
Analysis of Promoter Methylation of the Bovine FOXO1 Gene and Its Effect on Proliferation and Differentiation of Myoblasts
by
 ,
,
Pengfei Shi
1,2, 
Yong Ruan
1,2,
Wenjiao Liu
1,2,
Jinkui Sun
1,2,
Jiali Xu
1 and
Houqiang Xu
1,2,* 1
Key Laboratory of Animal Genetics, Breeding and Reproduction in the Plateau Mountainous Region, Ministry of Education, Guizhou University, Guiyang 550025, China
2
College of Animal Science, Guizhou University, Guiyang 550025, China
*
Author to whom correspondence should be addressed.
Animals 2023, 13(2), 319; https://doi.org/10.3390/ani13020319
Submission received: 19 November 2022
/
Revised: 1 January 2023
/
Accepted: 12 January 2023
/
Published: 16 January 2023
(This article belongs to the Collection Advances in Cattle Breeding, Genetics and Genomics)
Abstract
:Simple Summary
The purpose of this study was to initially determine the role of methylation of the promoter region of Forkhead box O 1 (FOXO1) in regulating its transcriptional level and to further investigate the effect of FOXO1 on the proliferation and differentiation of bovine myogenic cells. In this study, we used bisulfite sequencing polymerase chain reaction, real-time quantitative PCR, western blot, cell counting kit-8 (CCK-8), and flow cytometry and found that the mRNA expression of the FOXO1 was low when the methylation of FOXO1 promoter region was high, and silencing the expression of the FOXO1 gene could promote the proliferation and differentiation of myoblasts.
Abstract
This study aimed to explore the regulatory role of FOXO1 promoter methylation on its transcriptional level and unravel the effect of FOXO1 on the proliferation and differentiation of bovine myoblasts. Bisulfite sequencing polymerase chain reaction (BSP) and real-time quantitative PCR were performed to determine the methylation status and transcript levels of the FOXO1 promoter region at different growth stages. BSP results showed that the methylation level in the calf bovine (CB) group was significantly higher than that in the adult bovine (AB) group (p < 0.05). On the other hand, qRT-PCR results indicated that the mRNA expression level in the AB group was significantly higher than that in the CB group (p < 0.05), suggesting a significant decrease in gene expression at high levels of DNA methylation. CCK-8 and flow cytometry were applied to determine the effect of silencing the FOXO1 gene on the proliferation of bovine myoblasts. Furthermore, qRT-PCR and Western blot were conducted to analyze the expression of genes associated with the proliferation and differentiation of bovine myoblasts. Results from CCK-8 revealed that the short hairpin FOXO1 (shFOXO1) group significantly promoted the proliferation of myoblasts compared to the short-hairpin negative control (shNC) group (p < 0.05). Flow cytometry results showed a significant decrease in the number of the G1 phase cells (p < 0.05) and a significant increase in the number of the S phase cells (p < 0.05) in the shFOXO1 group compared to the shNC group. In addition, the expression of key genes for myoblast proliferation (CDK2, PCNA, and CCND1) and differentiation (MYOG, MYOD, and MYHC) was significantly increased at both mRNA and protein levels (p < 0.05). In summary, this study has demonstrated that FOXO1 transcription is regulated by methylation in the promoter region and that silencing FOXO1 promotes the proliferation and differentiation of bovine myoblasts. Overall, our findings lay the foundation for further studies on the regulatory role of epigenetics in the development of bovine myoblasts.
1. Introduction
Muscle proliferation and differentiation, a highly coordinated and complex biological process, have been extensively studied at the genetic level. Studies have demonstrated that several vital genes are involved in its regulation, including the cell cyclin gene family (CCN), the cyclin-dependent kinase gene family (CDK), the proliferating cell nuclear antigen (PCNA), and myogenic regulatory factors (MRF) [1,2,3]. However, there is a need for an in-depth understanding of the regulatory mechanisms of muscle proliferation and differentiation, with the overarching goal of improving the economic benefits of meat.
The mammalian Forkhead Box O (FOXO) transcription factor family comprises five members: FOXO1, FOXO3, FOXO3b, FOXO4, and FOXO6 [4]. FOXO1, the first member to be identified, acts as an essential mammalian transcription factor that plays a regulatory role in various cells [5]. Most early studies on FOXO1 were associated with cancer therapy since it has been considered a tumor suppressor mainly due to its potential to promote apoptosis [6]. Studies have found that FOXO1 deficiency causes lethal defects in mouse embryonic angiogenesis [7,8], suggesting that it modulates cell growth and development. In addition, numerous studies have reported that FOXO1 regulates myoblasts, ovarian granulosa cells, and adipocytes, and its activation in cells can lead to cell cycle arrest and subsequent apoptosis [9,10]. Qi et al. [11] regulated FOXO1 gene expression in sheep ovarian granulosa cells and found that the gene inhibited the proliferation of sheep GCs and affected the mRNA expression abundance of genes involved in the regulation of apoptosis, cell cycle, and steroid synthesis. It has also been reported that abundant expression of FOXO1 in adipocytes regulates the expression of genes associated with adipocyte differentiation, anti-oxidative stress, and lipid metabolism and modulates the adipocyte size to adapt to calorie intake, thereby regulating energy balance [12,13]. A previous study revealed that overexpression of FOXO1 significantly inhibits the differentiation process of C2C12 myoblasts [14]. Moreover, inhibition of myogenic differentiation caused by FOXO1 was significantly restored by insulin treatment since it is one of the most important transcription factors in the insulin/insulin-like growth factor 1 (IGF-1) signaling pathway [15]. Although FOXO1 plays an inhibitory role in the early differentiation of myoblasts, it can stimulate myotube fusion in primary mouse myoblasts at the end of differentiation [9,16]. Studies have also shown that the expression of FOXO1 increases gradually during the differentiation of primary bovine myoblasts, and its polymorphism is associated with the productive performance of yellow cattle [17,18].
Mammalian DNA methylation is an important epigenetic modification that is dynamically regulated during development [19]. Its ability to modulate cell proliferation and differentiation by regulating the timing and level of gene expression, and excessive aberrant methylation can lead to cellular carcinogenesis [20,21]. Evidence suggests that DNA methylation in the promoter region negatively regulates mRNA expression [22]. The main aim of this study was to determine the role of methylation in the promoter region of FOXO1 in regulating its transcriptional level and to further explore the effect of FOXO1 on the proliferation and differentiation of bovine myoblasts. Collectively, our findings provide fundamental evidence to unravel the regulatory mechanisms of FOXO1 at the transcriptional level and its effects on the function of myoblasts.
2. Materials and Methods
2.1. Tissue Sample Collection
We randomly selected 3-day-old (calf bovine, CB) and 24-month-old (adult bovine, AB) healthy Guanling cattle under the same feeding conditions (Guanling Cattle Industrial Park, Guanling County, Anshun City, Guizhou Province, China). After slaughter by Guanling Cattle Industrial Park staff, the longissimus dorsi muscle was harvested, and the separated tissue samples were stored in liquid nitrogen for backup. The use of experimental animals in this study was approved by the Laboratory Animal Ethics of Guizhou University (No. EAE-GZU-2021-E019, Guiyang, China; 1 November 2021).
2.2. Bisulfite Sequencing Polymerase Chain Reaction
Genomic DNA was extracted from the tissue samples using the OMEGA (USA) DNA Extraction Kit according to the manufacturer’s instructions. The extracted DNA was treated with sodium bisulfite using the EZ DNA Methylation-Gold Kit (Zymo Research, Orange County, CA, USA), following the manufacturer’s instructions. Next, each unmethylated cytosine in the chemically transformed DNA was converted to uracil, whereas each methylated cytosine was left unchanged. The methylation status of the FOXO1 promoter region was subsequently analyzed by the bisulfite sequencing polymerase chain reaction (BSP). CpG islands in the FOXO1 promoter region (Gene ID: 506618) were predicted by the MethPrimer (http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi, accessed on 23 June 2021), followed by the design of BSP primers using Primer Premier 5.0 (Table 1). The treated DNA was amplified using the Hieff® Hotstart PCR Genotyping Master Mix (YEASEN, Shanghai, China) in a Bio-Rad thermal cycler. The following thermocycling conditions were applied: pre-denaturation for 5 min at 95 °C for one cycle; followed by denaturation for 30 s at 95 °C, annealing for 30 s at 51 °C, and extension for 40 s at 72 °C for 35 cycles; and final extension for 10 min at 72 °C. The BSP products were purified using an agarose gel DNA extraction and purification kit (OMEGA, Norcross, GA, USA) according to the manufacturer’s instructions and then ligated with the pMD19-T vector (TaKaRa, Dalian, China). Finally, 10 positive clones from each sample were randomly selected for sequencing (Qingke, Beijing, China).
2.3. Real-Time Quantitative PCR
Total RNA was extracted from the tissue samples and cells using the TRIzol reagent (Solarbio, Beijing, China). Next, first-strand cDNA synthesis was performed using the StarScrip II First Strand cDNA Kit (Genstar, Beijing, China) according to the manufacturer’s instructions, and the reaction products were stored at −20 °C. The qRT-PCR was then performed to analyze the relative expression of FOXO1, value-added key genes (CDK2, PCNA, and CCND1), and differentiation key genes (MYOD, MYOG, and MYHC) at the mRNA level. The details of the qRT-PCR primers are shown in Table 1. The qRT-PCR was performed with 2 × PowerUp™ SYBR™ Green Master Mix in a Bio-Rad CFX96™ (Thermo Fisher Scientific, Waltham, MA, USA) real-time detection system according to the manufacturer’s instructions. The following qRT-PCR procedure was applied: pre-denaturation for 2 min at 95 °C for one cycle, followed by denaturation for 15 s at 95 °C, annealing for 15 s at 60 °C, and extension for 30 s at 72 °C for 40 cycles. The final melting curve was created by heating in 0.5 °C steps from 60 °C to 95 °C, and the fluorescence acquisition time was 5 s. Notably, each sample was replicated three times. Relative mRNA expression was normalized to β-actin (ACTB) mRNA as an internal reference and calculated using the 2−∆∆CT quantification method [23].
2.4. Plasmid Construction
Multiple pairs of FOXO1 interfering sequences were designed following the shRNA design principles and sent to GEMA Ltd. (Shanghai, China) for synthesis and tested for interference efficiency by qRT-PCR and Western blot assay. The detailed shRNA target sequences are shown in Table 2.
2.5. Cell Culture and Transfection
Bovine myoblasts were isolated from calf longissimus dorsi muscle by enzymatic digestion, and cell purity was analyzed by indirect immunofluorescence [24,25]. The sorted myoblasts were inoculated in six-well plates using DMEM-F/12 (Gibco, San Diego, CA, USA), which contains 15% fetal bovine serum and 1% penicillin-streptomycin. When myoblasts reached 80% confluence, they were transfected with shFOXO1 and shNC using Lipofectamine 3000 (Thermo Fisher Scientific, Waltham, MA, USA) transfection reagent according to the manufacturer’s instructions. After 48 h of transient transfection, cells were harvested for subsequent analysis, including qRT-PCR, Western blot analysis to determine cell proliferation, and flow cytometry for cell cycle analysis. With regard to cell differentiation studies, the medium was changed to differentiation medium (DMEM-F/12 + 2% horse serum) 6 h after completing transient transfection to induce cell differentiation. Cells were incubated for 48 h and then harvested for qRT-PCR and Western blot analyses.
2.6. Western Blot
Total protein was extracted by treating collected cells with RIPA lysate (Solarbio, Beijing, China) containing protease inhibitors, followed by measuring the protein concentration using a BCA protein concentration assay kit (Solarbio, Beijing, China). Equal protein samples were resolved using 10% SDS-PAGE gel electrophoresis and transferred to PVDF membranes. Next, membranes were blocked with 5% skimmed milk for 2 h and incubated with primary antibodies overnight at 4 °C. The following primary antibodies were used: FOXO1 (Rabbit anti-FOXO1, ABclonal, Wuhan, China), PCNA (Rabbit anti-PCNA, Proteintech, Wuhan, China), CCND1 (Rabbit anti-CCND1, Proteintech, Wuhan, China), CDK2 (Rabbit anti-CDK2, Bioss, Beijing, China), MYOD (Rabbit anti-MYOD, Bioss, Beijing, China), MYOG (Rabbit anti-MYOG, ABclonal, Wuhan, China), MYHC (Rabbit anti-MYHC, ABclonal, Wuhan, China), and ACTB (Rabbit anti-ACTB, Proteintech, Beijing, China). On the next day, membranes were washed three times with Tris-buffered saline with Tween-20 (TBST) buffer and incubated with HRP-labelled secondary antibodies for 2 h at 37 °C. Membranes were washed three times with TBST and visualized using ECL (NeoSami, Suzhou, China) ultrasensitive luminescence. Finally, images were captured using the ChemiDocXRS system, and the bands were analyzed in greyscale using Image J software.
2.7. Cell Counting Kit-8 (CCK-8) Assay to Determine Cell Proliferation
The CCK-8 assay was performed to explore the effect of FOXO1 gene silencing on the proliferative activity of myoblasts. Briefly, myoblasts were inoculated on 96-well plates, transfected with the negative control and test groups, and then incubated at 37 °C and 5% CO2 for 12 h, 24 h, 48 h, and 72 h according to the instructions of the CCK-8 reagent (APExBIO, Houston, TX, USA). Finally, the absorbance (OD) was measured at 450 nm using an enzyme marker (Thermo Fisher Scientific, Waltham, MA, USA).
2.8. Flow Cytometry to Determine the Cell Cycle
Briefly, myoblasts were fixed overnight at 4 °C using pre-cooled 70% ethanol (7:3 ratio of ethanol to PBS) and then treated with a cell cycle assay kit (Servicebio, Wuhan, China) according to the manufacturer’s instructions. The DNA content of cells under different treatment conditions was then measured using a flow cytometer (CytoFLEX, Beckman, Brea, CA, USA). Notably, each treatment was replicated three times.
2.9. Statistical Analysis
All statistical analyses were performed using SPSS 18.0 software (IBM SPSS Statistics 18, Inc., Chicago, IL, USA), and all data are presented as means ± SD of three biological replicates and three technical replicates to ensure the accuracy of the experimental data. A one-way analysis of variance (ANOVA) was used to compare the differences between the various groups. Statistical significance was considered at p < 0.05.
3. Results
3.1. Effect of FOXO1 Promoter Methylation on Transcript Levels
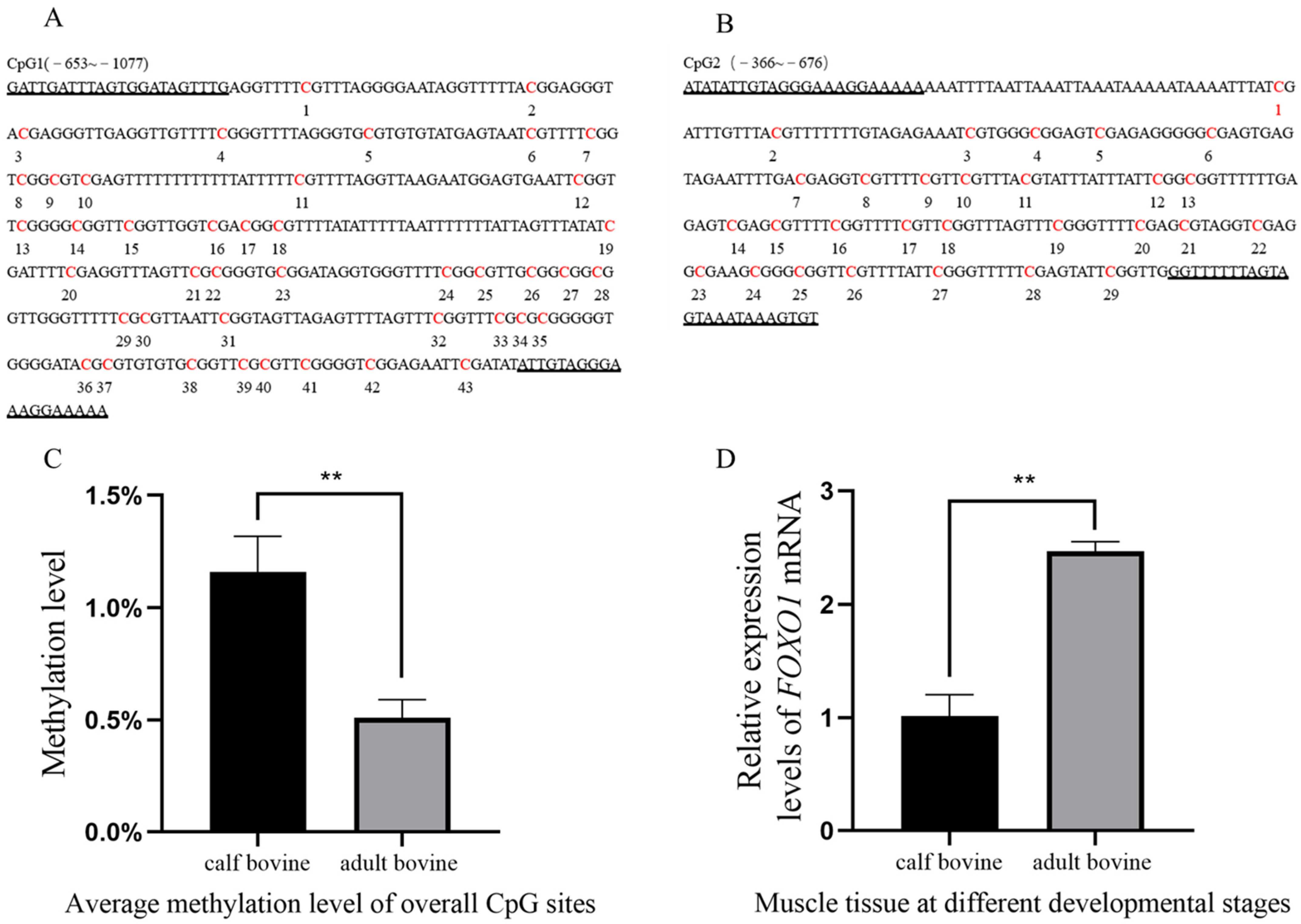
Analysis of the FOXO1 promoter region revealed the presence of a total of two CpG islands, with CpG1 (−653~−1077) and CpG2 (−366~−676) containing 43 and 29 methylation sites, respectively (Figure 1A,B). The BSP results showed that the average methylation rates were 0.972%, 1.25%, and 1.25% in the CB group and 0.556%, 0.417%, and 0.556% in the AB group, indicating that the methylation rate of the FOXO1 promoter region was significantly higher in the CB group than in the AB group (p < 0.05) (Figure 1C). To confirm whether the methylation status of the FOXO1 promoter region affected its expression level in muscle tissue, we examined the expression levels of FOXO1 mRNA in both groups. The results demonstrated that FOXO1 was expressed in both groups, but the relative expression of FOXO1 was significantly higher in the AB group compared to the CB group (p < 0.05) (Figure 1D), suggesting that high levels of DNA methylation significantly reduced gene expression. The results of the software analysis of the FOXO1 promoter region and details of the methylation rate of each CpG site are shown in Figure S1.
3.2. Cell Purity Assay
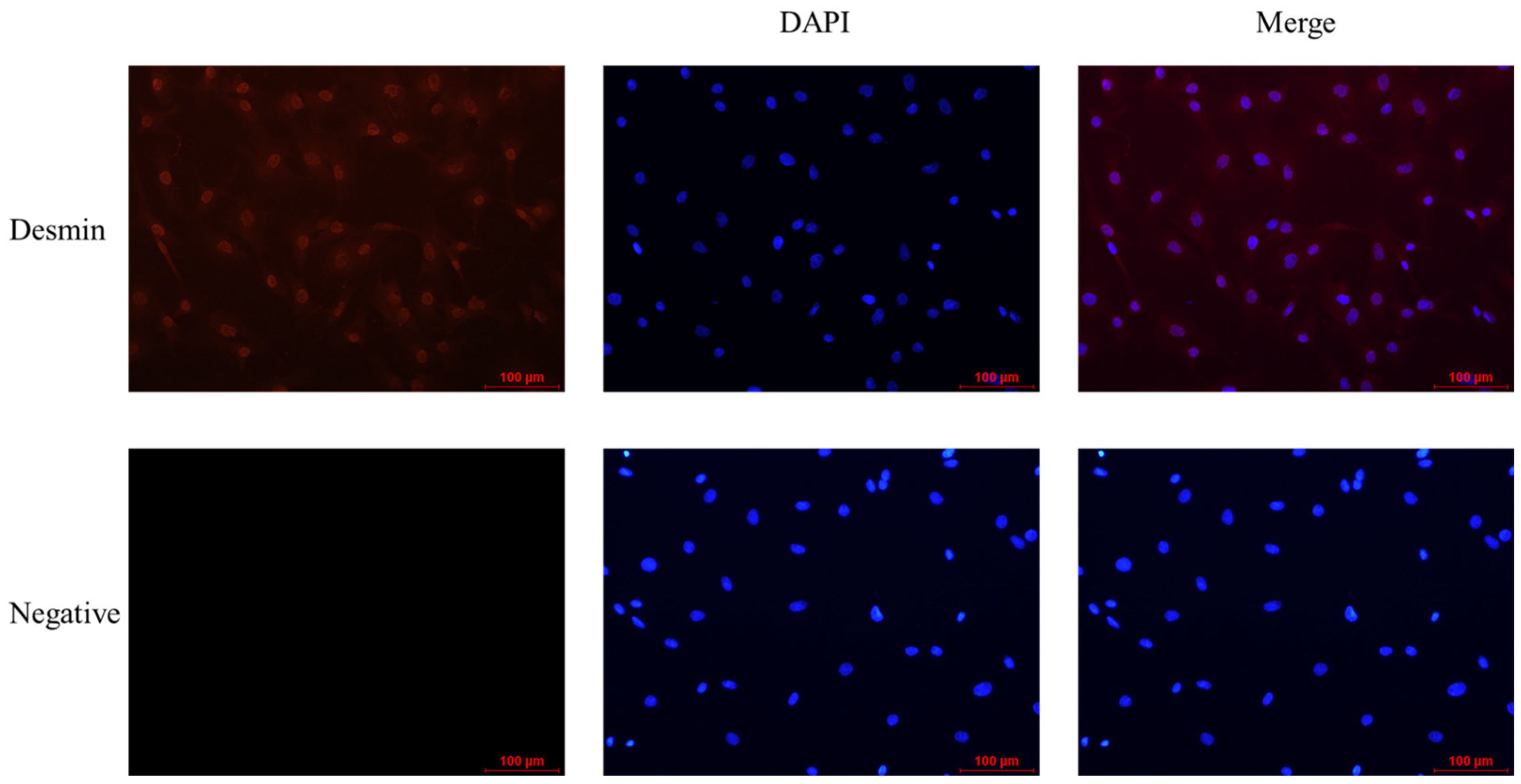
The purity of myoblasts was analyzed by indirect immunofluorescence, which showed that myoblasts bound to anti-Desmin fluoresced red, while unbound myoblasts did not. Myoblasts bound to anti-Desmin were observed under different fields of view, and by manual counting, the purity of myoblasts was calculated to be greater than 90% (Figure 2) and thus could be used for subsequent experiments.
3.3. Assay of shRNA Interference Efficiency
3.4. Effect of Silencing FOXO1 Gene Expression on Myoblast Proliferation
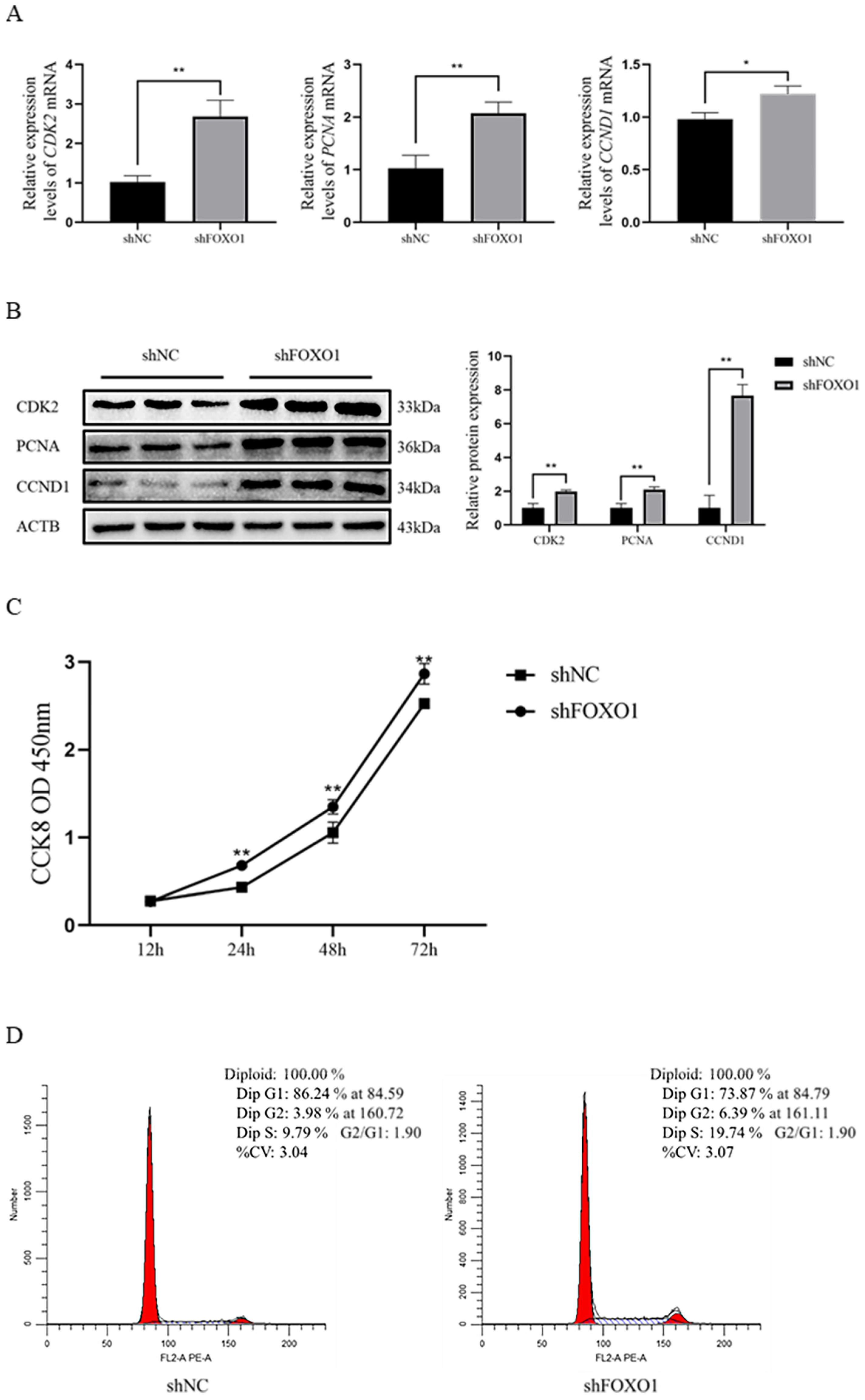
Results revealed that the expression of key genes (PCNA, CDK2, and CCND1) was significantly increased (p < 0.05) at both the mRNA and protein levels after silencing the FOXO1 gene (Figure 4A,B). The CCK-8 assay results showed that the shFOXO1 group significantly promoted the value-addition of myoblasts compared to the shNC group (Figure 4C). In addition, flow cytometry results indicated that the number of cells in the G1 phase was significantly reduced (p < 0.05), and the number of cells in the S phase was significantly increased (p < 0.05) in the shFOXO1 group compared to the shNC group (Figure 4D). Altogether, these results suggest that shFOXO1 promoted the proliferation of myoblasts.
3.5. Effect of Silencing FOXO1 Gene Expression on Myoblast Differentiation
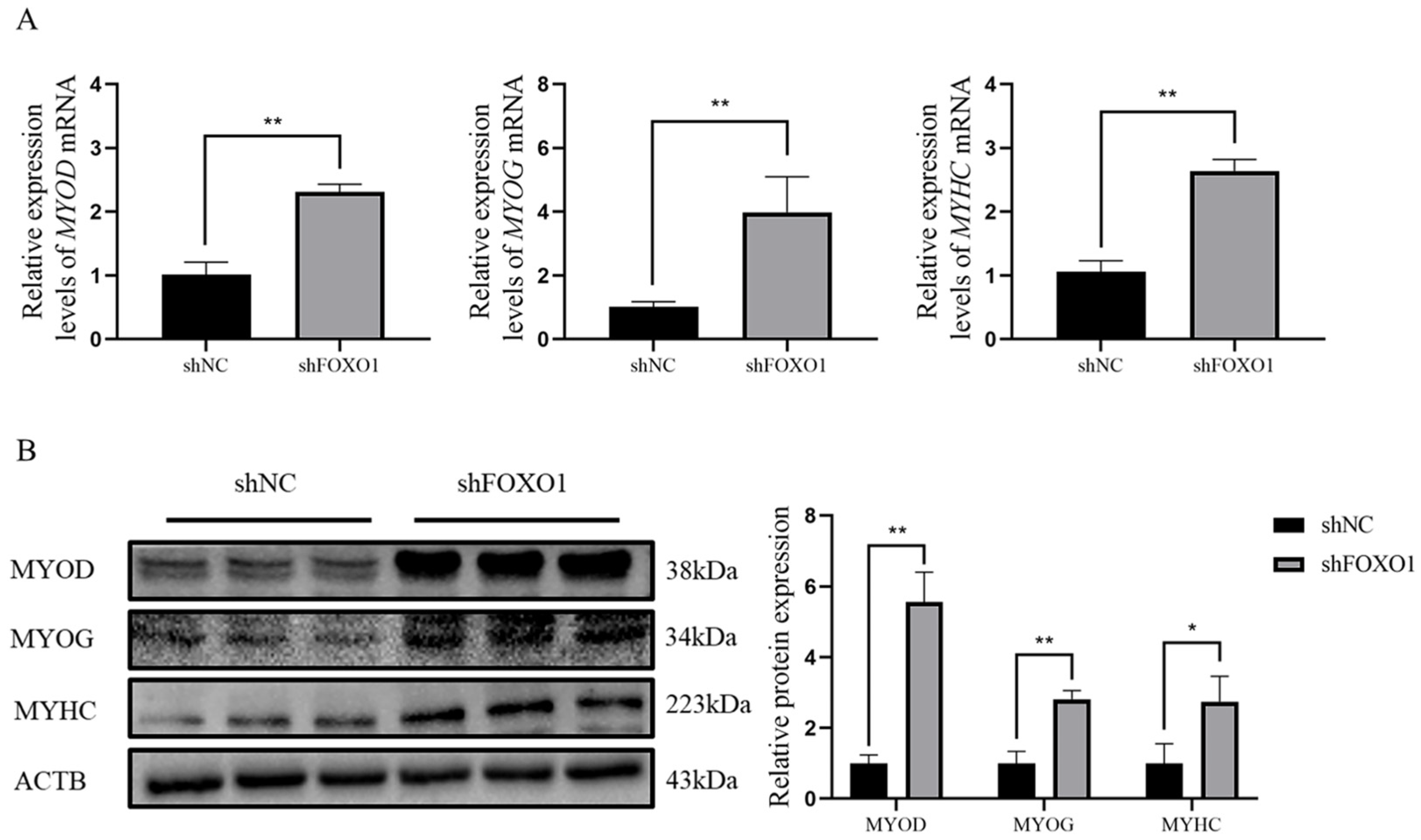
To further explore the effects of FOXO1 on bovine myoblast differentiation, qRT-PCR and Western blot analyses were applied to determine the relative expression of MYOD, MYOG, and MYHC at both mRNA and protein levels after silencing the FOXO1 gene. Results demonstrated that the expressions of MYOD, MYOG, and MYHC were significantly elevated at both levels (p < 0.05) after silencing the FOXO1 gene (Figure 5A,B), indicating that shFOXO1 promotes differentiation of myoblasts. Details of the original western blot figures are shown in Figure S2.
4. Discussion
Muscle development is an essential factor influencing animal growth and a crucial indicator of meat quality. Skeletal muscle development involves a process where myoblasts, differentiated from myosomes, progress toward a myogenic pathway through cell proliferation, terminal differentiation, and fusion into multinucleated muscle fibers [26]. Several studies have revealed that MYOD and MYOG, muscle-specific transcription factors, are key genes that regulate myoblasts’ fusion [27] and skeletal myogenesis [28,29]. It is worth noting that the skeletal muscle is a dynamic tissue with high contraction and good plasticity. Its composition of MYHC isoforms and metabolic activity determine the composition of different muscle fiber isoforms [16]. Muscle development is regulated by many genes whose expression is controlled at either the transcriptional or translational level, thereby affecting the mRNA and protein expression levels. The modification status of DNA methylation, one of the important epigenetic modifications, is dynamic during individual development and participates in the regulation of gene transcription, thereby affecting the expression of genes at the mRNA level [30]. In recent years, epigenetics has become a hotspot in livestock genetic breeding research. It is particularly important to explore the molecular genetic regulatory networks of genes involved in the regulation of muscle growth and development from an epigenetic perspective. In this study, BSP and qRT-PCR were applied to determine the effect of methylation of the FOXO1 promoter region on transcript levels. BSP results revealed differential methylation of the FOXO1 promoter region in muscle tissues from the CB and AB groups, whereas qRT-PCR results showed differences in the mRNA expression levels of FOXO1 at the two growth stages. These results suggest a tendency to significantly reduce gene expression at high DNA methylation levels, which lays the foundation for exploring the function of the differentially methylated region of the FOXO1 gene.
To further explore the potential function of the FOXO1 gene, we investigated the effect of FOXO1 on the proliferation and differentiation of bovine myoblasts through a gene interference strategy. A previous study reported that using cardiotoxin on transgenic mice overexpressing FOXO1 in the skeletal muscle caused muscle damage and that FOXO1 reduced the proliferative capacity of myoblasts and disrupted regeneration of the skeletal muscle [31]. In the present study, we found that interference with the FOXO1 gene caused a significant increase in the expression of the cell proliferation marker gene (PCNA), an essential gene for cell proliferation, and a significant increase in the expression abundance of the cell cycle-related proteins CDK2 and CCND1. In contrast, Qi et al. found no significant difference in the relative abundance of CCND1 and CCND2 gene transcription products after overexpressing the FOXO1 gene in sheep granulosa cells [11]. However, FOXO1 overexpression increased the relative abundance of P21 and P27 proteins, which are important cell cycle protein-dependent kinase inhibitors that can cause cell cycle arrest and proliferation through specific molecular mechanisms [32,33,34]. In addition, another study that explored curcumin-induced FOXO1 inhibition of lung cancer progression and metastasis found that activation of FOXO1 inhibited the spread of lung cancer cells by downregulating CCND1 gene expression [35]. These studies suggest that the FOXO1 gene regulates cellular proliferation but may affect cellular proliferation through different signaling pathways in different cell types. In this study, CCK-8 and flow cytometry assays were also applied to evaluate the effect of silencing the FOXO1 gene on myoblasts, with the obtained results validating that the FOXO1 gene negatively regulates the proliferation of bovine myoblasts.
Previous studies have reported that FOXO1 inhibits early differentiation of myoblasts and regulates the type of skeletal muscle fibers, with transgenic mice exhibiting significantly reduced skeletal muscle mass, impaired muscle function, and reduced abundance of slow fiber-related gene expression [36,37]. The FOXO1 protein mainly exerts its function in the nucleus. One study found that when myoblasts undergo multinuclear fusion to form myotubes, the process is associated with the phosphorylation state of FOXO1 protein, which is translocated to the cytoplasm and loses its regulatory role, suggesting that myoblasts require inhibition of FOXO1 protein activity during early differentiation [38]. Wu et al. reported that overexpression of the FOXO1 gene significantly inhibited the differentiation of C2C12 myoblasts [15]. Herein, we found that interference with the FOXO1 gene significantly increased the expression levels of the MYOD, MYOG, and MYHC genes and promoted the differentiation of myoblasts, which is consistent with Wu et al. Liu et al. found that skeletal muscle weight gain and MYOD expression abundance were significantly increased in mice after reducing FOXO1 expression in C2C12 myoblasts by RNA oligonucleotides [39].
However, one study reported that FOXO1 is involved in the late differentiation of myoblasts, and its transcript-level expression is gradually upregulated during the differentiation of bovine myoblasts, with a slight but not significant decrease in expression when myotubes are fully formed, suggesting a tendency for FOXO1 to induce differentiation of bovine skeletal muscle [17]. In addition, Bois et al. demonstrated that FOXO1 plays a role in cell cycle and apoptosis and controls the fusion rate of myotube formation during mouse myoblast differentiation [40]. This also suggests that FOXO1 may play a bidirectional regulatory role in the differentiation of myoblasts.
5. Conclusions
This study provides preliminary evidence that high methylation levels in the FOXO1 promoter region are associated with low mRNA expression. However, more data is needed to reveal this trend’s exact impact. Silencing FOXO1 gene expression promoted the expression of the proliferation key genes (CDK2, PCNA, and CCND1) and the differentiation key genes (MYOG, MYOD, and MYHC) at both mRNA and protein levels. Moreover, CCK-8 and flow cytometry assays further validated that silencing FOXO1 promotes the proliferation of myoblasts. In summary, this study has revealed that the level of methylation in the promoter region of FOXO1 leads to changes in its gene expression, which in turn affect the proliferation and differentiation of bovine myoblasts. Overall, our findings provide a theoretical basis for future studies on the regulation of epigenetics in the development of bovine myoblasts.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ani13020319/s1, Figure S1: Study on the DNA methylation level of the FOXO1 gene promoter. (A): The FOXO1 promoter region was analyzed by online software for the presence of two CpG islands at −366 bp and −1077 bp. (B): The average methylation level of each site in the longissimus dorsi CpG island. Rows indicate different age groups; columns indicate each CpG site with lower overall methylation levels. Figure S2: Original western blot figures.
Author Contributions
Conceptualization, P.S., Y.R. and H.X.; methodology, P.S., Y.R. and H.X.; validation, P.S., Y.R. and J.S.; formal analysis, P.S. and J.X.; investigation, P.S.; resources, P.S.; data curation, P.S.; writing—original draft preparation, P.S.; writing—review and editing, P.S., W.L. and H.X.; visualization, P.S.; supervision, W.L. and Y.R.; project administration, P.S., Y.R. and H.X.; funding acquisition, H.X. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China Project (31571279), the Ministry of Education Program to Promote Research Cooperation, and the High-Level Talent Training with U.S. University Region ([2015] 2062).
Institutional Review Board Statement
The use of experimental animals in this study was approved by the Laboratory Animal Ethics of Guizhou University (No. EAE-GZU-2021-E019, Guiyang, China; 1 November 2021).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Niu, Z.; Liu, H.; Zhou, M.; Wang, H.; Liu, Y.; Li, X.; Xiong, W.; Ma, J.; Li, X.; Li, G. Knockdown of c-Myc inhibits cell proliferation by negatively regulating the Cdk/Rb/E2F pathway in nasopharyngeal carcinoma cells. Acta Biochim. Biophys. Sin. 2015, 47, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, K.; Wang, Y.; Hua, H.; Qin, S.; Yang, A.; Shao, J. Ginsenoside Rg3 Combined with Oxaliplatin Inhibits the Proliferation and Promotes Apoptosis of Hepatocellular Carcinoma Cells via Downregulating PCNA and Cyclin D1. Biol. Pharm. Bull. 2019, 42, 900–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Hernández, J.M.; García-González, E.G.; Brun, C.E.; Rudnicki, M.A. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. Semin. Cell Dev. Biol. 2017, 72, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Michel, S. The FOXO’s Advantages of Being a Family: Considerations on Function and Evolution. Cells 2020, 9, 787. [Google Scholar]
- Xing, Y.-Q.; Li, A.; Yang, Y.; Li, X.-X.; Zhang, L.-N.; Guo, H.-C. The regulation of FOXO1 and its role in disease progression. Life Sci. 2018, 193, 124–131. [Google Scholar] [CrossRef]
- Hornsveld, M.; Dansen, T.B.; Derksen, P.W.; Burgering, B.M.T. Re-evaluating the role of FOXOs in cancer. Semin. Cancer Biol. 2018, 50, 90–100. [Google Scholar] [CrossRef]
- Furuyama, T.; Kitayama, K.; Shimoda, Y.; Ogawa, M.; Sone, K.; Yoshida-Araki, K.; Hisatsune, H.; Nishikawa, S.; Nakayama, K.; Nakayama, K.; et al. Abnormal angiogenesis in Foxo1 (Fkhr)-deficient mice. J. Biol. Chem. 2004, 279, 34741–34749. [Google Scholar] [CrossRef] [Green Version]
- Hosaka, T.; Biggs, W.H.; Tieu, D.; Boyer, A.D.; Varki, N.M.; Cavenee, W.K.; Arden, K.C. Disruption of forkhead transcription factor (FOXO) family members in mice reveals their functional diversification. Proc. Natl. Acad. Sci. USA. 2004, 101, 2975–2980. [Google Scholar] [CrossRef] [Green Version]
- Gross, D.N.; Wan, M.; Birnbaum, M.J. The role of FOXO in the regulation of metabolism. Curr. Diabetes Rep. 2009, 9, 208–214. [Google Scholar] [CrossRef]
- Chen, J.; Lu, Y.; Tian, M.; Huang, Q. Molecular mechanisms of FOXO1 in adipocyte differentiation. J. Mol. Endocrinol. 2019, 62, 239–253. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.; Guo, X.; Jia, K.; Jing, J.; Dang, W.; Li, Y.; Qin, X.; Li, P.; Ren, Y.; Liu, W.; et al. Effects of FOXO1 on the proliferation and cell cycle-, apoptosis- and steroidogenesis-related genes expression in sheep granulosa cells. Anim. Reprod. Sci. 2020, 221, 106604. [Google Scholar] [CrossRef]
- Daniele, L.B.; Katia, A.; Maria, R.C. FoxO1 at the nexus between fat catabolism and longevity pathways. Biochim. Biophys. Acta 2014, 1841, 1555–1560. [Google Scholar]
- Ana, O.; Alejo, E.; Elena, L.; Maribel, M.; Gonzalo, G.; Marta, C.; Francisca, M.; Joaquin, P.; Sonia, M.; Eduardo, R.; et al. Pten Positively Regulates Brown Adipose Function, Energy Expenditure, and Longevity. Cell Metab. 2012, 15, 382–394. [Google Scholar]
- Wu, Y.J.; Fang, Y.H.; Chi, H.C.; Chang, L.C.; Chung, S.Y.; Huang, W.C.; Wang, X.W.; Lee, K.W.; Chen, S.L. Insulin and LiCl synergistically rescue myogenic differentiation of FoxO1 over-expressed myoblasts. PLoS ONE 2014, 9, 88450. [Google Scholar] [CrossRef] [Green Version]
- Laura, I.; Fabio, C.; Maria, R.C. Adipose Tissue and FoxO1: Bridging Physiology and Mechanisms. Cells 2020, 9, 849. [Google Scholar]
- Xu, M.; Chen, X.; Chen, D.; Yu, B.; Huang, Z. FoxO1: A novel insight into its molecular mechanisms in the regulation of skeletal muscle differentiation and fiber type specification. Oncotarget 2017, 8, 10662–10674. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liu, K.; Huang, Y.; Lan, X.; Chen, H. Differential expression of FOXO1 during development and myoblast differentiation of Qinchuan cattle and its association analysis with growth traits. Sci. China Life Sci. 2018, 61, 826–835. [Google Scholar] [CrossRef]
- Sun, Y.; Xue, J.; Guo, W.; Li, M.; Huang, Y.; Lan, X.; Lei, C.; Zhang, C.; Chen, H. Haplotypes of bovine FoxO1 gene sequence variants and association with growth traits in Qinchuan cattle. J. Genet. 2013, 92, 8–14. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, Y. Reversing DNA methylation: Mechanisms, genomics, and biological functions. Cell 2014, 156, 45–68. [Google Scholar] [CrossRef] [Green Version]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Kazemier, H.G.; de Groote, M.L.; Ruiters, M.H.J.; Xu, G.; Rots, M.G. Induced DNA demethylation by targeting Ten-Eleven Translocation 2 to the human ICAM-1 promoter. Nucleic Acids Res. 2014, 42, 1563–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Cao, X.K.; Peng, S.J.; Wang, X.G.; Li, Z.; Elnour, I.E.; Huang, Y.Z.; Lan, X.Y.; Chen, H. Transcriptional regulation of the bovine FGFR1 gene facilitates myoblast proliferation under hypomethylation of the promoter. J. Cell. Physiol. 2020, 235, 8667–8678. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Su, X.; Tian, Y.; Song, G.; Zan, L.; Wang, H. Effect of Actin Alpha Cardiac Muscle 1 on the Proliferation and Differentiation of Bovine Myoblasts and Preadipocytes. Animals 2021, 11, 3468. [Google Scholar] [CrossRef] [PubMed]
- Rando, T.A.; Blau, H.M. Primary mouse myoblast purification, characterization, and transplantation for cell-mediated gene therapy. J. Cell Biol. 1994, 125, 1275–1287. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Takahashi, H.; Kitagawa, E.; Watanabe, H.; Sakurada, T.; Aso, H.; Yamaguchi, T. AMPK activation by AICAR inhibits myogenic differentiation and myostatin expression in cattle. Cell Tissue Res. 2012, 349, 615–623. [Google Scholar] [CrossRef]
- Margaret, B. Skeletal muscle formation in vertebrates. Curr. Opin. Genet. Dev. 2001, 11, 440–448. [Google Scholar]
- Zammit, P.S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin. Cell Dev. Biol. 2017, 72, 19–32. [Google Scholar] [CrossRef]
- Pipes, G.C.T.; Creemers, E.E.; Olson, E.N. The myocardin family of transcriptional coactivators: Versatile regulators of cell growth, migration, and myogenesis. Genes Dev. 2006, 20, 1545–1556. [Google Scholar] [CrossRef] [Green Version]
- Potthoff, M.J.; Olson, E.N. MEF2: A central regulator of diverse developmental programs. Development 2007, 134, 4131–4140. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Wang, L. Cell-Free DNA Methylation Profiling Analysis-Technologies and Bioinformatics. Cancers 2019, 11, 1741. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, A.; Hatazawa, Y.; Hirose, Y.; Ono, Y.; Kamei, Y. FOXO1 delays skeletal muscle regeneration and suppresses myoblast proliferation. Biosci. Biotechnol. Biochem. 2016, 80, 1531–1535. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.; Dutta, A. p21 in cancer: Intricate networks and multiple activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar] [CrossRef]
- Maryam, A.; Maryam, K.; Khalil, A.; Nazanin, B.; Sadra, S.T.; Mahmood, M.; Ansar, K. Multiple functions of p27 in cell cycle, apoptosis, epigenetic modification and transcriptional regulation for the control of cell growth: A double-edged sword protein. DNA Repair 2018, 69, 63–72. [Google Scholar]
- Nikolaos, Z.; Vasilios, P.; Dimitrios, D.; Konstantinos, S.; Christos, V.; Despina, N.P.; Konstantinos, K. A potential role of cyclin-dependent kinase inhibitor 1 (p21/WAF1) in the pathogenesis of endometriosis: Directions for future research. Med. Hypotheses 2019, 133, 109414. [Google Scholar]
- Li, Z.; Zhang, L.; Wang, H.; Ma, J.; Sun, J. Curcumin inhibits lung cancer progression and metastasis through induction of FOXO1. Tumour Biol. 2014, 35, 111–116. [Google Scholar] [CrossRef]
- Kamei, Y.; Miura, S.; Suzuki, M.; Kai, Y.; Mizukami, J.; Taniguchi, T.; Mochida, K.; Hata, T.; Matsuda, J.; Aburatani, H.; et al. Skeletal muscle FOXO1 (FKHR) transgenic mice have less skeletal muscle mass, down-regulated Type I (slow twitch/red muscle) fiber genes, and impaired glycemic control. J. Biol. Chem. 2004, 279, 41114–41123. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Chen, K.; Abdel, K.W.; Ward, J.L.; Yang, H.; Chabi, B.; Wrutniak-Cabello, C.; Tong, Q. Regulation of skeletal muscle oxidative capacity and muscle mass by SIRT3. PLoS ONE 2014, 9, 85636. [Google Scholar] [CrossRef]
- Hribal, M.L.; Nakae, J.; Kitamura, T.; Shutter, J.R.; Accili, D. Regulation of insulin-like growth factor-dependent myoblast differentiation by Foxo forkhead transcription factors. J. Cell Biol. 2003, 162, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.M.; Yang, Z.; Liu, C.W.; Wang, R.; Tien, P.; Dale, R.; Sun, L.Q. Effect of RNA oligonucleotide targeting Foxo-1 on muscle growth in normal and cancer cachexia mice. Cancer Gene Ther. 2007, 14, 945–952. [Google Scholar] [CrossRef]
- Bois, P.R.; Grosveld, G.C. FKHR (FOXO1a) is required for myotube fusion of primary mouse myoblasts. EMBO J. 2003, 22, 1147–1157. [Google Scholar] [CrossRef]
Figure 1.
Study on the DNA methylation level of the FOXO1 gene promoter. (A,B) CpG islands in the FOXO1 promoter region (CpG1: −653 to −1077; CpG2: −366 to −676). CpG sites and primer sequences on CpG islands (marked by underlining), where CpG1 and CpG2 contain 43 and 29 CpG sites, respectively. (C) Histogram of total methylation levels. Total methylation levels in CB and AB groups were significantly different (p < 0.01). (D) The mRNA expression levels of FOXO1 in muscle tissue at different developmental stages were significantly different (p < 0.01). Two asterisks (**) indicate significant differences (p < 0.01).
Figure 1.
Study on the DNA methylation level of the FOXO1 gene promoter. (A,B) CpG islands in the FOXO1 promoter region (CpG1: −653 to −1077; CpG2: −366 to −676). CpG sites and primer sequences on CpG islands (marked by underlining), where CpG1 and CpG2 contain 43 and 29 CpG sites, respectively. (C) Histogram of total methylation levels. Total methylation levels in CB and AB groups were significantly different (p < 0.01). (D) The mRNA expression levels of FOXO1 in muscle tissue at different developmental stages were significantly different (p < 0.01). Two asterisks (**) indicate significant differences (p < 0.01).

Figure 2.
The results of indirect immunofluorescence analysis of myoblasts’ purity were applied with anti-Desmin (red, 1:200, Bioss, Beijing, China), and the nuclei were counterstained with DAPI (blue) (scale bar, 100 μm).
Figure 2.
The results of indirect immunofluorescence analysis of myoblasts’ purity were applied with anti-Desmin (red, 1:200, Bioss, Beijing, China), and the nuclei were counterstained with DAPI (blue) (scale bar, 100 μm).

Figure 3.
shRNA interference efficiency assay. (A,B) The qRT-PCR and Western blot assay for interference efficiency of FOXO1 shRNA. Two asterisks (**) indicate significant differences (p < 0.01).
Figure 3.
shRNA interference efficiency assay. (A,B) The qRT-PCR and Western blot assay for interference efficiency of FOXO1 shRNA. Two asterisks (**) indicate significant differences (p < 0.01).

Figure 4.
The effect of shFOXO1 on the proliferation of myoblasts. (A) qRT-PCR to determine the expression of CDK2, PCNA, and CCND1 mRNA in the shFOXO1 and shNC groups. (B) Western blot to detect the expression of CDK2, PCNA, and CCND1 protein in the shFOXO1 and shNC groups (greyscale analysis). (C) CCK-8 assay was used to detect the proliferation of transfected cells. (D) Flow cytometry was used to determine the DNA content of myoblasts 48 h after transfection. An asterisk (*) indicates significant differences (p < 0.05), and two asterisks (**) indicate significant differences (p < 0.01).
Figure 4.
The effect of shFOXO1 on the proliferation of myoblasts. (A) qRT-PCR to determine the expression of CDK2, PCNA, and CCND1 mRNA in the shFOXO1 and shNC groups. (B) Western blot to detect the expression of CDK2, PCNA, and CCND1 protein in the shFOXO1 and shNC groups (greyscale analysis). (C) CCK-8 assay was used to detect the proliferation of transfected cells. (D) Flow cytometry was used to determine the DNA content of myoblasts 48 h after transfection. An asterisk (*) indicates significant differences (p < 0.05), and two asterisks (**) indicate significant differences (p < 0.01).

Figure 5.
Effect of shFOXO1 on myoblast differentiation. (A) qRT-PCR to determine the expression of MYOG, MYOD, and MYHC mRNA in shFOXO1 and shNC groups. (B) Western blot to detect the expression of MYOG, MYOD, and MYHC proteins in shFOXO1 and shNC groups (greyscale analysis). An asterisk (*) indicates significant differences (p < 0.05), and two asterisks (**) indicate significant differences (p < 0.01).
Figure 5.
Effect of shFOXO1 on myoblast differentiation. (A) qRT-PCR to determine the expression of MYOG, MYOD, and MYHC mRNA in shFOXO1 and shNC groups. (B) Western blot to detect the expression of MYOG, MYOD, and MYHC proteins in shFOXO1 and shNC groups (greyscale analysis). An asterisk (*) indicates significant differences (p < 0.05), and two asterisks (**) indicate significant differences (p < 0.01).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used in this study.
| Primer | Primer Sequence (5′-3′) | Accession Numbers | Amplified DNA Fragment (bp) |
|---|---|---|---|
| Primers used in real-time PCR | |||
| FOXO1 | F: GCAGATTTACGAGTGGATGGTC R: GCAGGGACAGATTATGACGAA | XM_025000053.1 | 107 |
| PCNA | F: ACATCAGCTCAAGTGGCGTGAAC R: GCAGCGGTAAGTGTCGAAGCC | NM_001034494.1 | 101 |
| CCND1 | F: CTGGTCCTGGTGAACAAACT R: ACAGAGGGCAACGAAGGT | NM_001046273.2 | 144 |
| CDK2 | F: CAAGTTGACGGGAGAAGTGGT R: CTTTATGAGCGGAAGAGGAAT | NM_001014934.1 | 247 |
| MYOD | F: GGCCGCTGTTTACTGTGGG R: CAGCCGCTGGTTTGGGTT | NM_001040478.2 | 162 |
| MYOG | F: TGGGCGTGTAAGGTGTGTAA R: TGCAGGCGCTCTATGTACTG | NM_001111325.1 | 197 |
| MYHC | F: GCCCACTTCTCCCTCATTCACT R: ACCCTTCTTCTTGCCACCTTTC | NM_174117.1 | 201 |
| β-actin | F: ATGATATTGCTGCGCTCGTGG R: TACGAGTCCTTCTGGCCCAT | NM_173979.3 | 151 |
| Primers used in methylation analysis | |||
| MF1-1 | F: GATTGATTTAGTGGATAGTTTG R: TTTTTCCTTTCCCTACAAT | Gene ID:506618 | 424 |
| MF1-2 | F: ATATATTGTAGGGAAAGGAAAA R: ACACTTTATTTACTACTAAAAAACC | 310 | |
Note: F stands for upstream primer, and R stands for downstream primer.
Table 2.
The specific shRNA sequences.
| shRNA Name | Sequences |
|---|---|
| shRNA1 | 5′-3′ CAGTCTGTCCGAGATCAGTAA |
| shRNA2 | 5′-3′ AGCGGGCTGGAAGAATTCAAT |
| shRNA3 | 5′-3′ CAGGACAACAAATCGAGTTAT |
| shRNA4 | 5′-3′ CTGTGACATGGAGTCCATCAT |
| shNC | 5′-3′ GTTCTCCGAACGTGTCACGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shi, P.; Ruan, Y.; Liu, W.; Sun, J.; Xu, J.; Xu, H. Analysis of Promoter Methylation of the Bovine FOXO1 Gene and Its Effect on Proliferation and Differentiation of Myoblasts. Animals 2023, 13, 319. https://doi.org/10.3390/ani13020319
AMA Style
Shi P, Ruan Y, Liu W, Sun J, Xu J, Xu H. Analysis of Promoter Methylation of the Bovine FOXO1 Gene and Its Effect on Proliferation and Differentiation of Myoblasts. Animals. 2023; 13(2):319. https://doi.org/10.3390/ani13020319
Chicago/Turabian StyleShi, Pengfei, Yong Ruan, Wenjiao Liu, Jinkui Sun, Jiali Xu, and Houqiang Xu. 2023. "Analysis of Promoter Methylation of the Bovine FOXO1 Gene and Its Effect on Proliferation and Differentiation of Myoblasts" Animals 13, no. 2: 319. https://doi.org/10.3390/ani13020319
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.