
Morphostructural Differences between the Historical Genetic Lines of the Spanish Merino Sheep

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Measurements
2.2. Statistical Analysis
3. Results
3.1. Head
3.2. Trunk and Extremities
3.3. Zoometric Indexes
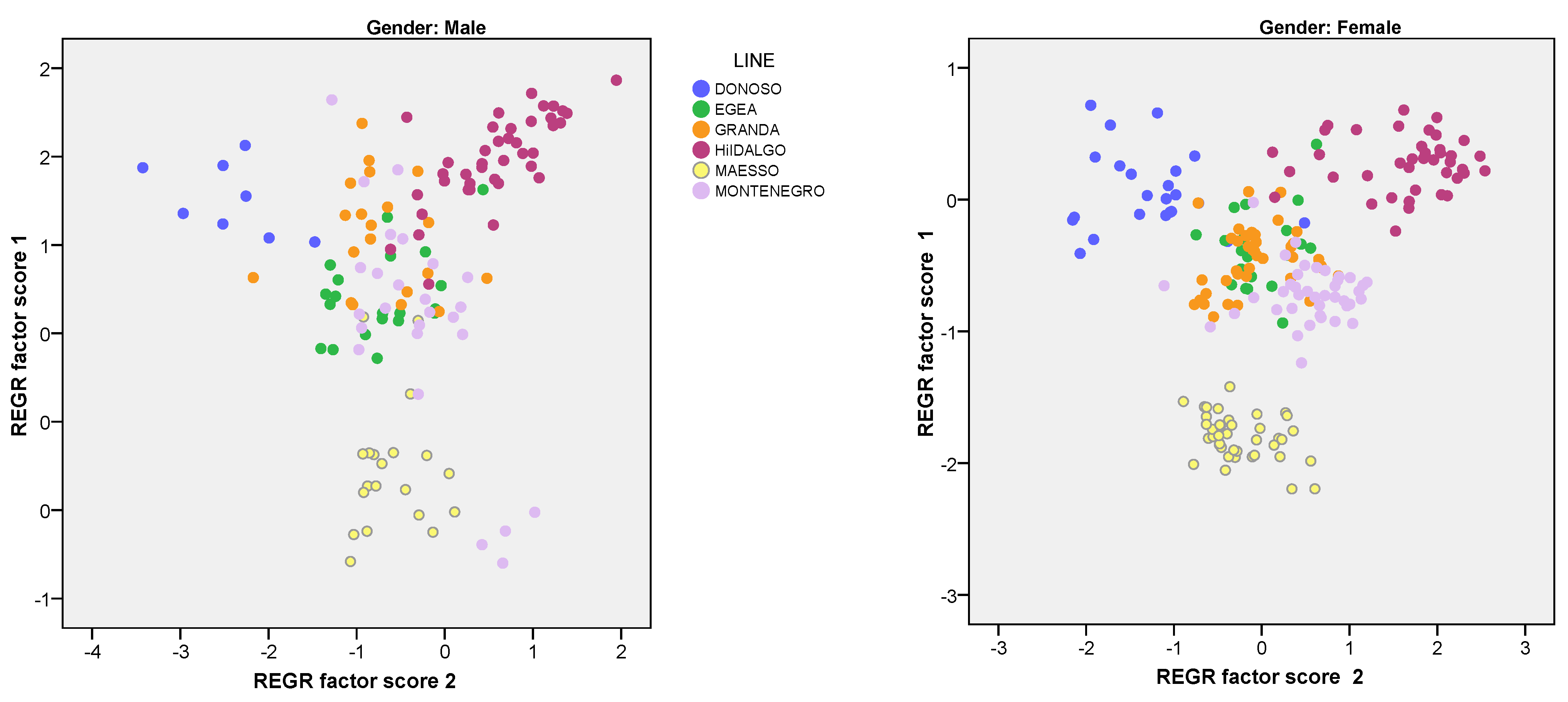
3.4. Principal Component Analysis (ACP)
3.5. Discriminant Analysis
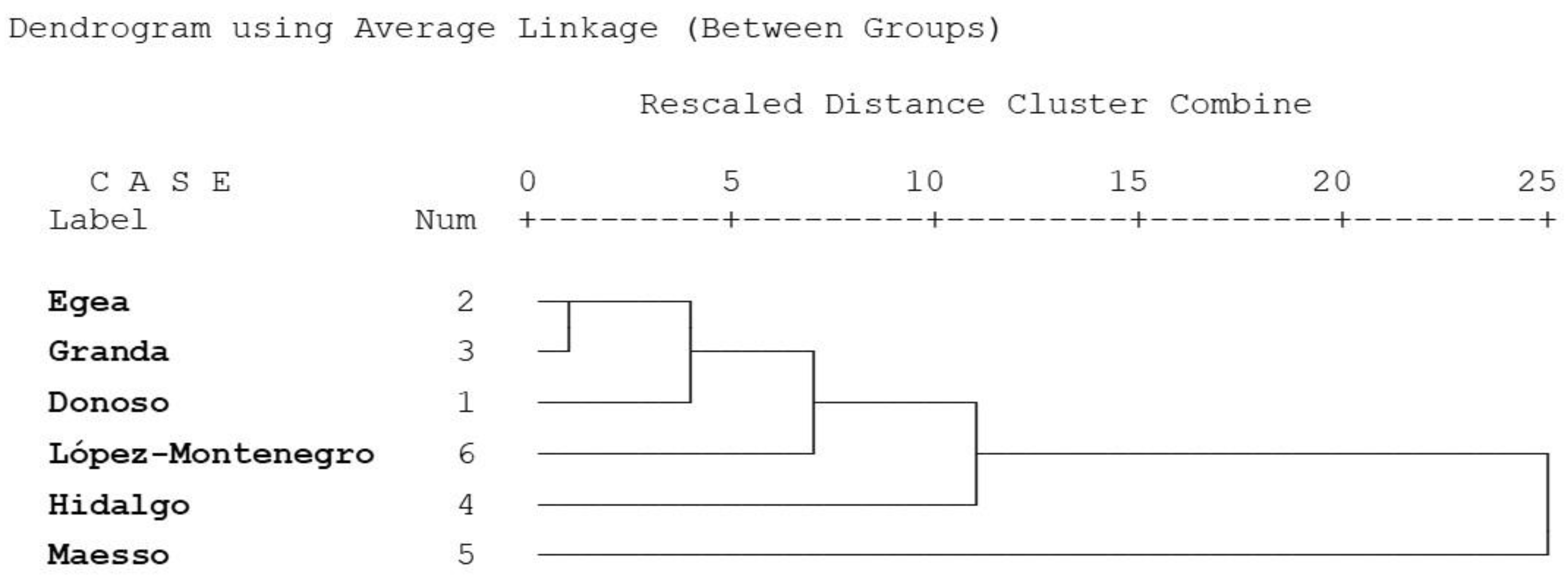
3.6. Cluster Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kijas, J.W.; Lenstra, J.A.; Hayes, B.; Boitard, S.; Porto Neto, L.R.; San Cristobal, M.; Servin, B.; McCulloch, R.; Whan, V.; Gietzen, K.; et al. Genome-Wide analysis of the world’s sheep breeds reveals high levels of historic mixture and strong recent selection. PLoS Biol. 2012, 10, e1001258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, E. Raza Merina y sus Cruces en la Producción de Carne; Pesca y Alimentación; Ministerio de Agricultura: Madrid, Spain, 1994. [Google Scholar]
- Sánchez-Belda, A.S.-T.M. Razas Ovinas Españolas; Ministerio de Agricultura: Madrid, Spain, 1986; p. 887. [Google Scholar]
- Ciani, E.; The International Sheep Genomics Consortium; Lasagna, E.; D’Andrea, M.; Alloggio, I.; Marroni, F.; Ceccobelli, S.; Bermejo, J.V.D.; Sarti, F.M.; Kijas, J.; et al. Merino and Merino-derived sheep breeds: A genome-wide intercontinental study. Genet. Sel. Evol. 2015, 47, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarti, D. Le Razze derivate Merinos ad attitudine carne nel centro sud europeo. L’allevatore Di Ovini E Caprini 1996, 7, 1–2. [Google Scholar]
- Ciani, E.; Crepaldi, P.; Nicoloso, L.; Lasagna, E.; Sarti, F.M.; Moioli, B.; Napolitano, F.; Carta, A.; Usai, G.; D’Andrea, M.; et al. Genome-wide analysis of Italian sheep diversity reveals a strong geographic pattern and cryptic relationships between breeds. Anim. Genet. 2013, 45, 256–266. [Google Scholar] [CrossRef]
- Peña Blanco, F.; Alcalde Aldea, M.J. Las razas ovinas Integradas en Andalucía: Merino Precoz Francés, Ille de France, Fleischscaff, Landschaff y Lacaune; Las razas ganaderas de Andalucía; Patrimonio ganadero Patrimonio ganadero andaluz; Junta de Andalucía: El Torno: El Torno, Spain, 2007; Volume II. [Google Scholar]
- Azor, O.C.I.; Valera, M.; Arranz, J.J.; Medina, C.; Gutiérrez, J.P.; Goyache, F.; Muñoz, A.; Medina, C. Análisis preliminar de la estructura genética del Merino: Situación de las estirpes tradicionales mediante análisis genealógico y molecular. In Proceedings of the En XIV Reunión Nacional de Mejora Genética Animal: Asociación Interprofesional para el Desarrollo Agrario (AIDA); 2008; Volume 104, pp. 295–302. [Google Scholar]
- Garzón, R.; Luque, J.; Llanes, D.; Povedano, C.; Rodero, A.; Rodero, J.; Zarazaga, I. Fundamentos Históricos y Genéticos del Merino Español. Publicaciones del Monte de Piedad y Caja de Ahorros de Córdoba: Cordoba, Spain, 1977; p. 145. [Google Scholar]
- Laguna, E. Historia del Merino; Ministerio de Agricultura: Madrid, Spain, 1986. [Google Scholar]
- Esteban, M. La raza Merina y sus Cruces en la Producción de Carne; MAPA, Ministerio de Agricultura Pesca y Alimentación del Gobierno de España: Madrid, Spain, 1994; p. 175. [Google Scholar]
- Granero, A.; Anaya, G.; Demyda-Peyrás, S.; Alcalde, M.J.; Arrebola, F.; Molina, A. Genomic Population Structure of the Main Historical Genetic Lines of Spanish Merino Sheep. Animals 2022, 12, 1327. [Google Scholar] [CrossRef]
- Zaitoun, I.S.; Tabbaa, M.; Bdour, S. Differentiation of native goat breeds of Jordan on the basis of morphostructural characteristics. Small Rumin. Res. 2005, 56, 173–182. [Google Scholar] [CrossRef]
- Mujica, L.; Hernández, C.A.; Vargas-Bayona, J.E.; Leaño Á, J.; Quiceno, V.A.; Novoa, C.S.; Bedoya, J.T. Diversidad genética de la cabra santandereana (Capra hircus) mediante microsatélites: Modelo aplicable para la conservación y aprovechamien to genético de las cabras nativas mexicanas. El Sudcaliforniano 2014, 7, 147. [Google Scholar]
- Andersson, L.; Georges, M. Domestic-Animal Genomics: Deciphering the Genetics of Complex Traits. Nat. Rev. Genet. 2004, 5, 202–212. [Google Scholar] [CrossRef]
- Herrera García, M.R.S.E. El concepto de raza. Un enfoque epistemológico. Arch. De Zootec. 2000, 49, 12. [Google Scholar]
- Herrera, M.; Luque, M. Morfoestructura y sistemas para el futuro en la valoración morfológica. In Valoración Morfológica de los Animales Domésticos; Ministerio de Medio Ambiente y Medio Rural y Marino: Madrid, Spain, 2009. [Google Scholar]
- Parés i Casanova, P.M. Zoometría. In Valoración Morfológica de los animales domésticos; Sañudo, A.C., Ed.; Ministerio de Medio Ambiente y Medio Rural y Marino: Madrid, Spain, 2009. [Google Scholar]
- Aparicio, G. Exterior de los Grandes Animales Domésticos; Imprenta Moderna: Córdoba, Spain, 1960; pp. 5–27. [Google Scholar]
- Sañudo, C. Valoración Morfológica de los Animales Domésticos; Ministerio de Medio Ambiente y Medio Rural y Marino; Ministerio de Medio Ambiente y Medio Rural y Marino: Madrid, Spain, 2009. [Google Scholar]
- Granero, A.M.A.; Anaya, G.; Ziadib, C.; Alcalde, M.J. Morphometric differences based on quantitative traits between different genetic lines in the Merino Español sheep breed. ASPA 24th Congress Book of Abstract. Ital. J. Anim. Sci. 2021, 20 (Suppl. 1), 115. [Google Scholar]
- ARCA. Sistema Nacional de Información de Razas; P.y.A. Ministerio de Agricultura: Madrid, Spain, 2022. [Google Scholar]
- Díaz Montilla, R. Estudio Biométrico Estación Pecuaria Regional de Badajoz; Ganado Lanar: Badajoz, Spain, 1945; Volume 14. [Google Scholar]
- Esteban, C. Spanish Sheep Breeds; Ministry of Agriculture, Fisheries and Food: Madrid, Spain, 2003; p. 287. [Google Scholar]
- Fontenay, L.D. Le Betail En Ecosse. Race Bovine, Pratiques d’Elevage Et d’Engraissement Des Fermiers Anglais; Librairie D’Agriculture et D´Horticulture: Paris, France, 1862. [Google Scholar]
- Perucho, L.; Lauvie, A.; Nozières-Petit, M.O.; Moulin, C.H. Adaptation of local breeds is not only a biological question: Illustration with four French Mediterranean sheep breeds. Addressing Chall. Agro-Pastor. Farming Syst. 2022, 129, 25. [Google Scholar]
- Hagen, J.B. Bergmann’s Rule, Adaptation, and Thermoregulation in Arctic Animals: Conflicting Perspectives from Physiology, Evolutionary Biology, and Physical Anthropology After World War II. J. Hist. Biol. 2017, 50, 235–265. [Google Scholar] [CrossRef] [PubMed]
- Joy, A.; Dunshea, F.R.; Leury, B.J.; Clarke, I.J.; DiGiacomo, K.; Chauhan, S.S. Resilience of small ruminants to climate change and increased environmental temperature: A review. Animals 2020, 10, 867. [Google Scholar] [CrossRef]
- Rodríguez Pascual, M. The Transhumance. Culture, Canyons and Travel; Edilesa: Leon, Spain, 2001; p. 50. [Google Scholar]
- Valera, M.; Arrebola, F.; Juárez, M.; Molina, A. Genetic improvement of wool production in Spanish Merino sheep: Genetic parameters and simulation of selection strategies. Anim. Prod. Sci. 2009, 49, 43–47. [Google Scholar] [CrossRef]
- Avellanet Torres, R. Conservación de Recursos Genéticos Ovinos en la raza Xisqueta: Caracterización Estructural, Racial y Gestión de la Diversidad en Programas “In Situ”; Universitat Autònoma de Barcelona: Bellaterra, Spain, 2007. [Google Scholar]
- Latorre, E.; Uribe, H.; Martínez, M.E.; Calderón, C.; De la Barra, R. Morphology differentiation and structural functionality of ewes due to incomplete crossbreeding. Int. J. Morphol. 2011, 29, 954–959. [Google Scholar] [CrossRef]
- Khargharia, G.; Kadirvel, G.; Kumar, S.; Doley, S.; Bharti, P.K.; Das, M. Principal component analysis of morphological traits of Assam Hill goat in Eastern Himalayan India. JAPS J. Anim. Plant Sci. 2015, 25, 1251–1258. [Google Scholar]
- Mavule, B.; Muchenje, V.; Bezuidenhout, C.C.; Kunene, N.W. Morphological structure of Zulu sheep based on principal component analysis of body measurements. Small Rumin. Res. 2013, 111, 23–30. [Google Scholar] [CrossRef]
- Marković, B.; Dovč, P.; Marković, M.; Radonjić, D.; Adakalić, M.; Simčič, M. Differentiation of some Pramenka sheep breeds based on morphometric characteristics. Arch. Anim. Breed. 2019, 62, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Yunusa, A.; Salako, A.; Oladejo, O. Morphometric characterization of Nigerian indigenous sheep using multifactorial discriminant analysis. Int. J. Biodivers. Conserv. 2013, 5, 661–665. [Google Scholar]
- Salako, A. Application of morphological indices in the assessment of type and function in sheep. Int. J. Morphol. 2006, 24, 13–18. [Google Scholar] [CrossRef]
- Yadav, D.K.; Jain, A.; Kulkarni, V.S.; Govindaiah, M.G.; Aswathnarayan, T.; Sadana, D.K. Classification of four ovine breeds of southern peninsular zone of India: Morphometric study using classical discriminant function analysis. SpringerPlus 2013, 2, 29. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Head length (HL): | From the occipital protuberance or nape to the midpoint of the upper jaw. |
| Head width (HW): | Maximum distance between the two orbits. |
| Wool extension on forehead “bow” (WF): | Extension of wool covering the area from the occipital protuberance to the midpoint of an imaginary line passing below the eyes (frontonasal suture). |
| Wool extension on cheeks (WCh): | Extension of wool covering the cheeks, which goes from the lower point of insertion of the ear to the end where the wool ends. |
| Ear length (EL): | Rectilinear distance between the base of insertion of the ear and its free end. |
| Shoulder width (SW): | Maximum distance between the highest points of the scapulo-humeral joints. |
| Neck length (NL): | Distance between the lower point of insertion of the ear and the upper end of the point of shoulder. |
| Lumbar back line length (LBL): | Distance between the most declining point of the withers and the external iliac tuberosity (tip of hip). |
| Rump width (RW): | Maximum distance between the two external iliac tuberosities or tips of the hip. |
| Rump length (RL): | Distance between the external iliac tuberosity (tip of hip) and the ischial tuberosity (tip of the buttock). |
| Buttock width (BuW): | Maximum distance between the animal’s thighs. |
| Buttock length (BL): | Distance between the ischial tuberosity (tip of the buttock) and the upper end of the leg. |
| Withers height (WH): | Distance from the ground to the highest point of the withers (inter-scapular region). |
| Bicostal diameter (BD): | Maximum width of the thorax in a vertical plane that passes behind the elbow (5th rib). |
| Height to substernal foramen (HSF): | Distance from the ground to the substernal foramen. |
| Dorso-sternal diameter (DSD): | Distance between the most declining point of the withers and the sternal region behind the elbow. |
| Longitudinal diameter (LD): | Distance between the point of shoulder and the tip of the ischium. |
| Rump height (RH): | Distance from the ground to the highest point of the internal tuberosities of the ileum (apex of the first process of the sacrum). |
| Thoracic perimeter (ThP): | Beginning at the most declining point of the withers, passing through the right side, sternum (immediately behind the elbow), left side, and returning to the withers. |
| Anterior and Posterior cannon bone perimeter (ACP and PCP): | Minimum perimeter of these cannon bones. |
| Genetic lines | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Donoso | Egea | Granda | Hidalgo | Maesso | López- Montenegro | Line | Gender | Line × Gender | |
| HL | 27.15 ± 0.35 c | 26.28 ± 0.31 b | 26.01 ± 0.24 b | 28.16 ± 0.18 d | 24.48 ± 0.32 a | 25.86 ± 0.27 b | <0.001 | <0.001 | <0.001 |
| HW | 11.66 ± 0.16 b | 12.05 ± 0.10 d | 11.94 ± 0.11 cd | 12.29 ± 0.08 e | 11.22 ± 0.11 a | 11.80 ± 0.10 bc | <0.001 | <0.001 | <0.001 |
| WF | 15.52 ± 0.39 b | 15.43 ± 0.19 b | 15.52 ± 0.21 b | 16.18 ± 0.22 c | 14.05 ± 0.20 a | 15.30 ± 0.22 b | <0.001 | <0.001 | 0.004 |
| WCh | 8.40 ± 0.40 ab | 9.68 ± 0.19 d | 10.90 ± 0.18 e | 8.76 ± 0.26 bc | 7.69 ± 0.15 a | 9.39 ± 0.21 cd | <0.001 | <0.001 | <0.001 |
| EL | 8.65 ± 0.15 a | 10.52 ± 0.12 c | 10.83 ± 0.09 c | 13.21 ± 0.07 d | 9.62 ± 0.08 b | 10.86 ± 0.06 c | <0.001 | >0.050 | 0.020 |
| SW | 21.97 ± 0.33 d | 21.49 ± 0.34 d | 20.62 ± 0.31 c | 22.68 ± 0.29 e | 17.18 ± 0.29 a | 19.58 ± 0.31 b | <0.001 | <0.001 | 0.044 |
| NL | 28.90 ± 0.08 b | 30.00 ± 0.16 c | 31.06 ± 0.25 d | 34.64 ± 0.19 e | 26.99 ± 0.24 a | 30.61 ± 0.17 d | <0.001 | <0.001 | <0.001 |
| WH | 72.10 ± 0.62 c | 70.90 ± 0.65 c | 69.51 ± 0.53 b | 74.78 ± 0.38 d | 61.43 ± 0.64 a | 70.82 ± 0.59 bc | <0.001 | <0.001 | <0.001 |
| BD | 32.50 ± 0.35 b | 32.48 ± 0.30 b | 32.18 ± 0.36 b | 35.06 ± 0.27 c | 29.15 ± 0.40 a | 31.84 ± 0.34 b | <0.001 | <0.001 | <0.001 |
| HSF | 41.34 ± 0.47 b | 40.73 ± 0.38 b | 40.36 ± 0.26 b | 43.30 ± 0.21 c | 34.36 ± 0.32 a | 41.24 ± 0.26 b | <0.001 | <0.001 | 0.008 |
| DSD | 31.58 ± 0.33 d | 31.30 ± 0.25 cd | 30.63 ± 0.32 bc | 32.99 ± 0.21 e | 28.36 ± 0.36 a | 30.10 ± 0.30 b | <0.001 | <0.001 | <0.001 |
| LD | 71.71 ± 0.76 c | 71.94 ± 0.82 c | 72.37 ± 0.72 c | 78.92 ± 0.64 d | 62.98 ± 0.55 a | 69.81 ± 0.68 b | <0.001 | <0.001 | >0.050 |
| LBL | 30.52 ± 0.20 b | 30.87 ± 0.37 b | 30.20 ± 0.27 b | 34.73 ± 0.71 c | 26.63 ± 0.33 a | 30.23 ± 0.21 b | <0.001 | <0.001 | >0.050 |
| RH | 71.85 ± 0.65 bc | 72.64 ± 0.63 c | 71.96 ± 0.60 bc | 76.32 ± 0.34 d | 62.38 ± 0.59 a | 70.89 ± 0.52 b | <0.001 | <0.001 | 0.001 |
| RW | 19.98 ±0.17 e | 18.77 ± 0.15 d | 18.26 ± 0.16 c | 19.56 ± 0.08 e | 16.22 ± 0.16 a | 17.69 ± 0.16 b | <0.001 | <0.001 | <0.001 |
| RL | 24.13 ± 0.31 d | 24.09 ± 0.26 d | 23.42 ± 0.29 c | 25.51 ± 0.20 e | 20.47 ± 0.24 a | 22.80 ± 0.24 b | <0.001 | <0.001 | 0.007 |
| BuW | 28.81 ± 0.29 e | 24.02 ± 0.29 c | 24.04 ± 0.26 c | 25.78 ± 0.18 d | 21.06 ± 0.22 a | 22.55 ± 0.28 b | <0.001 | <0.001 | >0.050 |
| BL | 18.47 ± 0.23 d | 17.56 ± 0.22 c | 17.45 ± 0.15 c | 19.15 ± 0.18 e | 13.73 ± 0.14 a | 16.70 ± 0.24 b | <0.001 | <0.001 | 0.002 |
| ThP | 99.42 ± 0.80 f | 91.76 ± 0.62 d | 89.42 ± 0.90 c | 95.31 ± 0.46 e | 79.07 ± 0.79 a | 85.56 ± 0.80 b | <0.001 | <0.001 | <0.001 |
| ACP | 9.55 ± 0.13 d | 9.09 ± 0.11 c | 9.12 ± 0.09 c | 9.41 ± 0.07 d | 7.93 ± 0.08 a | 8.70 ± 0.09 b | <0.001 | <0.001 | >0.050 |
| PCP | 11.06 ± 0.15 e | 10.16 ± 0.11 c | 10.17 ± 0.08 c | 10.50 ± 0.08 d | 8.78 ± 0.10 a | 9.80 ± 0.10 b | <0.001 | <0.001 | >0.050 |
| BW | 61.98 ± 2.23 cd | 64.21 ± 1.51 d | 59.43 ± 1.77 c | 73.35 ± 1.23 e | 42.29 ± 1.18 a | 52.74 ± 1.58 b | <0.001 | <0.001 | 0.001 |
| Genetic Lines | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Donoso | Egea | Granda | Hidalgo | Maesso | López- Montenegro | Line | Gender | Line × Gender | |
| BI | 72.20 ± 0.77 a | 78.41 ± 0.75 b | 81.02 ± 0.47 cd | 82.77 ± 0.45 d | 79.78 ± 0.44 bc | 81.67 ± 0.49 cd | <0.001 | <0.001 | <0.001 |
| CoI | 85.54 ± 2.51 c | 90.29 ± 1.59 d | 84.81 ± 1.95 c | 97.73 ± 1.26 e | 68.23 ± 1.23 a | 73.83 ± 1.67 b | <0.001 | <0.001 | <0.001 |
| PeI | 83.06 ± 0.86 c | 78.06 ± 0.51 ab | 78.24 ± 0.49 ab | 76.95 ± 0.56 a | 79.41 ± 0.49 b | 77.77 ± 0.59 ab | <0.001 | <0.001 | <0.001 |
| ThI | 102.90 ± 0.18 a | 103.78 ± 0.50 ab | 105.07 ± 0.38 bc | 106.22 ± 0.37 c | 102.75 ± 0.37 a | 105.76 ± 0.34 c | <0.001 | <0.001 | >0.050 |
| CeI | 42.98 ± 0.30 a | 45.94 ± 0.25 b | 45.93 ± 0.16 b | 43.65 ± 0.15 a | 45.95 ± 0.24 b | 45.69 ± 0.20 b | <0.001 | <0.001 | <0.001 |
| PI | 100.71 ± 0.86 de | 98.69 ± 0.50 cd | 96.28 ± 0.62 ab | 94.99 ± 0.47 a | 97.53 ± 0.50 bc | 101.62 ± 0.56 e | <0.001 | 0.005 | <0.001 |
| LCI | 15.79 ± 0.38 c | 14.32 ± 0.20 b | 15.91 ± 0.36 c | 13.01 ± 0.14 a | 19.29 ± 0.36 e | 17.15 ± 0.36 d | <0.001 | <0.001 | <0.001 |
| Components | |||
|---|---|---|---|
| Traits and Indexes | Factor 1 | Factor 2 | Communality |
| HL | 0.930 | −0.089 | 0.873 |
| HW | 0.855 | −0.115 | 0.745 |
| WF | 0.820 | −0.239 | 0.730 |
| WCh | 0.359 | −0.377 | 0.272 |
| EL | 0.485 | 0.788 | 0.856 |
| SW | 0.942 | −0.142 | 0.908 |
| NL | 0.810 | 0.476 | 0.883 |
| WH | 0.936 | 0.093 | 0.885 |
| BD | 0.953 | −0.011 | 0.909 |
| HSF | 0.804 | 0.263 | 0.716 |
| DSD | 0.934 | −0.055 | 0.875 |
| LD | 0.930 | 0.142 | 0.885 |
| LBL | 0.720 | 0.304 | 0.611 |
| RH | 0.942 | 0.114 | 0.901 |
| RW | 0.860 | −0.023 | 0.741 |
| RL | 0.965 | −0.005 | 0.931 |
| BuW | 0.805 | −0.266 | 0.719 |
| BL | 0.897 | 0.066 | 0.808 |
| ThP | 0.910 | −0.170 | 0.857 |
| ACP | 0.806 | −0.246 | 0.881 |
| PCP | 0.888 | −0.287 | 0.870 |
| BW | 0.961 | −0.004 | 0.923 |
| Total variance (%) | 73.80 | 7.01 | |
| Cumulative variance (%) | 73.80 | 80.81 | |
| Predicted/Actual Membership | Hidalgo | Maesso | Egea | López-Montenegro | Granda | Donoso |
|---|---|---|---|---|---|---|
| Hidalgo | 95.0 | 0 | 1.3 | 0 | 3.8 | 0 |
| Maesso | 0 | 96.7 | 0 | 3.3 | 0 | 0 |
| Egea | 0 | 0 | 70.7 | 4.9 | 24.4 | 0 |
| López-Montenegro | 0 | 1.5 | 12.1 | 80.3 | 6.1 | 0 |
| Granda | 5.0 | 0 | 18.3 | 15.0 | 61.7 | 0 |
| Donoso | 0 | 0 | 0 | 0 | 0 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Granero, A.; Anaya, G.; Alcalde, M.J. Morphostructural Differences between the Historical Genetic Lines of the Spanish Merino Sheep. Animals 2023, 13, 313. https://doi.org/10.3390/ani13020313
Granero A, Anaya G, Alcalde MJ. Morphostructural Differences between the Historical Genetic Lines of the Spanish Merino Sheep. Animals. 2023; 13(2):313. https://doi.org/10.3390/ani13020313
Chicago/Turabian StyleGranero, Antonio, Gabriel Anaya, and María J. Alcalde. 2023. "Morphostructural Differences between the Historical Genetic Lines of the Spanish Merino Sheep" Animals 13, no. 2: 313. https://doi.org/10.3390/ani13020313