
A New AS-PCR Method to Detect CSN201 Allele, Genotyping at Ca-Sensitive Caseins Loci and Milk Traits Association Studies in Autochthonous Lazio Goats
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Farms and Animals
2.2. DNA Extraction
2.3. Genotyping at the CSN1S1 Locus
2.4. Genotyping at the CSN1S2 Locus
2.5. Genotyping at the CSN2 Locus
2.6. Milk Analyses
- -
- Milkoscan FT 6000 (the i.r. spectrophotometry method for: fat, protein, total caseins, lactose, solids-not-fat, and urea, according to the International Dairy Federation (IDF) standard 141:2013 (ISO-IDF, 2013) [28].
- -
- Fossomatic FT 5000 (the fluoro-optoelectronic method) for somatic cells, according to the IDF 148–2:2006 method (ISO-IDF, 2006) [29].
2.7. Statistical Analysis
3. Results
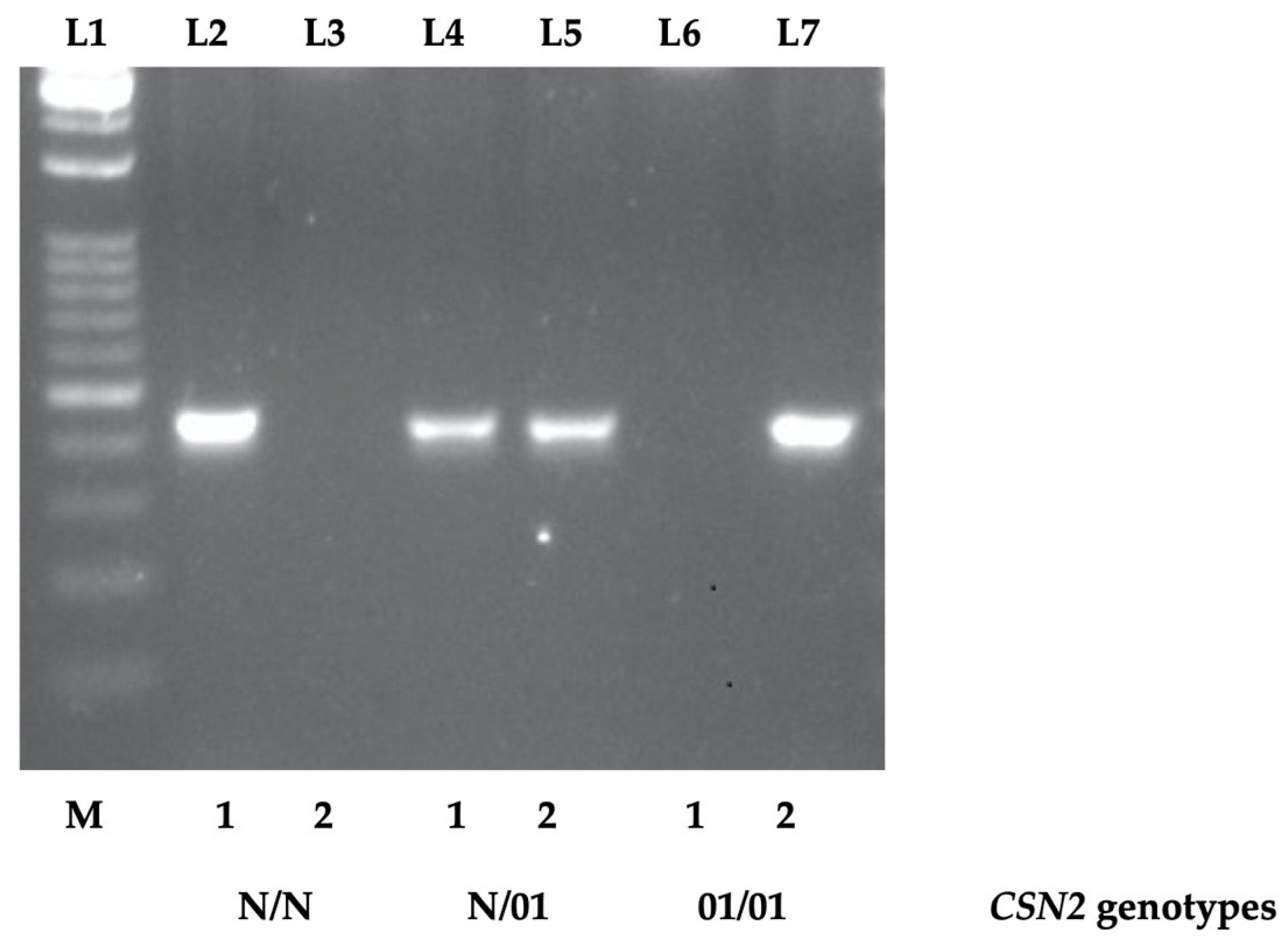
3.1. A New AS-PCR for the CSN201 Allele Detection
3.2. Genotyping
3.3. Allele Effect on Milk Parameters
4. Discussions
4.1. AS-PCR Protocol for CSN201 Allele Detection
4.2. Milk Traits Phenotyping
4.3. Calcium-Sensitive Caseins Loci Genotyping and Population Genetic Structure
4.4. Association Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Albarella, S.; Selvaggi, M.; D’Anza, E.; Cosenza, G.; Caira, S.; Scaloni, A.; Fontana, A.; Peretti, V.; Ciotola, F. Influence of the Casein Composite Genotype on Milk Quality and Coagulation Properties in the Endangered Agerolese Cattle Breed. Animals 2020, 10, 892. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, G.; Pauciullo, A.; Gallo, D.; Colimoro, L.; D’Avino, A.; Mancusi, A.; Ramunno, L. Genotyping at the CSN1S1 locus by PCR-RFLP and AS-PCR in a Neapolitan Goat Population. Small Rumin. Res. 2008, 74, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Caroli, A.; Chiatti, F.; Chessa, S.; Rignanese, D.; Ibeagha-Awemu, E.M.; Erhardt, G. Characterization of the casein gene complex in West African goats and description of a new αS1-casein polymorphism. J. Dairy Sci. 2007, 90, 2989–2996. [Google Scholar] [CrossRef] [PubMed]
- Garro, G.; Ferranti, P.; De Pascale, S.; Nicolai, M.A.; Mauriello, R.; Quarto, M.; Pilla, F.; Chianese, L. The occurrence of genetic polymorphism and related non-allelic proteins increases the composition complexity of goat αS1-CN. Electrophoresis 2012, 33, 2337–2344. [Google Scholar] [CrossRef]
- Mangia, N.P.; Saliba, L.; Zoumpopoulou, G.; Chessa, S.; Anastasiou, R.; Karayiannis, Ι.; Sgouras, D.; Tsakalidou, E.; Nudda, A. Goat Milk with Different Alpha-s1 Casein Genotype (CSN1S1) Fermented by Selected Lactobacillus paracasei as Potential Functional Food. Fermentation 2019, 5, 55. [Google Scholar] [CrossRef] [Green Version]
- Mestawet, T.A.; Girma, A.; Ådnøy, T.; Devold, T.G.; Vegarud, G.E. Newly identified mutations at the CSN1S1 gene in Ethiopian goats affect casein content and coagulation properties of their milk. J. Dairy Sci. 2013, 96, 4857–4869. [Google Scholar] [CrossRef] [Green Version]
- Ramunno, L.; Cosenza, G.; Rando, A.; Pauciullo, A.; Illario, R.; Gallo, D.; Di Berardino, D.; Masina, P. Comparative analysis of gene sequence of goat CSN1S1 F and N alleles and characterization of CSN1S1 transcript variants in mammary gland. Gene 2005, 345, 289–299. [Google Scholar] [CrossRef]
- Devold, T.G.; Nordbø, R.; Langsrud, T.; Svenning, C.; Brovold, M.J.; Sørensen, E.S.; Christensen, B.; Ådnøy, T.; Vegarud, G.E. Extreme frequencies of the αs1-casein “null” variant in milk from Norwegian dairy goats—Implications for milk composition, micellar size and renneting properties. Dairy Sci. Technol. 2011, 91, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Cosenza, G.; Pauciullo, A.; Gallo, D.; Di Berardino, D.; Ramunno, L. A Ssp I PCR-RFLP detecting a silent allele at the goat CSN2 locus. J. Dairy Res. 2005, 72, 456–459. [Google Scholar] [CrossRef] [Green Version]
- Cosenza, G.; Iannaccone, M.; Pico, B.A.; Ramunno, L.; Capparelli, R. The SNP g.1311T>C associated with the absence of β-casein in goat milk influences CSN2 promoter activity. Anim. Genet. 2016, 47, 615–617. [Google Scholar] [CrossRef]
- Nicolai, M.A.; Garro, G.; Caira, S.; Mauriello, R.; Quarto, M.; De Pascale, S.; Chianese, L. Occurrence of quantitative genetic polymorphism at the caprine β-CN locus, as determined by a proteomic approach. Int. Dairy J. 2021, 112, 104855. [Google Scholar] [CrossRef]
- Erhardt, G.; Jäger, S.; Budelli, E.; Caroli, A. Genetic polymorphism of goat αS2-casein (CSN1S2) and evidence for a further allele. Milchwissenschaft 2002, 57, 137–140. [Google Scholar]
- Lagonigro, R.; Pietrolà, E.; D’Andrea, M.; Veltri, C.; Pilla, F. Molecular genetic characterization of the goat αs2-casein E allele. Anim. Genet. 2001, 32, 391–393. [Google Scholar] [CrossRef] [PubMed]
- Ramunno, L.; Cosenza, G.; Pappalardo, M.; Longobardi, E.; Gallo, D.; Pastore, N.; Di Gregorio, P.; Rando, A. Characterization of two new alleles at the goat CSN1S2 locus. Anim. Genet. 2001, 32, 264–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmatalla, S.A.; Arends, D.; Ahmed, A.S.; Hassan, L.M.A.; Krebs, S.; Reissmann, M.; Gudrun, A.; Brockmann, G.A. Capture Capture Sequencing to Explore and Map Rare Casein Variants in Goats. Front. Genet. 2021, 12, 620253. [Google Scholar] [CrossRef] [PubMed]
- Jansà-Perez, M.; Leroux, C.; Bonastre, A.S.; Martin, P. Occurrence of a Line sequence in the 3′ UTR of the goat αs1-casein E-encoding allele associated with reduced protein synthesis level. Gene 1994, 147, 179–187. [Google Scholar] [CrossRef]
- Ramunno, L.; Cosenza, G.; Pappalardo, M.; Pastore, N.; Gallo, D.; Di Gregorio, P.; Masina, P. Identification of the goat CSN1S1F allele by means of PCR-RFLP method. Anim. Genet. 2000, 31, 342–343. [Google Scholar] [CrossRef]
- Cosenza, G.; Illario, R.; Rando, A.; Di Gregorio, P.; Masina, P.; Ramunno, L. Molecular characterization of the goat CSN1S101 allele. J. Dairy Res. 2003, 70, 237–240. [Google Scholar] [CrossRef]
- Bevilacqua, C.; Martin, P.; Candalh, C.; Fauquant, J.; Piot, M.; Bouvier, F.F.; Manfredi, E.; Pilla, F.; Heyman, M. Allergic sensitisation to milk proteins in guinea-pigs fed cow milk and goat milks of different genotypes. In Proceedings of the VIIth International Conference on Goat, Tours, France, 15–21 May 2000. [Google Scholar]
- Marletta, D.; Bordonaro, S.; Guastella, A.M.; Falagiani, P.; Crimi, N.; D’Urso, G. Goat milk with different αS2-casein content: Analysis of allergenic potency by REAST-inhibition assay. Small Rumin. Res. 2004, 52, 19–24. [Google Scholar] [CrossRef]
- Martin, P.; Szymanowska, M.; Zwierzchowski, L.; Leroux, C. The impact of genetic polymorphisms on the protein composition of ruminant milks. Reprod. Nutr. Dev. 2002, 42, 433–459. [Google Scholar] [CrossRef]
- Selvaggi, M.; Tufarelli, V. Caseins of goat and sheep milk: Analytical and technological aspects. In Casein: Production, Uses and Health Effects; Nova Science Publisher: Hauppage, NY, USA, 2012; pp. 1–26. [Google Scholar]
- Selvaggi, M.; Laudadio, V.; Cataldo, D.; Tufarelli, V. Major proteins in goat milk: An updated overview on genetic variability. Mol. Biol. Rep. 2014, 41, 1035–1048. [Google Scholar] [CrossRef] [PubMed]
- Tortorici, L.; Di Gerlando, R.; Mastrangelo, S.; Sardina, M.T.; Portolano, B. Genetic Characterisation of CSN2 Gene in Girgentana Goat Breed. Ital. J. Anim. Sci. 2014, 13, 3414. [Google Scholar] [CrossRef]
- Vacca, G.M.; Dettori, M.L.; Piras, G.; Manca, F.; Paschino, P.; Pazzola, M. Goat casein genotypes are associated with milk production traits in the Sarda breed. Anim. Genet. 2014, 45, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.P.; Zhang, X.M.; Wang, W.; Ma, R.N.; Deng, C.J.; Lan, X.Y.; Chen, H.; Li, F.; Xu, X.R.; Ma, Y.; et al. The CSN1S1 N and F alleles identified by PCR–SSCP and their associations with milk yield and composition in Chinese dairy goats. Mol. Biol. Rep. 2011, 38, 2821–2825. [Google Scholar] [CrossRef]
- Zullo, A.; Barone, C.M.A.; Chianese, L.; Colatruglio, P.; Occidente, M.; Matassino, D. Protein polymorphisms and coagulation properties of Cilentana goat milk. Small Rumin. Res. 2005, 58, 223–230. [Google Scholar] [CrossRef]
- ISO 9622:2013 | IDF 141:2013; Milk and Liquid Milk Products. Determination of Fat, Protein, Casein, Lactose and pH Content. ISO—International Organization for Standardization: Geneva, Switzerland; IDF—International Dairy Federation: Brussels, Belgium, 2013.
- ISO 13366-2:2006 | IDF 148-2:2006; Milk. Enumeration of Somatic Cells. Part 2: Guidance on the Operation of Fluoro-Opto-Electronic Counters. ISO—International Organization for Standardization: Geneva, Switzerland; IDF—International Dairy Federation: Brussels, Belgium, 2006.
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Longman Group Ltd.: Essex, UK, 1996. [Google Scholar]
- Yeh, F.C.; Yang, R.; Boyle, T.J.; Ye, Z.; Xiyan, J.M. PopGene32, Microsoft Windows-based freeware for population genetic analysis; Version 1.32; University of Alberta: Edmonton, Canada, 2000. [Google Scholar]
- Botstein, D.; White, R.L.; Skalnick, M.H.; Davies, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphism. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Littell, R.C.; Henry, P.R.; Ammerman, C.B. Statistical analysis of repeated measures data using SAS procedures. J. Anim. Sci. 1998, 76, 1216–1231. [Google Scholar] [CrossRef] [Green Version]
- Boulanger, A.; Grosclaude, F.; Mahé, M.F. Polymorphisme des caséines α(s1) et α(s2) de la chèvre (Capra hircus). Genet. Sel. 1984, 16, 157–176. [Google Scholar] [CrossRef] [Green Version]
- Dall’Olio, S.; Davoli, R.; Russo, V. Una nuova variante di caseina caprina. Sci. Tecnol. Latt.-Casearia 1989, 40, 24–28. [Google Scholar]
- Ramunno, L.; Mariani, P.; Pappalardo, M.; Rando, A.; Capuano, M.; Di Gregorio, P.; Cosenza, G. Un gene ad effetto maggiore sul contenuto di caseina β nel latte di capra. In Proceedings of the Atti XI Congresso Nazionale ASPA, Grado (GO), Palermo, Italy, 19–22 June 1995. [Google Scholar]
- Currò, S.; Manuelian, C.L.; De Marchi, M.; Goi, A.; Claps, S.; Esposito, L.; Neglia, G. Italian local goat breeds have better milk coagulation properties than cosmopolitan breed. Ital. J. Anim. Sci. 2020, 19, 593–601. [Google Scholar] [CrossRef]
- Ferro, M.M.; Tedeschi, L.O.; Atzori, A.S. The comparison of the lactation and milk yield and composition of selected breeds of sheep and goats. Transl. Anim. Sci. 2017, 1, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Currò, S.; Manuelian, C.L.; De Marchi, M.; De Palo, P.; Claps, S.; Maggiolino, A.; Campanile, G.; Rufrano, D.; Fontana, A.; Pedota, G.; et al. Autochthonous dairy goat breeds showed better milk quality than Saanen under the same environmental conditions. Arch. Anim. Breed. 2019, 62, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Todaro, M.; Scatassa, M.L.; Giaccone, P. Multivariate factor analysis of Girgentana goat milk composition. Ital. J. Anim. Sci. 2005, 4, 403–410. [Google Scholar] [CrossRef]
- Marletta, D.; Bordonaro, S.; Guastella, A.M.; Criscione, A.; D’Urso, G. Genetic polymorphism of the calcium sensitive caseins in Sicilian Girgentana and Argentata dell’Etna goat breeds. Small Rumin. Res. 2005, 57, 133–139. [Google Scholar] [CrossRef]
- Sacchi, P.; Chessa, S.; Budelli, E.; Bolla, P.; Ceriotti, G.; Soglia, D.; Rasero, R.; Cauvin, E.; Caroli, A. Casein haplotype structure in five Italian goat breeds. J. Dairy Sci. 2005, 88, 1561–1568. [Google Scholar] [CrossRef] [Green Version]
- Pazzola, M.; Dettori, M.L.; Pira, E.; Noce, A.; Paschino, P.; Vacca, G.M. Effect of polymorphisms at the casein gene cluster on milk renneting properties of the Sarda goat. Small Rumin. Res. 2014, 117, 124–130. [Google Scholar] [CrossRef]
- Deniz, D.; Sena, A.; Hale, S.; Buse, V.; Faruk, B. Identification of the frequency of CSN1S2 gene alleles and the effects of these alleles and parity on milk yield and composition in Saanen goats. Large Anim. Revi. 2021, 27, 91–96. [Google Scholar]
- Kusza, S.; Veress, G.; Kukovics, S.; Jávor, A.; Sanchez, A.; Angiolillo, A.; Bösze, Z. Genetic polymorphism of αs1- and αs2- caseins in Hungarian milking goats. Small Rumin. Res. 2007, 68, 329–332. [Google Scholar] [CrossRef]
- Caroli, A.; Chiatti, F.; Chessa, S.; Rignanese, D.; Bolla, P.; Pagnacco, G. Focusing on the goat casein complex. J. Dairy Sci. 2006, 89, 3178–3187. [Google Scholar] [CrossRef] [Green Version]
- Chessa, S.; Budelli, E.; Chiatti, F.; Cito, A.M.; Bolla, P.; Caroli, A. Short Communication: Predominance of β-Casein (CSN2) C Allele in Goat Breeds Reared in Italy. J. Dairy Sci. 2005, 88, 1878–1881. [Google Scholar] [CrossRef] [Green Version]
- Persuy, M.A.; Printz, C.; Medrano, J.F.; Mercier, J.C. A single nucleotide deletion resulting in a premature stop codon is associated with marked reduction of transcripts from a goat β-casein null allele. Anim. Genet. 1999, 30, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Frattini, S.; Nicoloso, L.; Coizet, B.; Chessa, S.; Rapetti, L.; Pagnacco, G.; Crepaldi, P. Short communication: The unusual genetic trend of αS1-casein in Alpine and Saanen breeds. J. Dairy Sci. 2014, 97, 7975–7979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avondo, M.; Pagano, R.I.; Guastella, A.M.; Criscione, A.; Di Gloria, M.; Valenti, B.; Piccione, G.; Pennisi, P. Diet selection and milk production and composition in Girgentana goats with different αs1-casein genotype. J. Dairy Res. 2009, 76, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Balia, F.; Pazzola, M.; Dettori, M.L.; Mura, M.C.; Luridiana, S.; Carcangiu, V.; Piras, G.; Vacca, G.M. Effect of CSN1S1 gene polymorphism and stage of lactation on milk yield and composition of extensively reared goats. J. Dairy Res. 2013, 80, 129–137. [Google Scholar] [CrossRef]
- Chilliard, Y.; Rouel, J.; Leroux, C. Goats alpha-s1-casein genotype influences its milk fatty acid composition and delta-9 desaturation ratios. Anim. Feed Sci. Technol. 2006, 131, 474–487. [Google Scholar] [CrossRef]
- Pagano, R.I.; Pennisi, P.; Valenti, B.; Lanza, M.; Di Trana, A.; Di Gregorio, P.; De Angelis, A.; Avondo, M. Effect of CSN1S1 genotype and its interaction with diet energy level on milk production and quality in Girgentana goats fed ad libitum. J. Dairy Res. 2010, 77, 245–251. [Google Scholar] [CrossRef]
- Schmidely, P.; Meschy, F.; Tessier, J.; Sauvant, D. Lactation response and nitrogen, calcium, and phosphorus utilization of dairy goats differing by the genotype for αs1-casein in milk, and feed diets varying in crude protein concentration. J. Dairy Sci. 2002, 85, 2299–2307. [Google Scholar] [CrossRef] [Green Version]
- Carillier-Jacquin, C.; Larroque, H.; Robert-Granié, C. Including αs1 casein gene information in genomic evaluations of French dairy goats. Genet. Sel. Evol. 2016, 48, 54. [Google Scholar] [CrossRef] [Green Version]
- Santillo, A.; Ciliberti, M.G.; D’Angelo, F.; Albenzio, M. The Effect of Alpha s1 Genotype on Some Physiological and Chemical Milk Characteristics in Garganica Goat. Anim. Front. 2022, 3, 897172. [Google Scholar] [CrossRef]
- Montalbano, M.; Tortorici, L.; Mastrangelo, S.; Sardina, M.T.; Portolano, B. Development and validation of RP-HPLC method for the quantitative estimation of αs1-genetic variants in goat milk. Food Chem. 2014, 156, 165–169. [Google Scholar] [CrossRef] [Green Version]
- Turhan Dinçel, D.; Ardicli, S.; Samli, H.; Balci, F. Determining the frequencies of B1, B2, B3 and E alleles of the CSN1S1 gene and their effects on milk yield and composition in Saanen goats. S. Afr. J. Anim. Sci. 2016, 46, 180–190. [Google Scholar] [CrossRef] [Green Version]
- Cieslak, J.; Wodas, L.; Borowska, A.; Pawlak, P.; Czyzak-Runowska, G.; Wojtowski, J.; Puppel, K.; Kuczynska, B.; Mackowski, M. 5′-flanking variants of equine casein genes (CSN1S1, CSN1S2, CSN2, CSN3) and their relationship with gene expression and milk composition. J. Appl. Genet. 2019, 60, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prinzenberg, E.M.; Weimann, C.; Brandt, H.; Bennewitz, J.; Kalm, E.; Schwerin, M.; Erhardt, G. Polymorphism of the bovine CSN1S1 promoter: Linkage mapping, intragenic haplotypes, and effects on milk production traits. J. Dairy Sci. 2003, 86, 2696–2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, K.; Bennewitz, J.; Reinsch, N.; Thaller, G.; Prinzenberg, E.M.; Kühn, C.; Kalm, E. Characterization of the DGAT1 mutations and the CSN1S1 promoter in the German Angeln dairy cattle population. J. Dairy Sci. 2006, 89, 3164–3174. [Google Scholar] [CrossRef] [PubMed]
- Kuss, A.W.; Gogol, J.; Bartenschlager, H.; Geldermann, H. Polymorphic AP-1 binding site in bovine CSN1S1 shows quantitative differences in protein binding associated with milk protein expression. J. Dairy Sci. 2005, 88, 2246–2252. [Google Scholar] [CrossRef] [Green Version]
- Mastrangelo, S.; Sardina, M.T.; Tolone, M.; Portolano, B. Genetic polymorphism at the CSN1S1 gene in Girgentana dairy goat breed. Anim. Prod. Sci. 2013, 53, 403–406. [Google Scholar] [CrossRef] [Green Version]
- Guastella, A.M.; Criscione, A.; Zuccaro, A.; Tidona, F.; Marietta, D.; Bordonaro, S. Genetic polymorphism of CSN1S1 and CSN2 loci in Rossa Mediterranea goat population. Ital. J. Anim. Sci. 2009, 8, 101–103. [Google Scholar] [CrossRef]


{kind=link}
| Breed | Genotype Numbers | Allele Frequency | Population Indices | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A*A* | A*B* | B*B* | FF | B*F | A*F | FN | A* | B* | F | N | Ho | He | FIS | ||
| Capestrina n = 27 | Obs | 1 | 2 | 2 | 9 | 8 | 5 | 0 | 0.170 | 0.260 | 0.570 | 0 | 0.42 | 0.58 | 0.14 |
| Exp | 0.75 | 2.33 | 1.81 | 8.90 | 8.04 | 5.17 | 0 | ||||||||
| χ2 = 0.16 p = 0.01 d.f. = 5 | |||||||||||||||
| Bianca Monticellana n = 125 | Obs | 15 | 18 | 9 | 24 | 35 | 23 | 1 | 0.284 | 0.284 | 0.428 | 0.004 | 0.61 | 0.66 | 0.06 |
| Exp | 10.08 | 20.16 | 10.08 | 22.90 | 30.39 | 30.39 | 0.43 | ||||||||
| χ2 = 6.06 p = 0.01 d.f. = 6 | |||||||||||||||
| Ciociara Grigia n = 36 | Obs | 1 | 6 | 5 | 3 | 12 | 9 | 0 | 0.240 | 0.390 | 0.380 | 0 | 0.75 | 0.66 | -0.14 |
| Exp | 2.01 | 6.61 | 5.44 | 5.06 | 10.50 | 6.38 | 0 | ||||||||
| χ2 = 2.73 p = 0.01 d.f. = 5 | |||||||||||||||
| All breeds n = 188 | Obs | 17 | 26 | 16 | 36 | 55 | 37 | 1 | 0.26 | 0.30 | 0.44 | 0.003 | 0.63 | 0.65 | 0.03 |
| Exp | 12.51 | 29.15 | 16.98 | 36.20 | 49.59 | 42.57 | 0.44 | ||||||||
| χ2 = 4.04 p = 0.01 d.f. = 6 | |||||||||||||||
| Parameters | Genotypes | |||||
|---|---|---|---|---|---|---|
| A*A* n = 16 | A*B* n = 24 | B*B* n = 15 | A*F n = 34 | B*F n = 44 | FF n = 35 | |
| Milk yield (g/milking) | 608.13 ± 66.26 | 521.54 ± 51.98 | 507.14 ± 70.83 | 534.46 ± 43.58 | 470.00 ± 39.51 | 552.57 ± 44.80 |
| Fat yield (g) | 29.54 ± 3.05 a | 21.23 ± 2.39 b | 25.10 ± 3.26 | 22.55 ± 2.01 | 19.96 ± 1.82b | 22.41 ± 2.06 |
| Protein yield (g) | 22.28 ± 2.27 a | 18.83 ± 1.78 | 18.91 ± 2.42 | 18.80 ± 1.49 | 16.46 ± 1.35b | 17.42 ± 1.53 |
| Lactose yield (g) | 26.68 ± 3.00 | 22.93 ± 2.35 | 22.26 ± 3.20 | 23.35 ± 1.97 | 20.71 ± 1.79 | 24.49 ± 2.03 |
| Solids-not-fat (g) | 53.08 ± 5.64 | 45.21 ± 4.42 | 44.49 ± 6.03 | 45.85 ± 3.71 | 40.26 ± 3.36 | 45.46 ± 3.81 |
| Casein (g) | 16.78 ± 1.67 | 14.10 ± 1.33 | 14.25 ± 1.81 | 14.02 ± 1.11 | 12.30 ± 1.01 | 12.77 ± 1.14 |
| Fat (%) | 4.90 ± 0.26 a | 4.06 ± 0.20 b | 4.97 ± 0.26 a | 4.33 ± 0.17 | 4.32 ± 0.15 | 4.08 ± 0.17 b |
| Protein (%) | 3.67 ± 0.09 A | 3.65 ± 0.07 A | 3.78 ± 0.09 A | 3.56 ± 0.06A | 3.50 ± 0.05 a | 3.22 ± 0.06 B,b |
| Lactose (%) | 4.38 ± 0.06 | 4.35 ± 0.05 | 4.40 ± 0.06 | 4.33 ± 0.04 | 4.44 ± 0.03 | 4.43 ± 0.04 |
| Solids-not-fat (%) | 8.73 ± 0.10 a | 8.66 ± 0.08 a | 8.85 ± 0.10 A | 8.60 ± 0.07 a | 8.61 ± 0.06 a | 8.31 ± 0.07 B,b |
| Casein (%) | 2.76 ± 0.08 A | 2.72 ± 0.06 A | 2.86 ± 0.08 A | 2.66 ± 0.05 A | 2.62 ± 0.05 A | 2.38 ± 0.05 B |
| Urea (mg/100 mL) | 48.94 ± 2.14 | 49.36 ± 1.68 | 47.46 ± 2.14 | 50.05 ± 1.40 | 51.63 ± 1.25 | 50.53 ± 1.39 |
| Somatic cell count (×103) | 1909.13 ± 689.73 | 1616.04 ± 541.07 | 1602.31 ± 689.73 | 2006.89 ± 466.35 | 1610.36 ± 402.43 | 1598.94 ± 447.55 |
| Cluster | |||
|---|---|---|---|
| Parameters | Strong n = 55 | Intermediate n = 78 | Weak n = 35 |
| Milk yield (g/milking) | 542.68 ± 35.37 | 499.09 ± 29.23 | 552.57 ± 44.74 |
| Fat yield (g) | 24.57 ± 1.64 | 21.13 ± 1.36 | 22.41 ± 2.08 |
| Protein yield (g) | 19.83 ± 1.21 | 17.51 ± 1.00 | 17.42 ± 1.53 |
| Lactose yield (g) | 23.84 ± 1.60 | 21.90 ± 1.32 | 24.49 ± 2.02 |
| Solids-not-fat (g) | 47.28 ± 3.01 | 42.78 ± 2.49 | 45.46 ± 3.81 |
| Casein (g) | 14.90 ± 0.90 | 13.08 ± 0.75 | 12.77 ± 1.14 |
| Fat (%) | 4.54 ± 0.14 A | 4.32 ± 0.12 | 4.08 ± 0.17 B |
| Protein (%) | 3.69 ± 0.05 A,a | 3.53 ± 0.07 A,b | 3.22 ± 0.09 B |
| Lactose (%) | 4.37 ± 0.03 | 4.39 ± 0.03 | 4.43 ± 0.04 |
| Solids-not-fat (%) | 8.73 ± 0.05 A | 8.61 ± 0.05 A | 8.31 ± 0.07 B |
| Casein (%) | 2.77 ± 0.04 A,a | 2.64 ± 0.03 A,b | 2.38 ± 0.05 B |
| Urea (mg/100 mL) | 48.72 ± 1.12 | 50.94 ± 0.93 | 50.53 ± 1.38 |
| Somatic cell count (×103) | 1693.10 ± 359.72 | 1779.61 ± 302.53 | 1598.94 ± 444.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cosenza, G.; Albarella, S.; D’Anza, E.; Iannuzzi, A.; Selvaggi, M.; Pugliano, M.; Galli, T.; Saralli, G.; Ciotola, F.; Peretti, V. A New AS-PCR Method to Detect CSN201 Allele, Genotyping at Ca-Sensitive Caseins Loci and Milk Traits Association Studies in Autochthonous Lazio Goats. Animals 2023, 13, 239. https://doi.org/10.3390/ani13020239
Cosenza G, Albarella S, D’Anza E, Iannuzzi A, Selvaggi M, Pugliano M, Galli T, Saralli G, Ciotola F, Peretti V. A New AS-PCR Method to Detect CSN201 Allele, Genotyping at Ca-Sensitive Caseins Loci and Milk Traits Association Studies in Autochthonous Lazio Goats. Animals. 2023; 13(2):239. https://doi.org/10.3390/ani13020239
Chicago/Turabian StyleCosenza, Gianfranco, Sara Albarella, Emanuele D’Anza, Alessandra Iannuzzi, Maria Selvaggi, Mariagiulia Pugliano, Tiziana Galli, Giorgio Saralli, Francesca Ciotola, and Vincenzo Peretti. 2023. "A New AS-PCR Method to Detect CSN201 Allele, Genotyping at Ca-Sensitive Caseins Loci and Milk Traits Association Studies in Autochthonous Lazio Goats" Animals 13, no. 2: 239. https://doi.org/10.3390/ani13020239