
Feed Clusters According to In Situ and In Vitro Ruminal Crude Protein Degradation
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Feedstuffs and Treatments
2.2. Origin of In Situ Data
2.3. In Vitro Incubations
2.4. Effective Protein Degradation
2.5. Chemical Analyses
2.6. Statistical Analysis
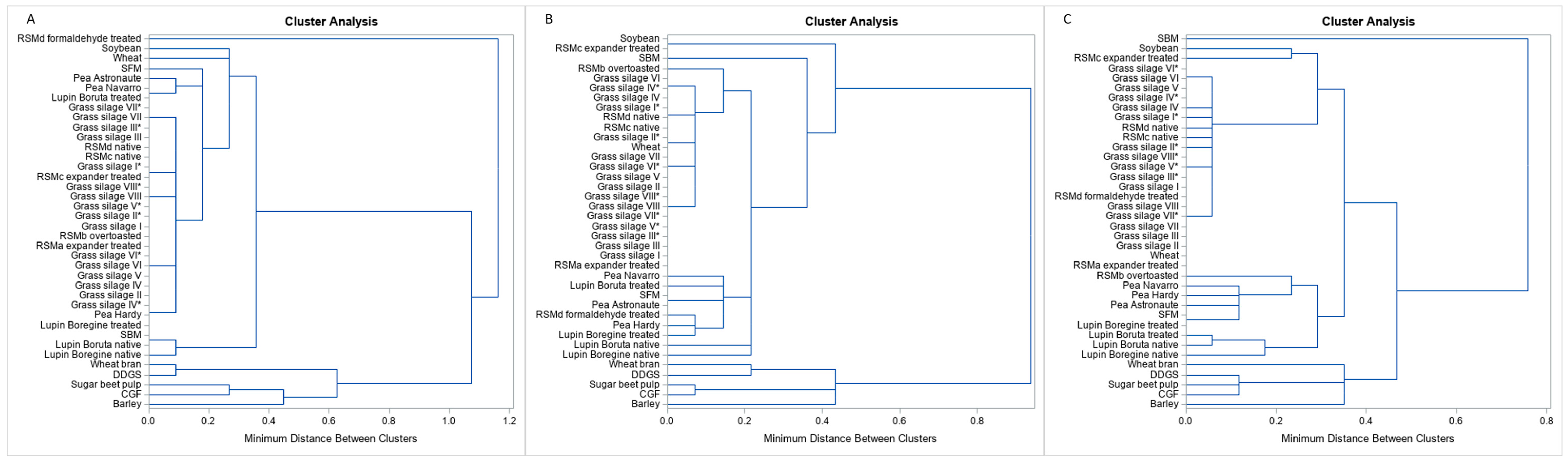
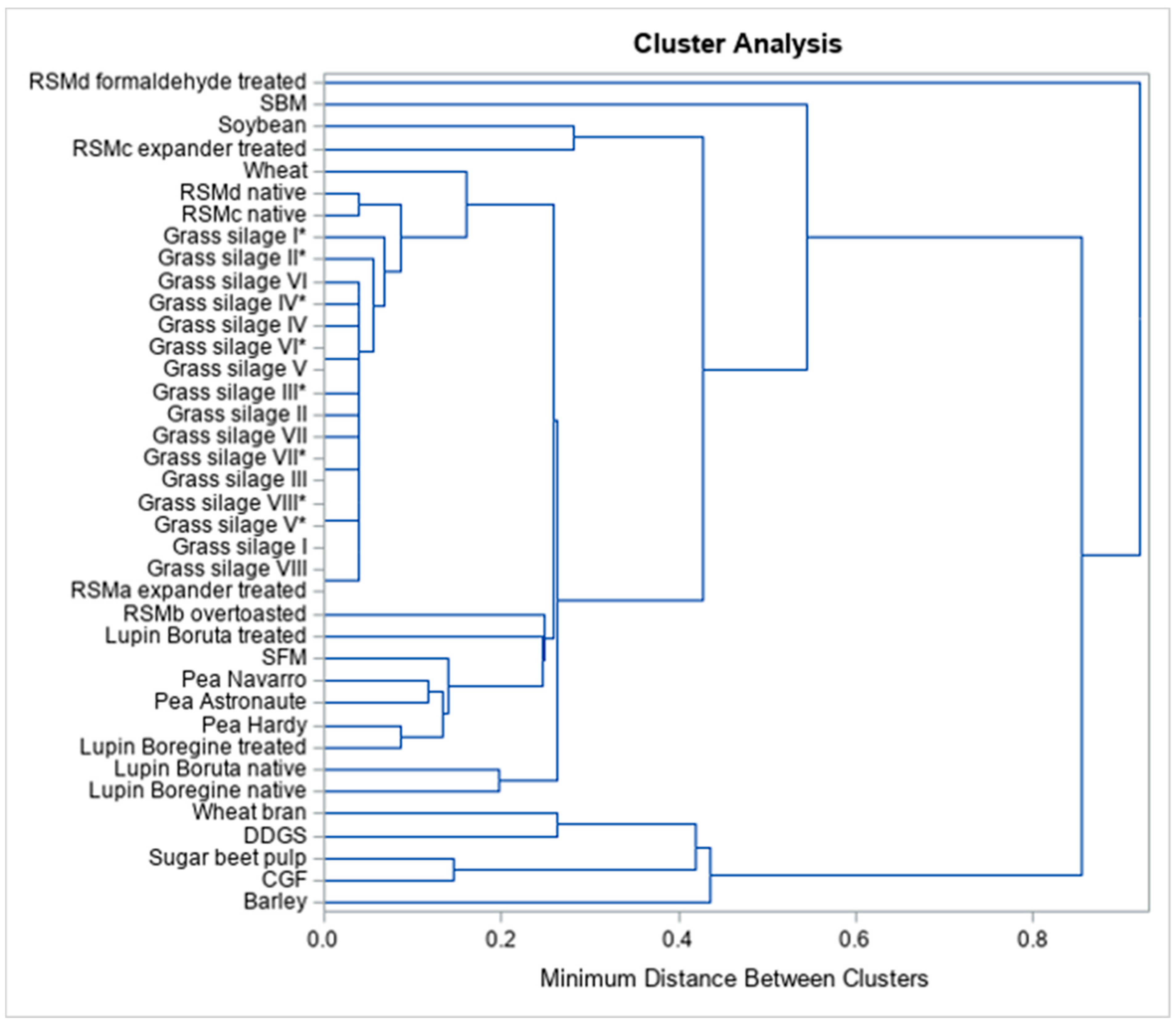
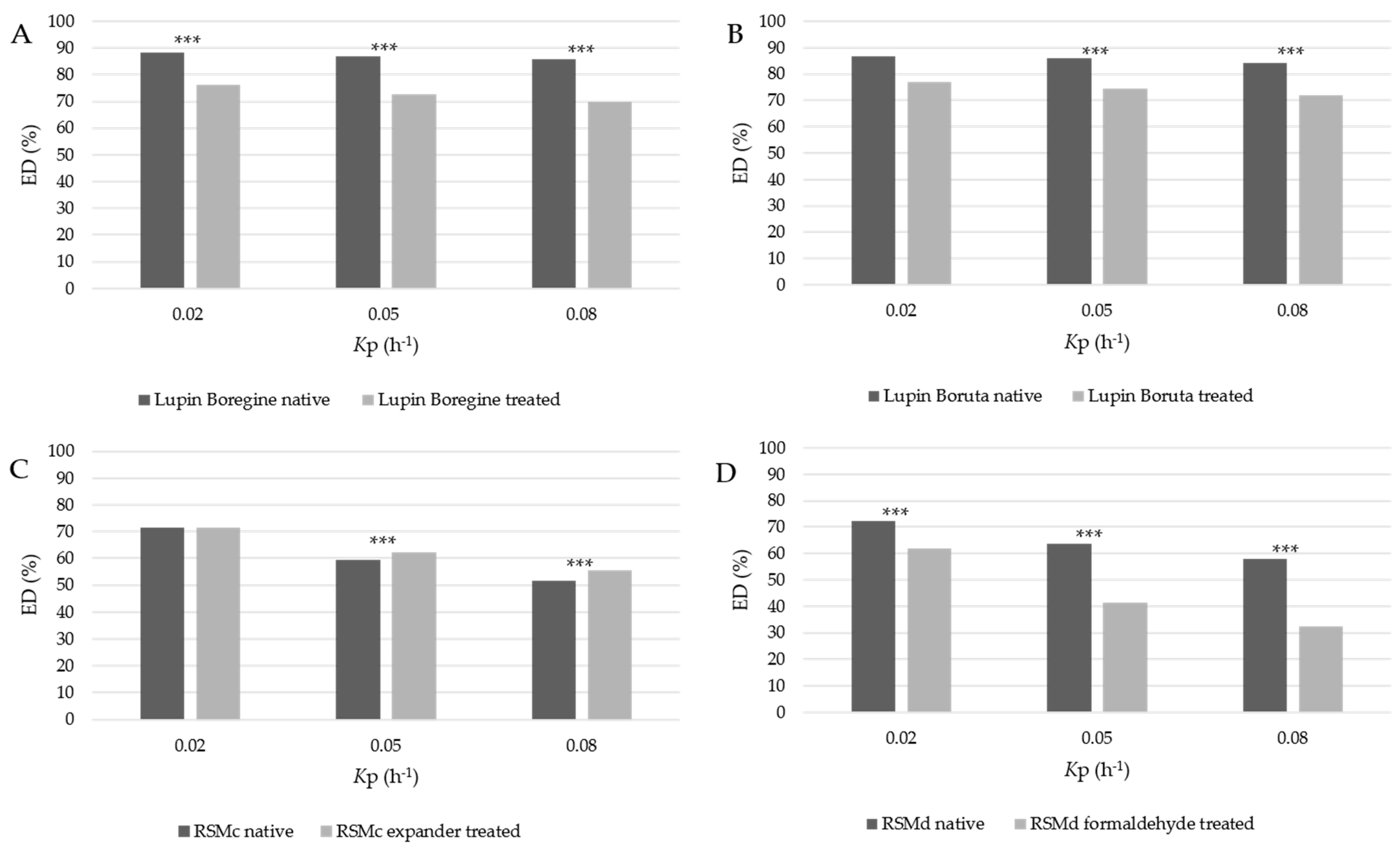
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poos-Floyd, M.; Klopfenstein, T.; Britton, R.A. Evaluation of Laboratory Techniques for Predicting Ruminal Protein Degradation. J. Dairy Sci. 1985, 68, 829–839. [Google Scholar] [CrossRef]
- Michalet-Doreau, B.; Ould-Bah, M.Y. In Vitro and in Sacco Methods for the Estimation of Dietary Nitrogen Degradability in the Rumen: A Review. Anim. Feed. Sci. Technol. 1992, 40, 57–86. [Google Scholar] [CrossRef]
- Stern, M.D.; Bach, A.; Calsamiglia, S. Alternative Techniques for Measuring Nutrient Digestion in Ruminants. J. Anim. Sci. 1997, 75, 2256. [Google Scholar] [CrossRef]
- Hvelplund, T.; Weisbjerg, M. Forage evaluation in ruminant nutrition. In In Situ Techniques for the Estimation of Protein Degradability and Postrumen Availability; CABI Publishing: Wallingford, UK, 2000; pp. 233–258. [Google Scholar]
- GfE [Gesellschaft für Ernährungsphysiologie] Recommended Protocol for the Determination of Nutrient Disappearance in Situ for Estimation of Ruminal Degradation. Proc. Soc. Nutr. Physiol. 2022, 31, 177–189.
- Parand, E.; Spek, J.W. Development of Equations to Estimate Microbial Nitrogen Contamination in Rumen Incubation Residues Using 15N Data and Chemical Composition of Feedstuffs. Anim. Feed Sci. Technol. 2021, 273, 114798. [Google Scholar] [CrossRef]
- Edmunds, B.; Südekum, K.-H.; Spiekers, H.; Schwarz, F.J. Estimating Ruminal Crude Protein Degradation of Forages Using in Situ and In Vitro Techniques. Anim. Feed Sci. Technol. 2012, 175, 95–105. [Google Scholar] [CrossRef]
- Krishnamoorthy, U.; Sniffen, C.J.; Stern, M.D.; Van Soest, P.J. Evaluation of a Mathematical Model of Rumen Digestion and an in Vitro Simulation of Rumen Proteolysis to Estimate the Rumen-Undegraded Nitrogen Content of Feedstuffs. Br. J. Nutr. 1983, 50, 555–568. [Google Scholar] [CrossRef] [Green Version]
- Nomoto, M.; Narahashi, Y.; Murakami, M. A Proteolytic Enzyme of Streptomyces Griseus VI. Hydrolysis of Protein by Streptomyces Griseus. J. Biochem. 1960, 48, 593–602. [Google Scholar] [CrossRef]
- Licitra, G.; Lauria, F.; Carpino, S.; Schadt, I.; Sniffen, C.J.; Van Soest, P.J. Improvement of the Streptomyces Griseus Method for Degradable Protein in Ruminant Feeds. Anim. Feed Sci. Technol. 1998, 72, 1–10. [Google Scholar] [CrossRef]
- Spungin, A.; Blumberg, S. Streptomyces Griseus Aminopeptidase Is a Calcium-Activated Zinc Metalloprotein. Purification and Properties of the Enzyme. Eur. J. Biochem. 1989, 183, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Licitra, G.; Van Soest, P.J.; Schadt, I.; Carpino, S.; Sniffen, C.J. Influence of the Concentration of the Protease from Streptomyces Griseus Relative to Ruminal Protein Degradability. Anim. Feed Sci. Technol. 1999, 77, 99–113. [Google Scholar] [CrossRef]
- Aufrère, J.; Graviou, D.; Demarquilly, C.; Vérité, R.; Michalet-Doreau, B.; Chapoutot, P. Predicting in Situ Degradability of Feed Proteins in the Rumen by Two Laboratory Methods (Solubility and Enzymatic Degradation). Anim. Feed Sci. Technol. 1991, 33, 97–116. [Google Scholar] [CrossRef]
- Cone, J.W.; van Gelder, A.H.; Steg, A.; Van Vuuren, A.M. Prediction of in Situ Rumen Escape Protein from in Vitro Incubation with Protease from Streptomyces Griseus. J. Sci. Food Agric. 1996, 72, 120–126. [Google Scholar] [CrossRef]
- Coblentz, W.K.; Abdelgadir, I.E.O.; Cochran, R.C.; Fritz, J.O.; Fick, W.H.; Olson, K.C.; Turner, J.E. Degradability of Forage Proteins by In Situ and In Vitro Enzymatic Methods. J. Dairy Sci. 1999, 82, 343–354. [Google Scholar] [CrossRef]
- Mathis, C.P.; Cochran, R.C.; Vanzant, E.S.; Abdelgadir, I.E.O.; Heldt, J.S.; Olson, K.C.; Johnson, D.E.; Caton, J.; Faulkner, D.; Horn, G.; et al. A Collaborative Study Comparing an in Situ Protocol with Single Time-Point Enzyme Assays for Estimating Ruminal Protein Degradability of Different Forages. Anim. Feed Sci. Technol. 2001, 93, 31–42. [Google Scholar] [CrossRef]
- Abdelgadir, I.E.; Cochran, R.C.; Titgemeyer, E.C.; Vanzant, E.S. In Vitro Determination of Ruminal Protein Degradability of Alfalfa and Prairie Hay via a Commercial Protease in the Presence or Absence of Cellulase or Driselase. J. Anim. Sci. 1997, 75, 2215. [Google Scholar] [CrossRef] [Green Version]
- Assoumani, M.B.; Vedeau, F.; Jacquot, L.; Sniffen, C.J. Refinement of an Enzymatic Method for Estimating the Theoretical Degradability of Proteins in Feedstuffs for Ruminants. Anim. Feed Sci. Technol. 1992, 39, 357–368. [Google Scholar] [CrossRef]
- Pedersen, M.B.; Dalsgaard, S.; Arent, S.; Lorentsen, R.; Knudsen, K.E.B.; Yu, S.; Lærke, H.N. Xylanase and Protease Increase Solubilization of Non-Starch Polysaccharides and Nutrient Release of Corn- and Wheat Distillers Dried Grains with Solubles. Biochem. Eng. J. 2015, 98, 99–106. [Google Scholar] [CrossRef]
- Okon, P.; Bachmann, M.; Titze, N.; Rodehutscord, M.; Zeyner, A. Determination of Ruminal Protein Degradation of Concentrate Feeds in Situ and in Vitro. Proc. Soc. Nutr. Physiol. 2021, 30, 132. [Google Scholar]
- Mosenthin, R.; Messerschmidt, U.; Sauer, N.; Carré, P.; Quinsac, A.; Schöne, F. Effect of the Desolventizing/Toasting Process on Chemical Composition and Protein Quality of Rapeseed Meal. J. Anim. Sci. Biotechnol. 2016, 7, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupp, C.; Westreicher-Kristen, E.; Susenbeth, A. Effect of Wilting and Lactic Acid Bacteria Inoculant on in Situ and in Vitro Determined Protein Value of Grass Silages. Anim. Feed Sci. Technol. 2021, 282, 115115. [Google Scholar] [CrossRef]
- Licitra, G.; Hernandez, T.M.; Van Soest, P.J. Standardization of Procedures for Nitrogen Fractionation of Ruminant Feeds. Anim. Feed Sci. Technol. 1996, 57, 347–358. [Google Scholar] [CrossRef]
- Higgs, R.J.; Chase, L.E.; Ross, D.A.; Van Amburgh, M.E. Updating the Cornell Net Carbohydrate and Protein System Feed Library and Analyzing Model Sensitivity to Feed Inputs. J. Dairy Sci. 2015, 98, 6340–6360. [Google Scholar] [CrossRef] [Green Version]
- Grubješić, G.; Titze, N.; Krieg, J.; Rodehutscord, M. Determination of in Situ Ruminal Crude Protein and Starch Degradation Values of Compound Feeds from Single Feeds. Arch. Anim. Nutr. 2019, 73, 414–429. [Google Scholar] [CrossRef]
- Titze, N.; Krieg, J.; Steingass, H.; Rodehutscord, M. Variation of Lupin Protein Degradation in Ruminants Studied In Situ and Using Chemical Protein Fractions. Animal 2019, 13, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Titze, N.; Krieg, J.; Steingass, H.; Rodehutscord, M. In Situ Crude Protein and Starch Degradation and In Vitro Evaluation of Pea Grains for Ruminants. Arch. Anim. Nutr. 2021, 75, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Haese, E.; Titze, N.; Rodehutscord, M. IN SITU Ruminal Disappearance of Crude Protein and Phytate from Differently Processed Rapeseed Meals in Dairy Cows. J. Sci. Food Agric. 2022, 102, 2805–2812. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.; Kuhnitzsch, C.; Okon, P.; Martens, S.D.; Greef, J.M.; Steinhöfel, O.; Zeyner, A. Ruminal In Vitro Protein Degradation and Apparent Digestibility of Energy and Nutrients in Sheep Fed Native or Ensiled + Toasted Pea (Pisum sativum) Grains. Animals 2019, 9, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, I. A Revised Model for the Estimation of Protein Degradability in the Rumen. J. Agric. Sci. 1981, 96, 251–252. [Google Scholar] [CrossRef]
- Steingass, H.; Südekum, K.-H. Proteinbewertung Beim Wiederkäuer—Grundlagen, Analytische Entwicklungen Und Perspektiven. Arch. Tierernähr. 2013, 41, 51–73. [Google Scholar]
- Wulf, M.; Südekum, K.-H. Effects of Chemically Treated Soybeans and Expeller Rapeseed Meal on in Vivo and in Situ Crude Fat and Crude Protein Disappearance from the Rumen. Anim. Feed Sci. Technol. 2005, 118, 215–227. [Google Scholar] [CrossRef]
- VDLUFA Die Chemische Unterschung von Futtermitteln; Methodenbuch 3; VDLUFA-Verlag: Darmstadt, Germany, 2012.
- Seifried, N.; Steingaß, H.; Schipprack, W.; Rodehutscord, M. Variation in Ruminal in Situ Degradation of Crude Protein and Starch from Maize Grains Compared to in Vitro Gas Production Kinetics and Physical and Chemical Characteristics. Arch. Anim. Nutr. 2016, 70, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Deutsche Landwirtschafts-Gesellschaft. DLG Futterwerttabellen Wiederkäuer; DLG-Verlag: Frankfurt am Main, Germany, 1997; Volume 7. [Google Scholar]
- Cone, J.W.; Kamman, A.A.; van Gelder, A.H.; Hindle, V.A. Rumen Escape Protein in Concentrate Ingredients Determined with the Nylon Bag and Enzymatic Techniques. Anim. Feed Sci. Technol. 2002, 97, 247–254. [Google Scholar] [CrossRef]
- Cone, J.W.; van Gelder, A.H.; Mathijssen-Kamman, A.A.; Hindle, V.A. Rumen Escape Protein in Grass and Grass Silage Determined with a Nylon Bag and an Enzymatic Technique. Anim. Feed Sci. Technol. 2004, 111, 1–9. [Google Scholar] [CrossRef]
- Hristov, A.N.; Bannink, A.; Crompton, L.A.; Huhtanen, P.; Kreuzer, M.; McGee, M.; Nozière, P.; Reynolds, C.K.; Bayat, A.R.; Yáñez-Ruiz, D.R.; et al. Invited Review: Nitrogen in Ruminant Nutrition: A Review of Measurement Techniques. J. Dairy Sci. 2019, 102, 5811–5852. [Google Scholar] [CrossRef] [Green Version]
- Krawielitzki, K.; Schmidt, T.; Voigt, J.; Kowalczyk, J.; Gabel, M. Dynamics of Microbial Contamination of Proteinduring Ruminal In Situ Incubation of Feedstuffs. J. Anim. Feed Sci. 2006, 15, 313–328. [Google Scholar] [CrossRef]
- Mackie, R.I.; McSweeney, C.S.; Aminov, R.I. Rumen. In Encyclopedia of Life Sciences; John Wiley & Sons Ltd., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2013; p. a0000404. ISBN 978-0-470-01590-2. [Google Scholar]
- Deusch, S.; Camarinha-Silva, A.; Conrad, J.; Beifuss, U.; Rodehutscord, M.; Seifert, J. A Structural and Functional Elucidation of the Rumen Microbiome Influenced by Various Diets and Microenvironments. Front. Microbiol. 2017, 8, 1605. [Google Scholar] [CrossRef] [Green Version]
- Ozbayram, E.; Ince, O.; Ince, B.; Harms, H.; Kleinsteuber, S. Comparison of Rumen and Manure Microbiomes and Implications for the Inoculation of Anaerobic Digesters. Microorganisms 2018, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhang, M.; Zhang, R.; Zhu, W.; Mao, S. Comparative Studies of the Composition of Bacterial Microbiota Associated with the Ruminal Content, Ruminal Epithelium and in the Faeces of Lactating Dairy Cows. Microb. Biotechnol. 2016, 9, 257–268. [Google Scholar] [CrossRef]
- Nyonyo, T.; Shinkai, T.; Mitsumori, M. Improved Culturability of Cellulolytic Rumen Bacteria and Phylogenetic Diversity of Culturable Cellulolytic and Xylanolytic Bacteria Newly Isolated from the Bovine Rumen. FEMS Microbiol. Ecol. 2014, 88, 528–537. [Google Scholar] [CrossRef] [Green Version]
- Bach, A.; Calsamiglia, S.; Stern, M.D. Nitrogen Metabolism in the Rumen. J. Dairy Sci. 2005, 88, E9–E21. [Google Scholar] [CrossRef] [Green Version]
- Debroas, D.; Blanchart, G. Interactions between Proteolytic and Cellulolytic Rumen Bacteria during Hydrolysis of Plant Cell Wall Protein. Reprod. Nutr. Dev. 1993, 33, 283–288. [Google Scholar] [CrossRef]
- Hedqvist, H.; Udén, P. Measurement of Soluble Protein Degradation in the Rumen. Anim. Feed Sci. Technol. 2006, 126, 1–21. [Google Scholar] [CrossRef]
- Nikokyris, P.N.; Kandylis, K. Feed Protein Fractions in Various Solvents of Ruminant Feedstuffs. J. Sci. Food Agric. 1997, 75, 198–204. [Google Scholar] [CrossRef]
- Romagnolo, D.; Polan, C.E.; Barbeau, W.E. Electrophoretic Analysis of Ruminal Degradability of Corn Proteins. J. Dairy Sci. 1994, 77, 1093–1099. [Google Scholar] [CrossRef]
- Jayanegara, A.; Dewi, S.P.; Laylli, N.; Laconi, E.B.; Nahrowi, N.; Ridla, M. Determination of Cell Wall Protein from Selected Feedstuffs and Its Relationship with Ruminal Protein Digestibility In Vitro. Media Peternak. 2016, 39, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Alzuwaid, N.T.; Sissons, M.; Laddomada, B.; Fellows, C.M. Nutritional and Functional Properties of Durum Wheat Bran Protein Concentrate. Cereal Chem. 2020, 97, 304–315. [Google Scholar] [CrossRef]
- DePeters, E.J.; Fadel, J.G.; Arosemena, A. Digestion Kinetics of Neutral Detergent Fiber and Chemical Composition within Some Selected By-Product Feedstuffs. Anim. Feed Sci. Technol. 1997, 67, 127–140. [Google Scholar] [CrossRef]
- Papadopoulos, M.C. Effect of Processing on High-Protein Feedstuffs: A Review. Biol. Wastes 1989, 29, 123–138. [Google Scholar] [CrossRef]
- Mustafa, A.F.; McKinnon, J.J.; Christensen, D.A. Protection of Canola (Low Glucosinolate Rapeseed) Meal and Seed Protein from Ruminal Degradation—Review—Asian-Australas. J. Anim. Sci. 2000, 13, 535–542. [Google Scholar] [CrossRef]
- Jha, R.; Woyengo, T.A.; Li, J.; Bedford, M.R.; Vasanthan, T.; Zijlstra, R.T. Enzymes Enhance Degradation of the Fiber–Starch–Protein Matrix of Distillers Dried Grains with Solubles as Revealed by a Porcine in Vitro Fermentation Model and Microscopy. J. Anim. Sci. 2015, 93, 1039. [Google Scholar] [CrossRef] [PubMed]
- McAllister, T.A.; Phillippe, R.C.; Rode, L.M.; Cheng, K.-J. Effect of the Protein Matrix on the Digestion of Cereal Grains by Ruminal Microorganisms. J. Anim. Sci. 1993, 71, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Doiron, K.; Yu, P.; McKinnon, J.J.; Christensen, D.A. Heat-Induced Protein Structure and Subfractions in Relation to Protein Degradation Kinetics and Intestinal Availability in Dairy Cattle. J. Dairy Sci. 2009, 92, 3319–3330. [Google Scholar] [CrossRef] [PubMed]
- Goelema, J.O.; Spreeuwenberg, M.A.M.; Hof, G.; van der Poel, A.F.B.; Tamminga, S. Effect of Pressure Toasting on the Rumen Degradability and Intestinal Digestibility of Whole and Broken Peas, Lupins and Faba Beans and a Mixture of These Feedstuffs. Anim. Feed Sci. Technol. 1998, 76, 35–50. [Google Scholar] [CrossRef]
- Goelema, J.O.; Smits, A.; Vaessen, L.M.; Wemmers, A. Effects of Pressure Toasting, Expander Treatment and Pelleting on In Vitro and In Situ Parameters of Protein and Starch in a Mixture of Broken Peas, Lupins and Faba Beans. Anim. Feed Sci. Technol. 1999, 78, 109–126. [Google Scholar] [CrossRef]
- Ljøkjel, K.; Harstad, O.M.; Prestløkken, E.; Skrede, A. In Situ Digestibility of Protein in Barley Grain (Hordeum vulgare) and Peas (Pisum sativum L.) in Dairy Cows: Influence of Heat Treatment and Glucose Addition. Anim. Feed Sci. Technol. 2003, 107, 87–104. [Google Scholar] [CrossRef]
- Hofmann, T.; Engling, A.C.; Martens, S.; Steinhöfel, O.; Henle, T. Quantification of Maillard Reaction Products in Animal Feed. Eur. Food Res. Technol. 2020, 246, 253–256. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Zhong, F.; Douglas Goff, H.; Li, Y. Study on Starch-Protein Interactions and Their Effects on Physicochemical and Digestible Properties of the Blends. Food Chem. 2019, 280, 51–58. [Google Scholar] [CrossRef]
- Wroblewitz, S.; Hüther, L.; Manderscheid, R.; Weigel, H.-J.; Wätzig, H.; Dänicke, S. Effect of Rising Atmospheric Carbon Dioxide Concentration on the Protein Composition of Cereal Grain. J. Agric. Food Chem. 2014, 62, 6616–6625. [Google Scholar] [CrossRef]
- Bochnia, M.; Walther, S.; Schenkel, H.; Romanowski, K.; Zeyner, A. Comparison of Scanning Electron Microscopic Examination of Oats, Barley and Maize Grains with the Analyzed Degree of Starch Breakdown and Glycaemic Responses in Horses. Int. J. Sci. Res. Sci. Technol. 2015, 1, 81–84. [Google Scholar]
- Bochnia, M.; Walther, S.; Schenkel, H.; Romanowski, K.; Zeyner, A. Scanning Electron Microscopic Examination of Different Varieties of Oat Grains in Comparison with the Analyzed Degree of Starch Breakdown and Glycaemic Responses in Horses. Int. J. Sci. Res. Sci. Technol. 2015, 1, 110–113. [Google Scholar]
- Bachmann, M.; Kuhnitzsch, C.; Thierbach, A.; Michel, S.; Bochnia, M.; Greef, J.M.; Martens, S.D.; Steinhöfel, O.; Zeyner, A. Effects of Toasting Temperature and Duration on in Vitro Ruminal Gas Production Kinetics and Post-Ruminal Crude Protein from Field Pea (Pisum sativum) Legume Grain Silages. Livest. Sci. 2020, 233, 103944. [Google Scholar] [CrossRef]
- Ljøkjel, K.; Harstad, O.M.; Prestløkken, E.; Skrede, A. In Situ Digestibility of Starch in Barley Grain (Hordeum vulgare) and Peas (Pisum sativum L.) in Dairy Cows: Influence of Heat Treatment and Glucose Addition. Anim. Feed Sci. Technol. 2003, 107, 105–116. [Google Scholar] [CrossRef]
- Vagadia, B.H.; Vanga, S.K.; Raghavan, V. Inactivation Methods of Soybean Trypsin Inhibitor—A Review. Trends Food Sci. Technol. 2017, 64, 115–125. [Google Scholar] [CrossRef]
- Guillamón, E.; Pedrosa, M.M.; Burbano, C.; Cuadrado, C.; Sánchez, M.D.C.; Muzquiz, M. The Trypsin Inhibitors Present in Seed of Different Grain Legume Species and Cultivar. Food Chem. 2008, 107, 68–74. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feedstuff | Treatment |
|---|---|
| Lupin Boregine native | Native |
| Lupin Boregine treated | Toasted at 115–120 °C for 1 min, conditioned for 30 min in cooling tower followed by cooling to 20 °C. |
| Lupin Boruta native | Native |
| Lupin Boruta treated | Moisture conditioned, short time toasted at 130 °C and drying to 940 g DM/kg. |
| RSMa expander-treated | Expanded (unknown conditions) |
| RSMb over-toasted | Toasted at 107 °C for 60 min under 450 kPa pressure [21] |
| RSMc native | Native |
| RSMc expander-treated | Expanded (unknown conditions) |
| RSMd native | Native |
| RSMd formaldehyde-treated | Formaldehyde-treated (unknown conditions) |
| Feedstuff | DM | CA | CP | TP | SP | AEE | aNDFom | ADFom | Starch |
|---|---|---|---|---|---|---|---|---|---|
| Barley | 894 | 27 | 125 | 114 | 28 | 33 | 162 | 76 | 532 |
| Wheat | 857 | 19 | 140 | 126 | 32 | 31 | 86 | 36 | 544 |
| Corn | 894 | 17 | 83 | 76 | 18 | 43 | 84 | 30 | 705 |
| Wheat bran | 859 | 59 | 186 | 163 | 34 | 55 | 398 | 135 | 158 |
| DDGS | 863 | 64 | 312 | 252 | 21 | 81 | 338 | 198 | 28 |
| CGF | 867 | 84 | 169 | 87 | 55 | 36 | 343 | 95 | 159 |
| Soybean | 901 | 56 | 391 | 373 | 9 | 221 | 109 | 65 | n.a. |
| SBM | 893 | 71 | 504 | 465 | 11 | 26 | 111 | 66 | 18 |
| SFM | 910 | 78 | 318 | 294 | 36 | 30 | 402 | 307 | 17 |
| RSMa et | 790 | 79 | 358 | 340 | 20 | 45 | 321 | 225 | 38 |
| RSMb ot | 924 | 85 | 366 | 352 | 15 | 23 | 476 | 269 | 9 |
| RSMc n | 885 | 86 | 384 | 357 | 15 | 46 | 324 | 210 | n.a. |
| RSMc et | 900 | 86 | 383 | 358 | 15 | 48 | 321 | 207 | n.a. |
| RSMd n | 900 | 78 | 374 | 359 | 24 | 37 | 324 | 233 | n.a. |
| RSMd ft | 911 | 86 | 370 | 359 | 9 | 41 | 339 | 221 | n.a. |
| Faba bean | 897 | 39 | 279 | 233 | 55 | 23 | 173 | 129 | 358 |
| Lupin Boregine n | 917 | 37 | 298 | 289 | 75 | 68 | 264 | 235 | 298 |
| Lupin Boregine t | 929 | 38 | 320 | 300 | 32 | 75 | 252 | 212 | 320 |
| Lupin Boruta n | 900 | 37 | 319 | 311 | 64 | 67 | 249 | 213 | 319 |
| Lupin Boruta t | 925 | 38 | 328 | 311 | 36 | 67 | 265 | 201 | 328 |
| Pea Hardy | 902 | 32 | 219 | 206 | 68 | 19 | 115 | 83 | 451 |
| Pea Astronaute | 901 | 30 | 228 | 215 | 70 | 19 | 118 | 75 | 432 |
| Pea Navarro | 898 | 30 | 248 | 234 | 73 | 20 | 142 | 81 | 392 |
| SBP | 862 | 85 | 94 | 57 | 40 | 19 | 347 | 174 | n.a. |
| GS I | 932 | 113 | 153 | 48 | 68 | 48 | 463 | 292 | n.a. |
| GS I * | 930 | 117 | 159 | 57 | 68 | 49 | 519 | 307 | n.a. |
| GS II | 926 | 102 | 153 | 50 | 66 | 40 | 498 | 293 | n.a. |
| GS II * | 917 | 105 | 156 | 56 | 65 | 46 | 503 | 309 | n.a. |
| GS III | 928 | 105 | 147 | 48 | 66 | 41 | 508 | 307 | n.a. |
| GS III * | 926 | 104 | 148 | 50 | 66 | 44 | 514 | 316 | n.a. |
| GS IV | 934 | 103 | 153 | 48 | 67 | 38 | 529 | 311 | n.a. |
| GS IV * | 937 | 107 | 155 | 50 | 66 | 42 | 520 | 315 | n.a. |
| GS V | 932 | 103 | 150 | 52 | 65 | 38 | 526 | 310 | n.a. |
| GS V * | 932 | 107 | 153 | 53 | 64 | 40 | 510 | 308 | n.a. |
| GS VI | 932 | 110 | 153 | 52 | 66 | 36 | 527 | 311 | n.a. |
| GS VI * | 929 | 110 | 153 | 51 | 64 | 38 | 518 | 312 | n.a. |
| GS VII | 926 | 108 | 146 | 55 | 65 | 34 | 523 | 307 | n.a. |
| GS VII * | 929 | 109 | 152 | 57 | 65 | 36 | 507 | 299 | n.a. |
| GS VIII | 927 | 111 | 151 | 58 | 65 | 33 | 526 | 309 | n.a. |
| GS VIII * | 932 | 110 | 149 | 60 | 65 | 34 | 514 | 301 | n.a. |
| Feedstuff | ED2 | ED5 | ED8 | |||
|---|---|---|---|---|---|---|
| In Situ | In Vitro | In Situ | In Vitro | In Situ | In Vitro | |
| Barley | 91 aA | 43 bB | 87 aA | 42 bB | 83 aA | 40 bB |
| Wheat | 92 aA | 73 bB | 84 aA | 70 bB | 78 aA | 67 bB |
| Wheat bran | 91 aA | 58 bA | 87 aA | 56 bA | 83 aA | 54 bA |
| DDGS | 86 aA | 55 bB | 82 aA | 50 bB | 79 aA | 46 bB |
| Corn gluten feed | 92 aA | 51 bA | 89 aA | 49 bA | 87 aA | 48 bA |
| Soybeans | 92 aA | 70 bB | 84 aA | 65 bB | 79 aA | 61 bB |
| SBM | 84 aA | 80 bB | 68 aA | 75 bB | 56 aA | 71 bB |
| SFM | 89 aA | 81 bB | 81 aA | 77 bB | 74 aA | 75 aA |
| RSMa et | 81 aA | 67 bB | 71 aA | 60 bB | 63 aA | 55 bB |
| RSMb ot | 70 aA | 58 bB | 58 aA | 52 bB | 50 A | 47 B |
| RSMc n | 85 aA | 71 bA | 72 aA | 59 bA | 63 aA | 52 bA |
| RSMc et | 88 A | 72 A | 81 A | 62 A | 76 aA | 55 bA |
| RSMd n | 86 aA | 72 bA | 77 aA | 64 bA | 70 aA | 58 bA |
| RSMd ft | 57 A | 62 A | 45 aA | 41 aA | 38 A | 32 A |
| Lupin Boregine n | 92 aA | 88 bB | 82 aA | 87 bB | 75 aA | 86 bB |
| Lupin Boregine t | 89 aA | 76 bB | 77 aA | 73 bA | 69 aA | 70 aA |
| Lupin Boruta n | 92 aA | 87 bB | 83 aA | 86 aA | 77 aA | 84 bB |
| Lupin Boruta t | 87 aA | 77 bB | 74 aA | 74 aA | 65 aA | 72 bB |
| Pea Hardy | 93 aA | 80 bB | 85 aA | 79 bB | 79 aA | 78 aA |
| Pea Astronaute | 92 aA | 81 bA | 84 aA | 81 aA | 77 aA | 80 aA |
| Pea Navarro | 92 A | 82 A | 83 aA | 81 aA | 76 aA | 80 bA |
| Sugar beet pulp | 89 aA | 47 bA | 77 aA | 42 bA | 69 aA | 39 bA |
| Grass Silage I | 94 aA | 78 aA | 91 aA | 77 bA | 89 aA | 76 bA |
| Grass Silage I * | 94 aA | 77 bA | 91 aA | 75 bA | 88 aA | 74 bA |
| Grass Silage II | 93 aA | 79 bA | 90 aA | 77 bA | 88 aA | 76 bA |
| Grass Silage II * | 93 aA | 77 bA | 89 aA | 75 bA | 86 aA | 74 bA |
| Grass Silage III | 93 aA | 78 bA | 89 aA | 76 bA | 87 aA | 75 bA |
| Grass Silage III * | 93 aA | 78 bA | 89 aA | 76 bA | 87 aA | 74 bA |
| Grass Silage IV | 93 aA | 79 bA | 89 aA | 78 bA | 87 aA | 77 bA |
| Grass Silage IV * | 92 aA | 79 bA | 89 aA | 77 bA | 86 aA | 76 bA |
| Grass Silage V | 92 aA | 78 bA | 88 aA | 76 bA | 86 aA | 75 bA |
| Grass Silage V * | 92 A | 77 A | 88 aA | 75 bA | 86 aA | 74 bA |
| Grass Silage VI | 93 aA | 79 bA | 89 aA | 77 bA | 86 aA | 76 bA |
| Grass Silage VI * | 93 aA | 79 bA | 89 aA | 77 bA | 86 aA | 75 bA |
| Grass Silage VII | 92 aA | 77 bA | 87 aA | 75 bA | 84 aA | 73 bA |
| Grass Silage VII * | 92 aA | 78 bA | 88 aA | 75 bA | 85 aA | 73 bA |
| Grass Silage VIII | 92 aA | 77 bA | 87 aA | 75 bA | 84 A | 73 A |
| Grass Silage VIII * | 92 aA | 77 bA | 87 aA | 74 bA | 83 aA | 72 bA |
| Range of SD | 0.07–16.38 | 0.29–3.68 | 0.31–9.22 | 0.24–3.16 | 0.16–5.8 | 0.17–3.70 |
| Feedstuffs | 0.02 h−1 | 0.05 h−1 | 0.08 h−1 |
|---|---|---|---|
| Barley | −0.52 | −0.52 | −0.51 |
| Wheat | −0.21 | −0.16 | −0.13 |
| Wheat bran | −0.37 | −0.36 | −0.35 |
| DDGS | −0.36 | −0.39 | −0.41 |
| CGF | −0.44 | −0.45 | −0.45 |
| Soybeans | −0.24 | −0.23 | −0.23 |
| SBM | −0.05 | 0.11 | 0.27 |
| SFM | −0.09 | −0.04 | 0.01 |
| RSMa et | −0.17 | −0.15 | −0.13 |
| RSMb ot | −0.17 | −0.11 | −0.05 |
| RSMc n | −0.16 | −0.17 | −0.18 |
| RSMc et | −0.18 | −0.23 | −0.27 |
| RSMd n | −0.16 | −0.17 | −0.17 |
| RSMd ft | 0.09 | −0.08 | −0.14 |
| Lupin Boregine n | −0.04 | 0.06 | 0.14 |
| Lupin Boregine t | −0.14 | −0.06 | 0.01 |
| Lupin Boruta n | −0.05 | 0.03 | 0.10 |
| Lupin Boruta t | −0.11 | 0.00 | 0.11 |
| Pea Hardy | −0.14 | −0.07 | −0.01 |
| Pea Astronaute | −0.12 | −0.04 | 0.03 |
| Pea Navarro | −0.11 | −0.02 | 0.05 |
| Sugar beet pulp | −0.47 | −0.46 | −0.43 |
| GS I | −0.17 | −0.15 | −0.14 |
| GS I * | −0.18 | −0.17 | −0.16 |
| GS II | −0.15 | −0.14 | −0.13 |
| GS II * | −0.17 | −0.16 | −0.15 |
| GS III | −0.16 | −0.15 | −0.13 |
| GS III * | −0.16 | −0.15 | −0.14 |
| GS IV | −0.15 | −0.13 | −0.11 |
| GS IV * | −0.14 | −0.13 | −0.12 |
| GS V | −0.15 | −0.14 | −0.12 |
| GS V * | −0.17 | −0.15 | −0.14 |
| GS VI | −0.15 | −0.13 | −0.12 |
| GS VI * | −0.15 | −0.14 | −0.12 |
| GS VII | −0.16 | −0.14 | −0.13 |
| GS VII * | −0.16 | −0.15 | −0.13 |
| GS VIII | −0.17 | −0.15 | −0.13 |
| GS VIII * | −0.17 | −0.15 | −0.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okon, P.; Bachmann, M.; Wensch-Dorendorf, M.; Titze, N.; Rodehutscord, M.; Rupp, C.; Susenbeth, A.; Greef, J.M.; Zeyner, A. Feed Clusters According to In Situ and In Vitro Ruminal Crude Protein Degradation. Animals 2023, 13, 224. https://doi.org/10.3390/ani13020224
Okon P, Bachmann M, Wensch-Dorendorf M, Titze N, Rodehutscord M, Rupp C, Susenbeth A, Greef JM, Zeyner A. Feed Clusters According to In Situ and In Vitro Ruminal Crude Protein Degradation. Animals. 2023; 13(2):224. https://doi.org/10.3390/ani13020224
Chicago/Turabian StyleOkon, Paul, Martin Bachmann, Monika Wensch-Dorendorf, Natascha Titze, Markus Rodehutscord, Christiane Rupp, Andreas Susenbeth, Jörg Michael Greef, and Annette Zeyner. 2023. "Feed Clusters According to In Situ and In Vitro Ruminal Crude Protein Degradation" Animals 13, no. 2: 224. https://doi.org/10.3390/ani13020224