing of 36 samples yielded a total of 1,440,209 high-quality reads (mean ± SD, 40,006 ± 20,920; range, 8136–67,785). Good coverage ranged from 97.3% to 100%, indicating that nearly the full extent of bacterial diversity was captured. Alpha diversity was significantly higher in CTR than in beet or cane molasses (
p < 0.05) (
Figure 1A). Similarly, CTR was significantly segregated from other groups in the PCoA based on unweighted UniFrac distances (
p = 0.01) (
Figure 1B), suggesting that compositional differences were mainly related to non-abundant members of the microbiota. Taxonomic analysis showed several compositional differences between beet molasses, cane molasses, and CTR in rumen bacteria family composition. The most abundant family was Prevotellaceae, as expected, which had a higher relative abundance in CTR (49.68%) compared to beet and cane molasses (37.13% and 28.88%, respectively;
p < 0.01). On the other hand, Ruminococcaceae, another important family in the rumen (about 13% on average in our dataset), showed no significant differences among treatments and over time. As for other families, the Lachnospiraceae family was higher in cows fed with beet molasses compared to cane molasses and CTR (11.08%, 9.05%, and 9.12%, respectively;
p < 0.05). Instead, the Streptococcaceae family had a higher relative abundance both in beet and cane compared to CTR (19,62%, 28.10%, and 6.23%, respectively;
p < 0.01). Additionally, the Veillonellaceae family was higher in both types: beet and cane molasses had average relative abundances of 6.48% and 8.67%, respectively, while the CTR result was lower (4.54%;
p < 0.05). Pseudomonadaceae had a lower relative abundance in beet and cane molasses as well (0.06% and 0.01%, respectively;
p < 0.05) compared to the CTR group (0.13%). As shown in
Table 4, Succinivibrionaceae showed a difference between groups: it has a lower relative abundance in beet molasses (0.71%) and a higher relative abundance (2.02%) in cane molasses compared to CTR (1.28%;
p < 0.05). An important cellulolytic family is Fibrobacteriaceae, which was higher in both beet and cane molasses (0.90% and 0.88%, respectively;
p < 0.05) compared to CTR. The lactic acid producer, Bifidobacteriaceae, was higher in beet but not in cane or CTR (0.49%, 0.16%, and 0.13%, respectively;
p < 0.05). An important observation was the lower relative abundance of methane producer Methanobacteriaceae family in molasses treatment (0.26% and 0.28% compared to 0.43% of CTR;
p < 0.01).
Similar differences were observed at the genus level (
Table 5). Among the numerous genera identified with the sequencing process, several of them were of interest and showed statistical differences between treatments. The relative abundance of
Metanobrevibacter was higher in CTR compared to beet and cane molasses (0.29, 0.15, 0.19%, for CTR, beet, and cane, respectively;
p < 0.01). The most represented genus was
Prevotella1, which showed differences among treatments, being higher in CTR compared to the other two (34.7, 25.5, and 16.8% in CTR, beet, and cane, respectively;
p < 0.05). Genus
Streptococcus acted differently, with higher relative abundances in beet and cane compared to CTR (16.2, 26.3, and 5.4% in beet, cane, and CTR, respectively;
p < 0.01), with the highest values in cane. A similar pattern resulted for the genera
Butyrivibrio (2.99, 2.08, and 1.63% in beet, cane, and CTR, respectively;
p < 0.01) and
Selenomonas (1.26, 1.01, and 0.36% in beet, cane, and CTR, respectively;
p < 0.01).
Succiniclasticum had a higher relative abundance in CTR (3.53, 2.81, and 5.94% in beet, cane, and CTR, respectively;
p < 0.01), as did genus
Ruminococcus (4.77, 4.51, and 7.38% in beet, cane, and CTR, respectively;
p < 0.01).
The Pearson’s coefficients calculated to evaluate possible correlations among bacteria families and VFA compositions showed interesting results (
Table 6). Acetic acid had a positive correlation with the genera
Ruminococcus (0.52,
p < 0.01),
Succiniclasticum (0.50,
p < 0.01), and
Fibrobacter (0.36), while it showed a negative correlation with
Streptococcus (−0.47,
p < 0.01),
Clostridium (−0.31), and
Butyrivibrio (−0.28). Propionic acid was positively correlated with the Prevotellaceae and Veilonellaceae families (0.41 and 0.31, respectively) and the genus
Selenomonas (0.34), while it was negatively correlated with the genera
Pseudomonas (−0.41),
Ruminococcus (−0.33), and
Acinetobacter (−0.47). Iso-butyric showed positive correlations with the genera
Pseudomonas (0.37),
Succinivibrio (0.37), and
Acinetobacter (0.28) and negative ones with
Butyrivibrio (−0.44,
p < 0.01),
Streptococcus (−0.37), and
Selenomonas (−0.29). On the contrary, both
Butyrivibrio and
Streptococcus genera were positively correlated with butyric acid (0.56 and 0.48, respectively,
p < 0.01), along with
Clostridium (0.36), while
Prevotella1 (−0.53,
p < 0.01),
Desulfovibrio (−0.50), and
Ruminococcus (−0.44) were negatively correlated. Regarding Iso-valeric acid, a positive correlation was observed with
Pseudomonas (0.42),
Sphaerochaeta (0.34), and
Mogibacterium (0.34), while a negative one was observed with
Selenomonas (−0.47),
Streptococcus (−0.35), and
Anaerovibrio (−0.28). The last evaluated VFA was valeric acid, which showed a positive correlation with the genera
Streptococcus (0.31),
Pseudomonas (0.26), and
Clostridium (0.25) and a negative one with
Succiniclasticum (−0.52,
p < 0.01),
Treponema (−0.48,
p < 0.01), and
Ruminococcus (−0.42,
p < 0.01).


 ,
, 


{kind=link}