

Effects of Different Dietary Protein Level on Growth Performance, Rumen Fermentation Characteristics and Plasma Metabolomics Profile of Growing Yak in the Cold Season
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethic
2.2. Study Location
2.3. Animals, Experimental Design and Diets
2.4. Growth Performance
2.5. Nutrients’ Apparent Digestibility
2.6. Plasma Parameters
2.7. Plasma Metabolomics Profile
2.8. Rumen Fermentation Characteristics
2.9. Statistical Analyses
3. Results
3.1. Effects of the Dietary Protein Level on the Growth Performance of Growing Yaks in the Cold Season
3.2. Effects of the Dietary Protein Level on the Nutrients’ Apparent Digestibility of Growing Yaks in the Cold Season
3.3. Effects of the Dietary Protein Level on the Plasma Biochemical indexes of Growing Yaks in the Cold Season
3.4. Effects of the Dietary Protein Level on the Plasma Antioxidant Indexes of Growing Yaks in the Cold Season
3.5. Effects of the Dietary Protein Level on the Plasma Cytokine and Immunoglobulin Content of Growing Yaks in the Cold Season
3.6. Effects of the Dietary Protein Level on the Rumen Fermentation Characteristics of Growing Yaks in the Cold Season
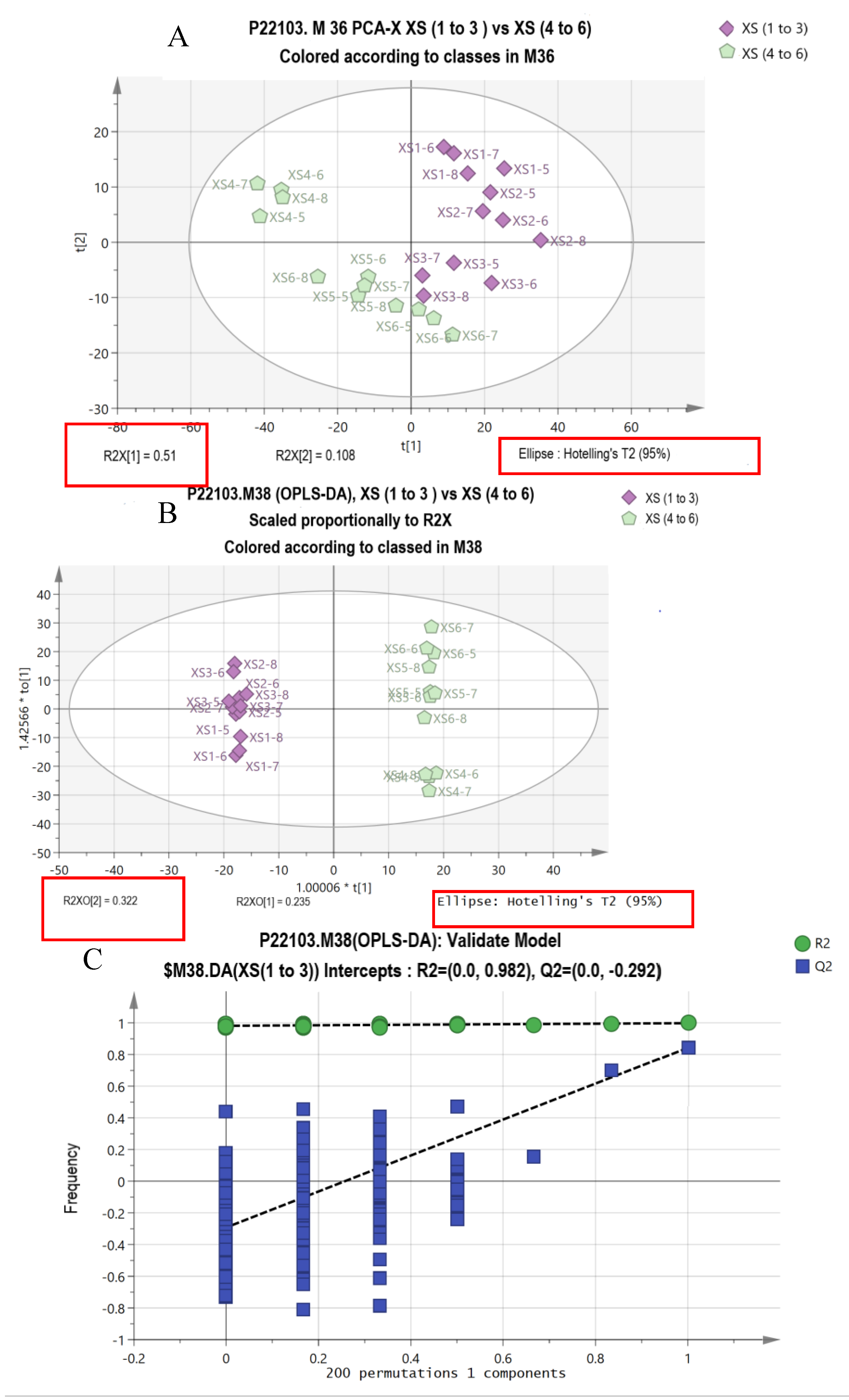
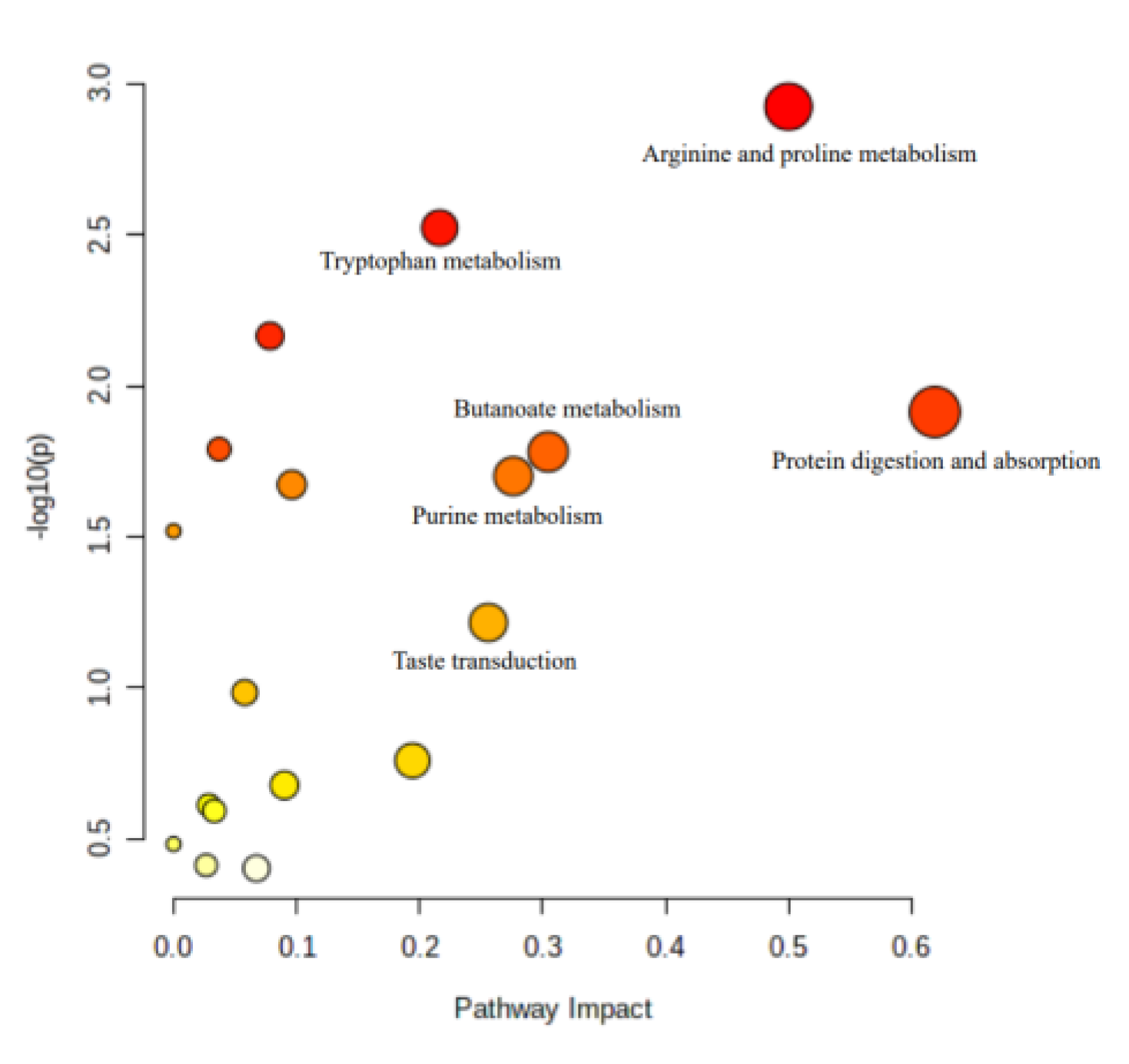
3.7. Multivariate and KEGG Analysis
4. Discussion
4.1. Growth Performance and Total Tract Digestibility
4.2. Plasma Parameters
4.3. Rumen Fermentation Characteristics
4.4. Metabolic Pathway
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jing, X.; Ding, L.; Zhou, J.; Huang, X.; Degen, A.; Long, R. The adaptive strategies of yaks to live in the Asian highlands. Anim. Nutr. 2022, 9, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.Y.; Guo, N.; Degen, A.A.; Niu, J.H.; Wei, H.Y.; Jing, X.P.; Ding, L.M.; Shang, Z.H.; Long, R.J. Effects of level of feed intake and season on digestibility of dietary components, efficiency of microbial protein synthesis, rumen fermentation and ruminal microbiota in yaks. Anim. Feed Sci. Technol. 2020, 259, 114359. [Google Scholar] [CrossRef]
- Yang, C.; Ahmad, A.A.; Bao, P.J.; Guo, X.; Wu, X.Y.; Liu, J.B.; Chu, M.; Liang, C.N.; Pei, J.; Long, R.J.; et al. Increasing dietary energy level improves growth performance and lipid metabolism through up-regulating lipogenic gene expression in yak (Bos grunniens). Anim. Feed Sci. Technol. 2020, 263, 114455. [Google Scholar] [CrossRef]
- Wu, S.R.; Chen, X.D.; Li, J.L.; Yang, Y.T.; Cui, Z.H.; Yao, J.H. Effect of alfalfa hay and starter feed supplementation on caecal microbiota and fermentation, growth, and health of yak calves. Animal. 2021, 15, 100019. [Google Scholar] [CrossRef]
- Liu, H.; Zhou, J.; Degen, A.; Liu, H.; Cao, X.; Hao, L.; Shang, Z.; Ran, T.; Long, R. A comparison of average daily gain, apparent digestibilities, energy balance, rumen fermentation parameters, and serum metabolites between yaks (Bos grunniens) and Qaidam cattle (Bos taurus) consuming diets differing in energy level. Anim. Nutr. 2022. [CrossRef]
- Zhou, J.W.; Zhong, C.L.; Liu, H.; Degen, A.A.; Titgemeyer, E.C.; Ding, L.M.; Shang, Z.H.; Guo, X.S.; Qiu, Q.; Li, Z.P.; et al. Comparison of nitrogen utilization and urea kinetics between yaks (Bos grunniens) and indigenous cattle (Bos taurus). J.Anim. Sci. 2017, 95, 4600–4612. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, J.W.; Guo, X.S.; Cui, G.X.; Ding, L.M.; Wang, H.C.; Li, L.W.; Long, R.J. Influences of dietary nitrogen and non-fiber carbohydrate levels on apparent digestibility, rumen fermentation and nitrogen utilization in growing yaks fed low quality forage based-diet. Livest. Sci. 2012, 147, 139–147. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, G. Decreasing ruminal methane production through enhancing the sulfate reduction pathway. Anim. Nutr. 2022, 9, 320–326. [Google Scholar] [CrossRef]
- Xue, B.C.; Zhang, J.X.; Wang, Z.S.; Wang, L.Z.; Peng, Q.H.; Da, L.C.; Bao, S.K.; Kong, X.Y.; Xue, B. Metabolism response of grazing yak to dietary concentrate supplementation in warm season. Animal. 2021, 15, 100175. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine (NASEM). Nutrient Requirements of Beef Cattle, 8th ed.; The National Academies Press: Washington, DC, USA, 2016. [Google Scholar]
- AOAC (2005) Official Methods of Analysis. AOAC (2005) Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar]
- Chaney, A.L.; Marbach, E.P. Modified Reagents for Determination of Urea and Ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.S.; Sharma, O.P.; Dawra, R.K.; Negi, S.S. Simple Determination of Microbial Protein in Rumen Liquor. J. Dairy Sci. 1982, 65, 2170–2183. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.W.; Liu, H.; Zhong, C.L.; Degen, A.A.; Yang, G.; Zhang, Y.; Qian, J.L.; Wang, W.W.; Hao, L.Z.; Qiu, Q.; et al. Apparent digestibility, rumen fermentation, digestive enzymes and urinary purine derivatives in yaks and Qaidam cattle offered forage-concentrate diets differing in nitrogen concentration. Livest. Sci. 2018, 208, 14–21. [Google Scholar] [CrossRef]
- Wiener, G.; Hu, J.L.; Long, R.J. The yak, 2nd ed.; FAO Regional office for Asia and the Pacific: Bangkok, Thailand, 2003. [Google Scholar]
- Zinn, R.A.; Plascencia, A. Interaction of whole cottonseed and supplemental fat on digestive function in cattle. J. Anim. Sci. 1993, 71, 11–17. [Google Scholar] [CrossRef]
- Agenäs, S.; Heath, M.F.; Nixon, R.; Wilkinson, J.M.; Phillips, C.J.C. Indicators of undernutrition in cattle. Anim. Welfare. 2006, 15, 149–160. [Google Scholar] [CrossRef]
- Abaker, J.A.; Xu, T.L.; Jin, D.; Chang, G.J.; Zhang, K.; Shen, X.Z. Lipopolysaccharide derived from the digestive tract provokes oxidative stress in the liver of dairy cows fed a high-grain diet. J. Dairy Sci. 2017, 100, 666–678. [Google Scholar] [CrossRef]
- Ma, J.; Shah, A.M.; Wang, Z.S.; Hu, R.; Zou, H.W.; Wang, X.Y.; Zhao, S.N.; Kong, X.Y. Dietary supplementation with glutamine improves gastrointestinal barrier function and promotes compensatory growth of growth-retarded yaks. Animal. 2021, 15, 100108. [Google Scholar] [CrossRef]
- Shen, X.Y. Sulfur-Induced Copper Deficiency in the Yaks. Agri. Sci. in China. 2009, 8, 1000–1013. [Google Scholar] [CrossRef]
- Costa-Roura, S.; Balcells, J.; de la Fuente, G.; Mora-Gil, J.; Llanes, N.; Villalba, D. Effects of protein restriction on performance, ruminal fermentation and microbial community in Holstein bulls fed high-concentrate diets. Anim. Feed Sci. Technol. 2020, 264, 14479. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, S.H.; Oh, S.J.; Lee, H.S.; Ji, M.; Choi, S.; Lee, S.S.; Paik, M.J. Metabolomic analysis of organic acids, amino acids, and fatty acids in plasma of Hanwoo beef on a high-protein diet. Metabolomics. 2020, 16, 114. [Google Scholar] [CrossRef]
- Ephraim, E.; Cochrane, C.Y.; Jewell, D.E. Varying Protein Levels Influence Metabolomics and the Gut Microbiome in Healthy Adult Dogs. Toxins. 2020, 12, 517. [Google Scholar] [CrossRef] [PubMed]
- Cruickshank-Quinn, C.; Jacobson, S.; Hughes, G.; Powell, R.; Petrache, I.; Kechris, K.; Bowler, R.; Reisdorph, N. Metabolomics and transcriptomics pathway approach reveals outcome-specific perturbations in COPD. Sci. Rep.-UK 2018, 8, 17132. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.W.; Feng, Q.Q.; Wang, J.; Wu, S.G.; Qi, G.H.; Zhang, H.J. Cyst(e)ine fortification in low crude protein diet improves growth performance of broilers by modulating serum metabolite profile. J. Proteomics. 2021, 238, 104154. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Wang, Z.; Li, H.; Cai, L.; Pan, J.; He, H.; Wu, Q.; Tang, Y.Z.; Ma, J.P.; Yang, L. l-Arginine induces antioxidant response to prevent oxidative stress via stimulation of glutathione synthesis and activation of Nrf2 pathway. Food Chem. Toxicol. 2018, 115, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Wei, Y.; Liu, D.; Hu, Y.; Lu, Q.; Ang, E.L.; Zhao, H.M.; Zhang, Y. An extended bacterial reductive pyrimidine degradation pathway that enables nitrogen release from β-alanine. J. Biol. Chem. 2019, 294, 15662–15671. [Google Scholar] [CrossRef]
- Tas, B.M.; Susenbeth, A. Urinary purine derivates excretion as an indicator of in vivo microbial N flow in cattle: A review. Livest. Sci. 2007, 111, 81–192. [Google Scholar] [CrossRef]
- Sun, F.; Aguerre, M.J.; Wattiaux, M.A. Starch and dextrose at 2 levels of rumen-degradable protein in iso-nitrogenous diets: Effects on lactation performance, ruminal measurements, methane emission, digestibility, and nitrogen balance of dairy cows. J. Dairy Sci. 2019, 102, 1281–1293. [Google Scholar] [CrossRef]
- Saleem, F.; Ametaj, B.N.; Bouatra, S.; Mandal, R.; Zebeli, Q.; Dunn, S.M.; Wishart, D.S. A metabolomics approach to uncover the effects of grain diets on rumen health in dairy cows. J. Dairy Sci. 2012, 95, 6606–6623. [Google Scholar] [CrossRef]
- Ma, N.; Li, Y.; Ren, L.; Hu, L.; Xu, R.; Shen, Y.; Cao, Y.; Gao, Y.; Li, J. Effects of dietary N-carbamylglutamate supplementation on milk production performance, nutrient digestibility and blood metabolomics of lactating Holstein cows under heat stress. Anim. Feed Sci. Technol. 2021, 273, 114797. [Google Scholar] [CrossRef]
- Jin, D.; Zhao, S.G.; Zheng, N.; Bu, D.P.; Beckers, Y.; Wang, J.Q. Urea nitrogen induces changes in rumen microbial and host metabolic profiles in dairy cows. Livest. Sci. 2018, 210, 104–110. [Google Scholar] [CrossRef]
- Li, X.P.; Tan, Z.L.; Li, Z.C.; Gao, S.; Yi, K.L.; Zhou, C.S.; Tang, S.X.; Han, X.F. Metabolomic changes in the liver tissues of cows in early lactation supplemented with dietary rumen-protected glucose during the transition period. Anim. Feed Sci.Technol. 2021, 281, 115093. [Google Scholar] [CrossRef]
- Rotzoll, N.; Dunkel, A.; Hofmann, T. Quantitative studies, taste reconstitution, and omission experiments on the key taste compounds in morel mushrooms (Morchella deliciosa Fr.). J. Agric. Food Chem. 2006, 54, 2705–2711. [Google Scholar] [CrossRef] [PubMed]
- López, A.; Arroquy, J.I.; Distel, R.A. Early exposure to and subsequent beef cattle performance with saline water. Livest. Sci. 2016, 185, 68–73. [Google Scholar] [CrossRef]
- Damkjær, M.; Isaksson, G.L.; Stubbe, J.; Jensen, B.L.; Assersen, K.; Bie, P. Renal renin secretion as regulator of body fluid homeostasis. Pflüg. Arch..Eur. J. Phy. 2013, 465, 153–165. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Item | Treatment | |
|---|---|---|
| Low-Protein Group | High-Protein Group | |
| Alfalfa hay | 13.30 | 13.30 |
| Oat hay | 13.30 | 13.30 |
| Full corn silage | 13.40 | 13.40 |
| Corn | 48.00 | 43.00 |
| Rapeseed oil | 0.50 | 0.50 |
| Wheat bran | 4.00 | 3.00 |
| Soybean meal | 1.50 | 3.50 |
| Cottonseed meal | 1.50 | 3.50 |
| Rapeseed meal | 1.50 | 3.50 |
| Calcium carbonate | 1.44 | 1.43 |
| Calcium hydrogen phosphate | 0.56 | 0.57 |
| Vitamin and minerals Premix 1 | 1.00 | 1.00 |
| ME MJ/kg DM 2 | 8.58 | 8.55 |
| CP % | 9.91 | 13.89 |
| RDP % CP | 67.5 | 78.8 |
| NDF % | 26.84 | 27.21 |
| ADF % | 14.67 | 15.30 |
| Ca % | 0.83 | 0.81 |
| P % | 0.43 | 0.41 |
| Item | Treatment | p-Value | |
|---|---|---|---|
| Low-Protein Group | High-Protein Group | ||
| IBW kg | 142.3 ± 2.436 | 143.7 ± 3.044 | 0.727 |
| FBW kg | 167.2 ± 2.670 | 178.3 ± 3.407 | 0.018 |
| ADG kg | 0.44 ± 0.034 | 0.62 ± 0.045 | 0.006 |
| ADFI kg/d | 5.41 ± 0.140 | 6.60 ± 0.44 | <0.001 |
| FCR | 12.30 ± 0.588 | 10.65 ± 0.694 | 0.021 |
| Item | Treatment | p-Value | |
|---|---|---|---|
| Low-Protein Group | High-Protein Group | ||
| DM % | 64.01 ± 0.312 | 64.63 ± 0.395 | 0.237 |
| OM % | 65.99 ± 0.354 | 66.61 ± 0.406 | 0.270 |
| CP % | 63.80 ± 0.692 | 66.16 ± 0.497 | 0.015 |
| NDF % | 62.16 ± 0.609 | 63.72 ± 0.378 | 0.047 |
| ADF % | 39.65 ± 1.077 | 40.23 ± 0.463 | 0.241 |
| Item | Treatment | p-Value | |
|---|---|---|---|
| Low-Protein Group | High-Protein Group | ||
| TP g/L | 59.59 ± 0.767 | 60.40 ± 1.329 | 0.610 |
| ALB g/L | 28.56 ± 0.598 | 28.70 ± 0.518 | 0.801 |
| GLOB g/L | 31.04 ± 0.811 | 31.70 ± 0.715 | 0.551 |
| GLU mmol/L | 4.34 ± 0.110 | 4.97 ± 0.145 | 0.004 |
| TG nmol/L | 0.098 ± 0.004 | 0.102 ± 0.005 | 0.568 |
| BUN mmol/L | 2.96 ± 0.164 | 3.42 ± 0.0.104 | 0.038 |
| ALT U/L | 3.62 ± 0.101 | 2.89 ± 0.078 | <0.001 |
| AST U/L | 44.73 ± 1.217 | 39.52 ± 0.659 | 0.002 |
| ALP U/L | 41.75 ± 0.456 | 39.26 ± 0.674 | 0.148 |
| Item | Treatment | p-Value | |
|---|---|---|---|
| Low-Protein Group | High-Protein Group | ||
| T-AOC U/mL | 10.36 ± 0.365 | 10.96 ± 0.352 | 0.266 |
| SOD U/mL | 118.24 ± 3.627 | 138.64 ± 4.764 | 0.004 |
| GSH-Px U/mL | 182.45 ± 10.113 | 204.32 ± 10.496 | 0.156 |
| MDA mmol/L | 5.37 ± 0.108 | 4.66 ± 0.141 | 0.001 |
| Item | Treatment | p-Value | |
|---|---|---|---|
| Low-Protein Group | High-Protein Group | ||
| IL-4 ng/L | 73.78 ± 2.173 | 75.19 ± 1.566 | 0.609 |
| IL-10 ng/L | 98.43 ± 5.180 | 109.00 ± 3.159 | 0.105 |
| IL-2 ng/L | 49.35 ± 2.744 | 62.40 ± 3.118 | 0.007 |
| IL-6 ng/L | 91.60 ± 3.337 | 89.36 ± 2.615 | 0.605 |
| IFN-γ ng/L | 115.02 ± 5.499 | 99.27 ± 1.823 | 0.017 |
| TNF-α ng/L | 84.49 ± 1.529 | 77.85 ± 2.342 | 0.032 |
| IgA g/L | 0.64 ± 0.015 | 0.68 ± 0.026 | 0.299 |
| IgG g/L | 0.20 ± 0.007 | 0.23 ± 0.004 | 0.012 |
| IgM g/L | 0.07 ± 0.001 | 0.07 ± 0.001 | 0.787 |
| Item | Treatment | p-Value | |
|---|---|---|---|
| Low-Protein Group | High-Protein Group | ||
| pH | 6.63 ± 0.053 | 6.71 ± 0.062 | 0.084 |
| MCP g/dL | 49.53 ± 1.187 | 53.56 ± 1.425 | 0.047 |
| NH3-n g/dL | 4.60 ± 0.130 | 5.48 ± 0.105 | <0.001 |
| TVFA mmol/L | 64.43 ± 0.902 | 67.88 ± 0.642 | 0.008 |
| Acetate % | 65.74 ± 0.527 | 65.31 ± 0.458 | 0.550 |
| Propionate % | 21.05 ± 0.519 | 22.10 ± 0.347 | 0.115 |
| Butyrate % | 9.76 ± 0.320 | 8.99 ± 0.267 | 0.088 |
| Acetate:Propionate | 3.14 ± 0.093 | 2.97 ± 0.058 | 0.127 |
| Compounds | MW | RT | VIP | FC | p Value |
|---|---|---|---|---|---|
| Guanosine | 283 | 1.707 | 1.446 | 2.145 | 0.007 |
| L-Isoleucine | 131 | 1.327 | 1.34 | 2.143 | 0.008 |
| Acetone | 58 | 0.974 | 1.354 | 2.135 | 0.002 |
| Piperidine | 85 | 1.009 | 1.476 | 2.125 | 0.004 |
| 1-Benzazole | 117 | 5.908 | 1.356 | 1.982 | 0.01 |
| Beta-Alanine | 89 | 0.925 | 1.547 | 1.893 | 0.002 |
| D-Serine | 105 | 0.9 | 1.354 | 1.874 | 0.002 |
| Harmaline | 214 | 6.551 | 1.462 | 1.822 | 0.000 |
| L-Tyrosine | 181 | 1.188 | 1.469 | 1.786 | 0.002 |
| Epinephrine | 183 | 4.913 | 1.44 | 1.764 | 0.006 |
| Xanthine | 152 | 2.054 | 1.005 | 1.715 | 0.000 |
| Adenosine | 267 | 1.421 | 1.008 | 1.706 | 0.001 |
| Indole-3-ethanol | 161 | 6.061 | 1.021 | 1.698 | 0.001 |
| Spermidine | 145 | 0.907 | 1.786 | 1.684 | 0.008 |
| Spermine | 202 | 0.765 | 1.764 | 1.684 | 0.010 |
| Histamine | 111 | 0.957 | 1.457 | 1.675 | 0.006 |
| Cytidine | 243 | 0.927 | 1.576 | 1.657 | 0.008 |
| Nandrolone phenpropionate | 406 | 6.943 | 1.301 | 1.652 | 0.000 |
| Guanine | 151 | 1.711 | 1.785 | 1.567 | 0.009 |
| Kynurenic acid | 189 | 5.365 | 1.56 | 1.564 | 0.004 |
| L-Tryptophan | 204 | 4.197 | 1.167 | 1.556 | 0.009 |
| gamma-Aminobutyric Acid | 103 | 0.922 | 1.453 | 1.476 | 0.008 |
| Thymine | 126 | 1.725 | 1.678 | 1.457 | 0.003 |
| Valine | 117 | 0.956 | 1.765 | 1.456 | 0.008 |
| L-Glutamic acid | 147 | 0.942 | 1.245 | 1.367 | 0.008 |
| (R)(-)-Allantoin | 158 | 0.977 | 1.487 | 1.346 | 0.009 |
| Creatinine | 113 | 0.853 | 1.547 | 1.335 | 0.001 |
| Cyclopentanone | 84 | 4.097 | 1.357 | 0.482 | 0.007 |
| Triethanolamine | 149 | 20.11 | 1.049 | 0.459 | 0.009 |
| Adenine | 135 | 1.531 | 1.085 | 0.454 | 0.001 |
| 2’-Deoxyadenosine | 251 | 1.526 | 1.071 | 0.452 | 0.001 |
| n-Acetylserotonin | 218 | 5.162 | 1.476 | 0.432 | 0.002 |
| L-Phenylalanine | 165 | 2.546 | 1.056 | 0.431 | 0.011 |
| Benzamide | 121 | 4.879 | 1.516 | 0.43 | 0.001 |
| Hypoxanthine | 136 | 1.724 | 1.521 | 0.417 | 0.022 |
| 3-Methylcrotonylglycine | 157 | 1.096 | 1.576 | 0.407 | 0.001 |
| (R)-Acetoin | 88 | 4.834 | 1.362 | 0.405 | 0.000 |
| 3-Methyldioxyindole | 163 | 5.225 | 1.069 | 0.369 | 0.001 |
| (R)-3-((R)-3-Hydroxybutanoyloxy) butanoate | 190 | 4.734 | 1.157 | 0.363 | 0.000 |
| Stearamide | 283 | 15.914 | 1.37 | 0.349 | 0.033 |
| Norepinephrine | 169 | 4.4 | 1.763 | 0.342 | 0.001 |
| L-Methionine | 149 | 1.111 | 1.334 | 0.335 | 0.003 |
| Uracil | 112 | 1.181 | 1.557 | 0.334 | 0.004 |
| Saccharin | 183 | 4.81 | 1.299 | 0.324 | 0.000 |
| Indole-3-acetic acid | 175 | 6.027 | 1.009 | 0.303 | 0.001 |
| n-Undecanoylglycine | 243 | 13.97 | 1.415 | 0.21 | 0.034 |
| Sulcatone | 126 | 2.575 | 1.64 | 0.163 | 0.020 |
| Dicloralurea | 352 | 0.283 | 2.207 | 0.06 | 0.009 |
| Pathway Name | Metabolites |
|---|---|
| Protein digestion and absorption (11) | Valine,Piperidine,L-Tyrosine,L-Tryptophan,L-Phenylalanine,L-Methionine,L-Isoleucine,L-Glutamic acid,1-Benzazole,Histamine,Beta-Alanine |
| Arginine and proline metabolism (8) | Spermine, Spermidine, n-Carbamoylputrescine, L-Glutamic acid, gamma-Glutamyl-gamma-aminobutyraldehyde, gamma-Aminobutyric acid, D-Proline, Creatinine |
| Tryptophan metabolism (8) | Xanthurenic acid, n-Acetylserotonin, L-Tryptophan, Kynurenic acid,1-Benzazole,Indole-3-ethanol,Indole-3-acetic acid,3-Methyldioxyindole |
| Purine metabolism (8) | Xanthine,Hypoxanthine,Guanosine,Guanine,Adenosine,Adenine,2’-Deoxyadenosine,(R)(-)-Allantoin |
| Butanoate metabolism (5) | L-Glutamic acid, gamma-Aminobutyric Acid,Acetone,(R)-Acetoin,(R)-3-((R)-3-Hydroxybutanoyloxy)butanoate |
| Taste transduction (5) | Saccharin, Norepinephrine, L-Glutamic acid, gamma-Aminobutyric acid, D-Serine |
| Pyrimidine metabolism (4) | Uracil, Thymine, Cytidine, Beta-Alanine |
| Pantothenate and CoA biosynthesis (4) | Valine, Uracil, Spermine, Beta-Alanine |
| Glutathione metabolism (3) | Spermine, Spermidine, L-Glutamic acid |
| Renin secretion (3) | Norepinephrine, Epinephrine, Adenosine |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Sun, G.; Dunzhu, L.; Li, X.; Zhaxi, L.; Zhaxi, S.; Suolang; Ciyang; Yangji, C.; Wangdui, B.; et al. Effects of Different Dietary Protein Level on Growth Performance, Rumen Fermentation Characteristics and Plasma Metabolomics Profile of Growing Yak in the Cold Season. Animals 2023, 13, 367. https://doi.org/10.3390/ani13030367
Zhu Y, Sun G, Dunzhu L, Li X, Zhaxi L, Zhaxi S, Suolang, Ciyang, Yangji C, Wangdui B, et al. Effects of Different Dietary Protein Level on Growth Performance, Rumen Fermentation Characteristics and Plasma Metabolomics Profile of Growing Yak in the Cold Season. Animals. 2023; 13(3):367. https://doi.org/10.3390/ani13030367
Chicago/Turabian StyleZhu, Yanbin, Guangming Sun, Luosang Dunzhu, Xin Li, Luosang Zhaxi, Suolang Zhaxi, Suolang, Ciyang, Cidan Yangji, Basang Wangdui, and et al. 2023. "Effects of Different Dietary Protein Level on Growth Performance, Rumen Fermentation Characteristics and Plasma Metabolomics Profile of Growing Yak in the Cold Season" Animals 13, no. 3: 367. https://doi.org/10.3390/ani13030367