
Seismic Signaling for Detection of Empty Tunnels in the Plateau Zokor, Eospalax baileyi
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
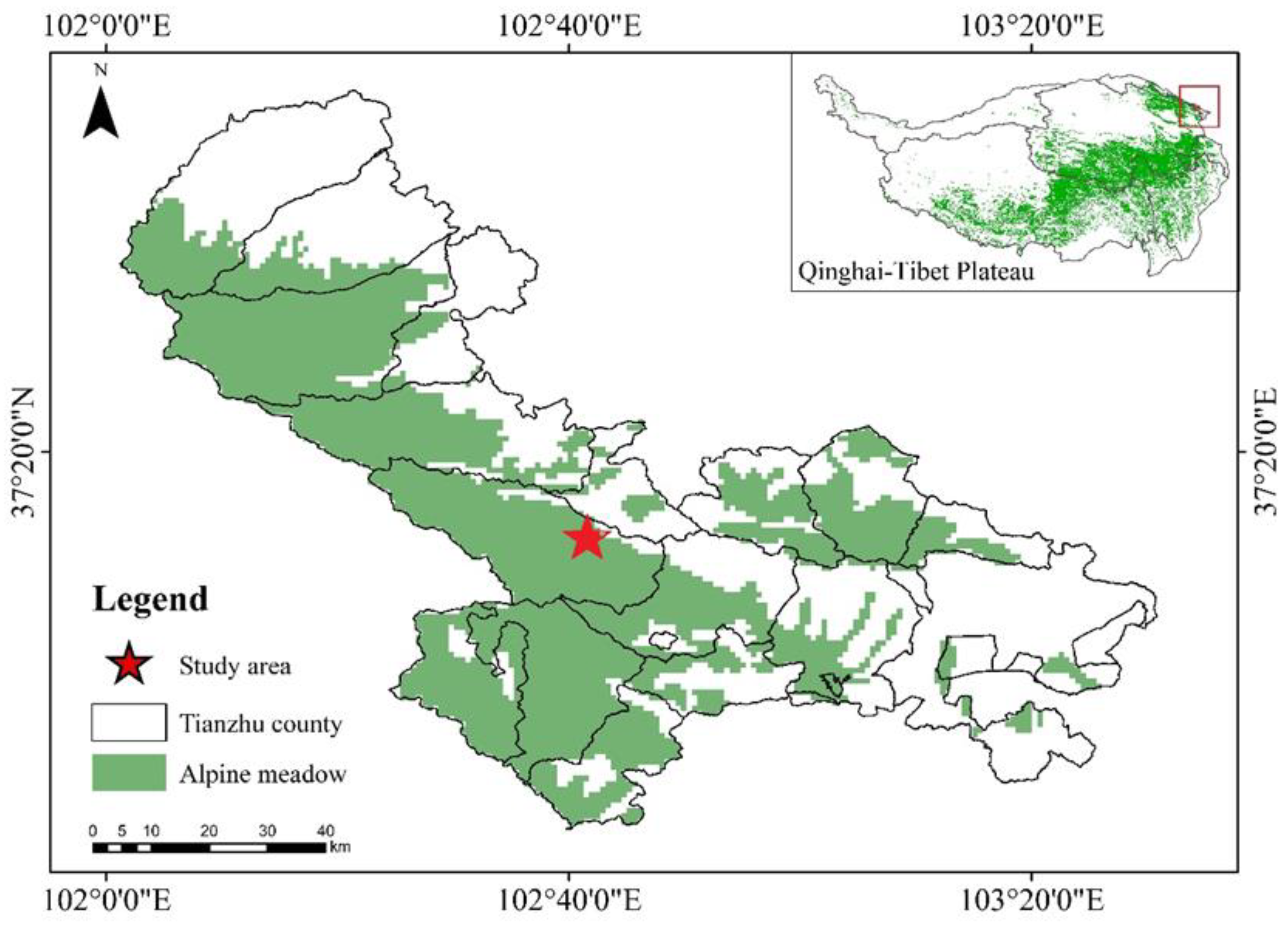
2.1. Study Site and Animals
2.2. Locating the Original Tunnels
2.3. Construction of the Artificial Tunnels
2.4. Recording of Seismic Signals in the Original Tunnel and during Occupation
2.5. Counting Occupations of the Artificial Tunnels
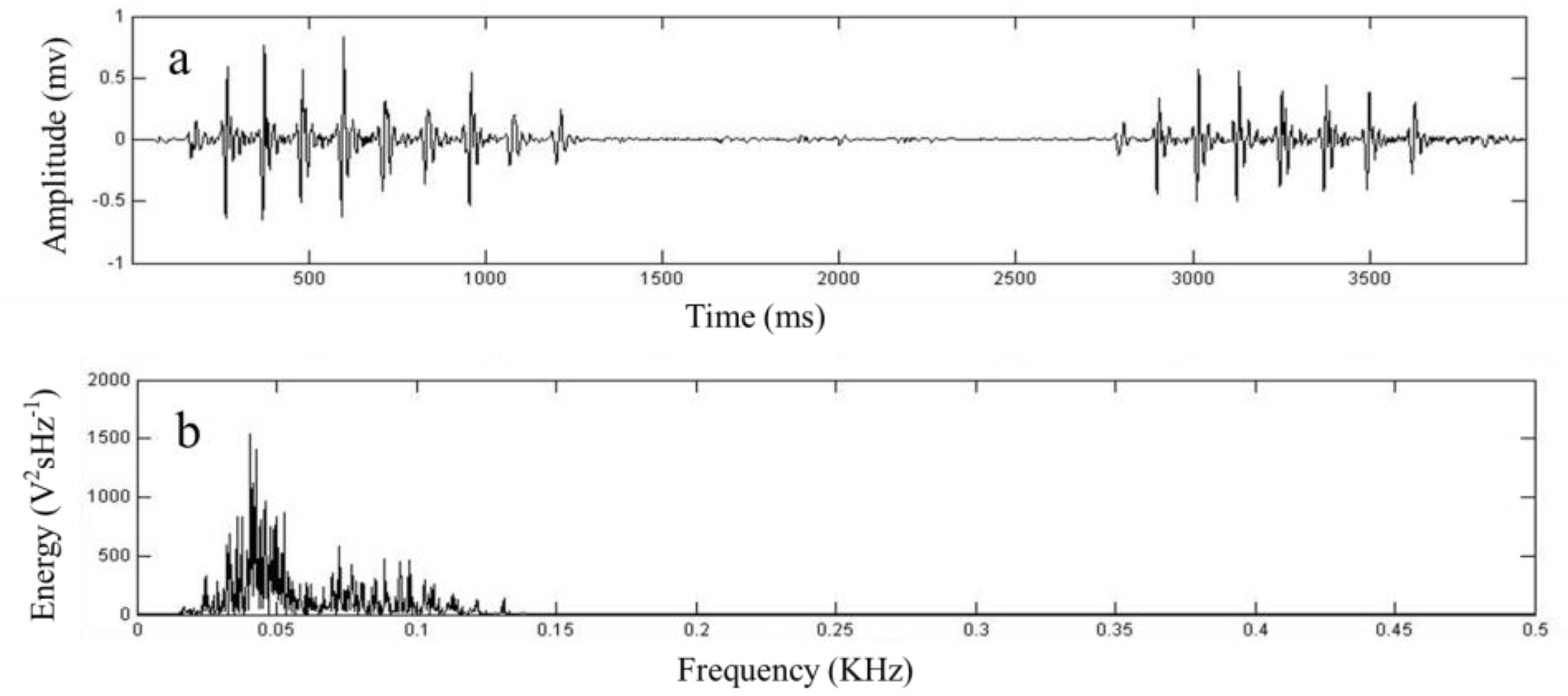
2.6. Definition of Measurement Parameters Related to Seismic Communication in the Study
2.7. Data Analysis
3. Results
3.1. Proportion of Occupied Tunnels among All Artificial Tunnels
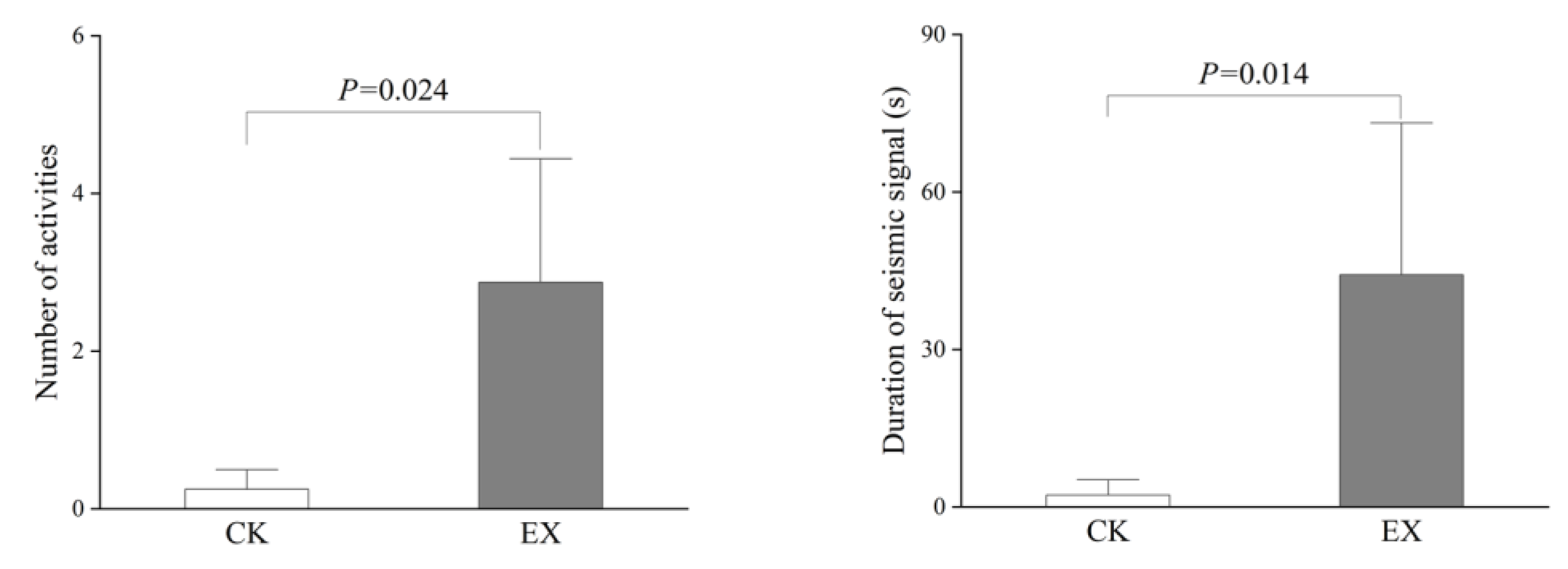
3.2. Number of Activities and the Duration of Seismic Signals Produced by Zokors in the Original Tunnel and When Artificial Tunnel Exists
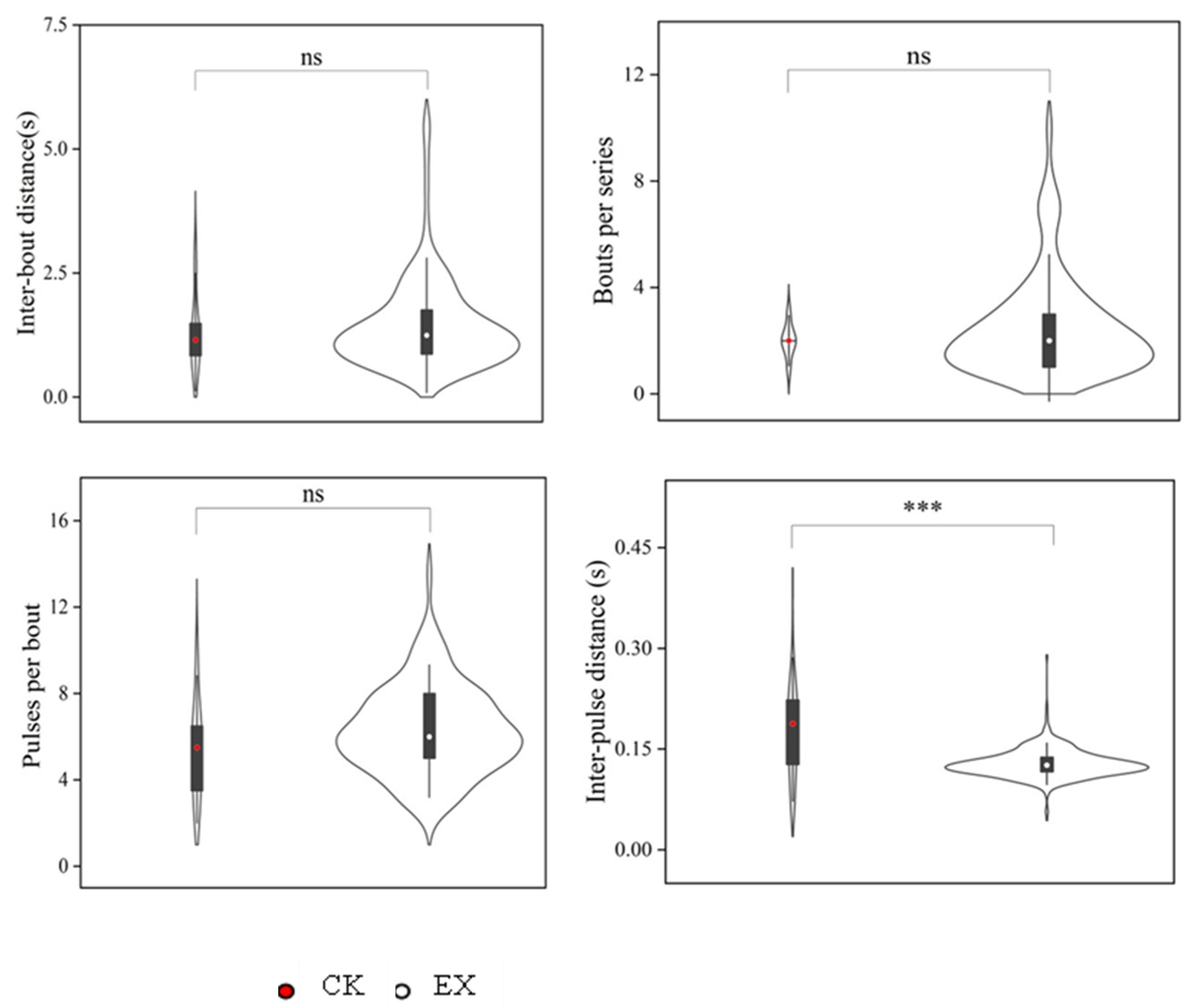
3.3. Differences in the Parameters of Seismic Signals of Plateau Zokors Recorded in the Original Tunnels and When Perceived by Artificial Tunnels
4. Discussion
4.1. Why Do Plateau Zokors Use Seismic Signals to Detect Neighboring Tunnels?
4.2. What Is the Mechanism Employed by Plateau Zokors When Occupying a Tunnel?
4.3. How Does the Plateau Zokor Perceive the Seismic Signals?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stamps, J.A. Territorial behavior: Testing the assumptions. J. Adv. Stud. Behav. 1994, 23, 232. [Google Scholar]
- He, P.; Maldonado-Chaparro, A.A.; Farine, D.R. The role of habitat configuration in shaping social structure: A gap in studies of animal social complexity. J. Behav. Ecol. Sociobiol. 2019, 73, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chapin, K.J.; Peixoto, P.E.C.; Briffa, M. Further mismeasures of animal contests: A new framework for assessment strategies. Behav. Ecol. 2019, 30, 1177–1185. [Google Scholar] [CrossRef]
- Pryke, S.R.; Andersson, S. Carotenoid-based status signalling in red-shouldered widowbirds (Euplectes axillaris): Epaulet size and redness affect captive and territorial competition. Behav. Ecol. Sociobiol. 2003, 53, 393–401. [Google Scholar] [CrossRef]
- Brown, J.L.; Orians, G.H. Spacing patterns in mobile animals. Annu. Rev. Ecol. Syst. 1970, 1, 239–262. [Google Scholar] [CrossRef]
- Goodale, E.; Beauchamp, G.; Magrath, R.D.; Nieh, J.C.; Ruxton, G.D. Interspecific information transfer influences animal community structure. Trends Ecol. Evol. 2010, 25, 354–361. [Google Scholar] [CrossRef]
- Searcy, W.A.; Nowicki, S. The Evolution of Animal Communication; Princeton University Press: Princeton, NJ, USA, 2010. [Google Scholar]
- Kubke, M.F.; Wild, J.M. Anatomy of vocal communication and hearing in rodents. In Rodent Bioacoustics; Springer: Cham, Switzerland, 2018; pp. 131–164. [Google Scholar]
- Pika, S.; Wilkinson, R.; Kendrick, K.H.; Vernes, S.C. Taking turns: Bridging the gap between human and animal communica-tion. Proc. R. Soc. B 2018, 285, 20180598. [Google Scholar] [CrossRef] [Green Version]
- Begall, S.; Burda, H.; Schleich, C.E. Subterranean rodents: News from underground. In Subterranean Rodents; Springer: Berlin/Heidelberg, Germany, 2007; pp. 3–9. [Google Scholar]
- Wilson, D.E.; Reeder, D.M. (Eds.) Mammal Species of the World: A Taxonomic and Geographic Reference; JHU Press: Baltimore, MD, USA, 2005; Volume 1. [Google Scholar]
- Nevo, E. Adaptive convergence and divergence of subterranean mammals. Annu. Rev. Ecol. Syst. 1979, 10, 269–308. [Google Scholar] [CrossRef]
- Zhang, F.Y.; Zhou, J.W.; Hua, L.M. A review on the seismic signal in fossorial rodents. Sichuan J. Zool. 2019, 38, 464–471. [Google Scholar]
- Hill, P.S.M. Vibrational Communication in Animals; Harvard University Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Kashash, Y.; Smarsh, G.; Zilkha, N.; Yovel, Y.; Kimchi, T. Alone, in the dark: The extraordinary neuroethology of the solitary blind mole rat. eLife 2022, 11, e78295. [Google Scholar] [CrossRef]
- Burda, H. Sensory perception of mole-rats and mole rats: Assessment of a complex natural global evolutionary “experi-ment”. In New Horizons in Evolution; Academic Press: Cambridge, MA, USA, 2021; pp. 161–191. [Google Scholar]
- Randall, J.A.; King, D.K.B. Assessment and defence of solitary kangaroo rats under risk of predation by snakes. Anim. Behav. 2001, 61, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Bednářová, R.; Hrouzková-Knotková, E.; Burda, H.; Sedláček, F.; Šumbera, R. Vocalizations of the giant mole-rat (Fukomys mechowii), a subterranean rodent with the richest vocal repertoire. Bioacoustics 2013, 22, 87–107. [Google Scholar] [CrossRef]
- Hill, P.S.M. Vibration and animal communication: A review. Am. Zool. 2001, 41, 1135–1142. [Google Scholar] [CrossRef] [Green Version]
- Lacey, E.A. Life Underground: The Biology of Subterranean Rodents; University of Chicago Press: Chicago, IL, USA, 2000. [Google Scholar]
- Rado, R.; Levi, N.; Hauser, H.; Witcher, J.; Alder, N.; Intrator, N.; Wollberg, Z.; Terkel, J. Seismic signalling as a means of communication in a subterranean mammal. Anim. Behav. 1987, 35, 1249–1251. [Google Scholar] [CrossRef]
- Giannoni, S.M.; Márquez, R.; Borghi, C.E. Airborne and substrate-borne communications of Microtus (Terricola) gerbei and M.(T.) duodecimcostatus. Acta Theriol. 1997, 42, 123–141. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Xu, M.; Ren, S.E.; Liang, N.N.; Han, C.X.; Nan, X.N.; Shi, M.N. Taxonomy and phylogenetic relationship of zokors. J.Genet. 2020, 99, 38. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, S.Z.; Cai, Z.Y.; Kuang, Z.R.; Wan, N.; Wang, Y.J.; Mao, L.Y.; An, X.; Li, F.; Feng, T.; et al. Genomic insights into zokors’ phylogeny and speciation in China. Proc. Natl. Acad. Sci. USA 2022, 119, e2121819119. [Google Scholar] [CrossRef]
- Niu, Y.J.; Yang, S.W.; Zhu, H.M.; Zhou, J.W.; Chu, B.; Ma, S.J.; Hua, R.; Hua, L.M. Cyclic formation of zokor mounds promotes plant diversity and renews plant communities in alpine meadows on the Tibetan Plateau. Plant Soil 2020, 446, 65–79. [Google Scholar] [CrossRef]
- Chu, B.; Ye, G.H.; Yang, S.W.; Zhou, F.F.; Zhang, F.Y.; Zhou, J.W.; Hua, L.M. Effect of plateau zokor (Myospalax fontanierii) disturbance on plant community structure and soil properties in the eastern Qinghai-Tibet Plateau, China. Rangel. Ecol. Manag. 2020, 73, 520–530. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, C.Y.; Zheng, X.H.; Fu, Y.F.; Hu, X.X.; Cao, G.M.; Butterbach-Bahl, K. The increasing distribution area of zokor mounds weaken greenhouse gas uptakes by alpine meadows in the Qinghai–Tibetan Plateau. Soil Biol. Biochem. 2014, 71, 105–112. [Google Scholar] [CrossRef]
- Fan, N.C.; Jing, Z.C.; Zhou, W.Y. On the invading behavior of plateau zokor and a new approach to its control. Acta Theriol. Sin. 1990, 10, 114–120. [Google Scholar] [CrossRef]
- Kimchi, T.; Reshef, M.; Terkel, J. Evidence for the use of reflected self-generated seismic waves for spatial orientation in a blind subterranean mammal. J. Exp. Biol. 2005, 208, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, J. Mole Catching: A Practical Guide; Crowood: Marlborough, UK, 2012. [Google Scholar]
- Zhou, J.W.; Zhang, F.Y.; Chu, B.; Dong, K.C.; Hua, L.M. Study on seismic communication mode and signal in plateau zokor. Grassl. Turf 2020, 40, 54–60. [Google Scholar]
- Nevo, E.; Heth, G.; Pratt, H. Seismic communication in a blind subterranean mammal: A major somatosensory mechanism in adaptive evolution underground. Proc. Natl. Acad. Sci. USA 1991, 88, 1256–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, C.P. Study on the Warm Season Activity Rhythm and Nest Area Change of Plateau Zokor in the Eastern Qilian Mountains; Gansu Agricultural University: Lanzhou, China, 2017. [Google Scholar]
- Dong, K.C.; Shi, J.J.; Zhou, J.W.; Hua, L.M. Observation of seismic generating body parts and analysis of signal characteristics of plateau zokor(Moyspalax baileyi). Grassl. Turf 2021, 41, 49–54. [Google Scholar] [CrossRef]
- Zhang, Y.M. The biology and ecology of plateau zokors (Eospalax fontanierii). In Subterranean Rodents; Springer: Berlin/Heidelberg, Germany, 2007; pp. 237–249. [Google Scholar]
- Narins, P.M.; Reichman, O.J.; Jarvis, J.U.; Lewis, E.R. Seismic signal transmission between burrows of the Cape mole-rat, Georychus capensis. J. Comp. Physiol. A 1992, 170, 13–21. [Google Scholar] [CrossRef]
- Li, J.G.; Wang, T.Z.; He, J.P.; Min, Y.J. Seismic communication in subterranean Gansu zokor (Myospalax cansus). Acta Theriol. Sin. 2001, 21, 152–154. [Google Scholar]
- Hegab, I.M.; Tan, Y.C.; Wang, C.; Yao, B.H.; Wang, H.F.; Ji, W.H.; Su, J.H. Examining object recognition and object-in-Place memory in plateau zokors, Eospalax baileyi. Behav. Process. 2018, 146, 34–41. [Google Scholar] [CrossRef]
- Rado, R.; Himelfarb, M.; Arensburg, B.; Terkel, J.; Wollberg, Z. Are seismic communication signals transmitted by bone con-duction in the blind mole rat? Hear. Res. 1989, 41, 23–29. [Google Scholar] [CrossRef]
- Lange, S.; Burda, H.; Wegner, R.E.; Dammann, P.; Begall, S.; Kawalika, M. Living in a “stethoscope”: Burrow-acoustics promote auditory specializations in subterranean rodents. Naturwissenschaften 2007, 94, 134–138. [Google Scholar] [CrossRef]
- Mason, M.J.; Lai, F.W.S.; Li, J.G.; Nevo, E. Middle ear structure and bone conduction in Spalax, Eospalax, and Tachyoryctes mole-rats (Rodentia: Spalacidae). J. Morphol. 2010, 271, 462–472. [Google Scholar] [PubMed]
- Xu, L.N.; Wei, L.N.; Wang, Y.; Li, X.; Wei, L.; Wei, D.B. The relationship between the structure of the prestin gene and specific expression in tissues of the plateau zokor and its adaption to the underground life. Acta Theriol. Sin. 2016, 36, 221. [Google Scholar] [CrossRef]
- Wei, W.H.; Wang, Q.Y.; Zhou, W.Y.; Fan, N.C. The population dynamics and dispersal of plateau zokor after removing. Acta Theriol. Sin. 1997, 17, 53–61. [Google Scholar]
- Zhang, J.J.; Zhang, Z.B. Animal Mating Systems. Chin. J. Zool. 2003, 2, 84–89. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Explanation |
|---|---|
| inter-pulse time interval | The duration of the silence between two successive pulses. |
| pulses per bout | The number of thumps (pulses) in a bout. |
| inter-bout time interval | The duration of the silence between two successive bouts. |
| bouts per series | Number of bouts in a series. |
| number of activities | The number of the series. |
| duration of seismic signals | The duration of the series. |
| peak frequency | Seismic signal amplitude FFT—Spectrogram abscissa value. |
| energy | Seismic signal amplitude FFT—Spectrogram ordinate value. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, K.; Zhou, J.; Zhang, F.; Dong, L.; Chu, B.; Hua, R.; Hua, L. Seismic Signaling for Detection of Empty Tunnels in the Plateau Zokor, Eospalax baileyi. Animals 2023, 13, 240. https://doi.org/10.3390/ani13020240
Dong K, Zhou J, Zhang F, Dong L, Chu B, Hua R, Hua L. Seismic Signaling for Detection of Empty Tunnels in the Plateau Zokor, Eospalax baileyi. Animals. 2023; 13(2):240. https://doi.org/10.3390/ani13020240
Chicago/Turabian StyleDong, Kechi, Jianwei Zhou, Feiyu Zhang, Longming Dong, Bin Chu, Rui Hua, and Limin Hua. 2023. "Seismic Signaling for Detection of Empty Tunnels in the Plateau Zokor, Eospalax baileyi" Animals 13, no. 2: 240. https://doi.org/10.3390/ani13020240