
Abundance of Non-Native Birds in the City: Spatial Variation and Relationship with Socioeconomics in a South American City


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Selection of Sampling Sites
2.3. Bird Counts
2.4. Data Analysis
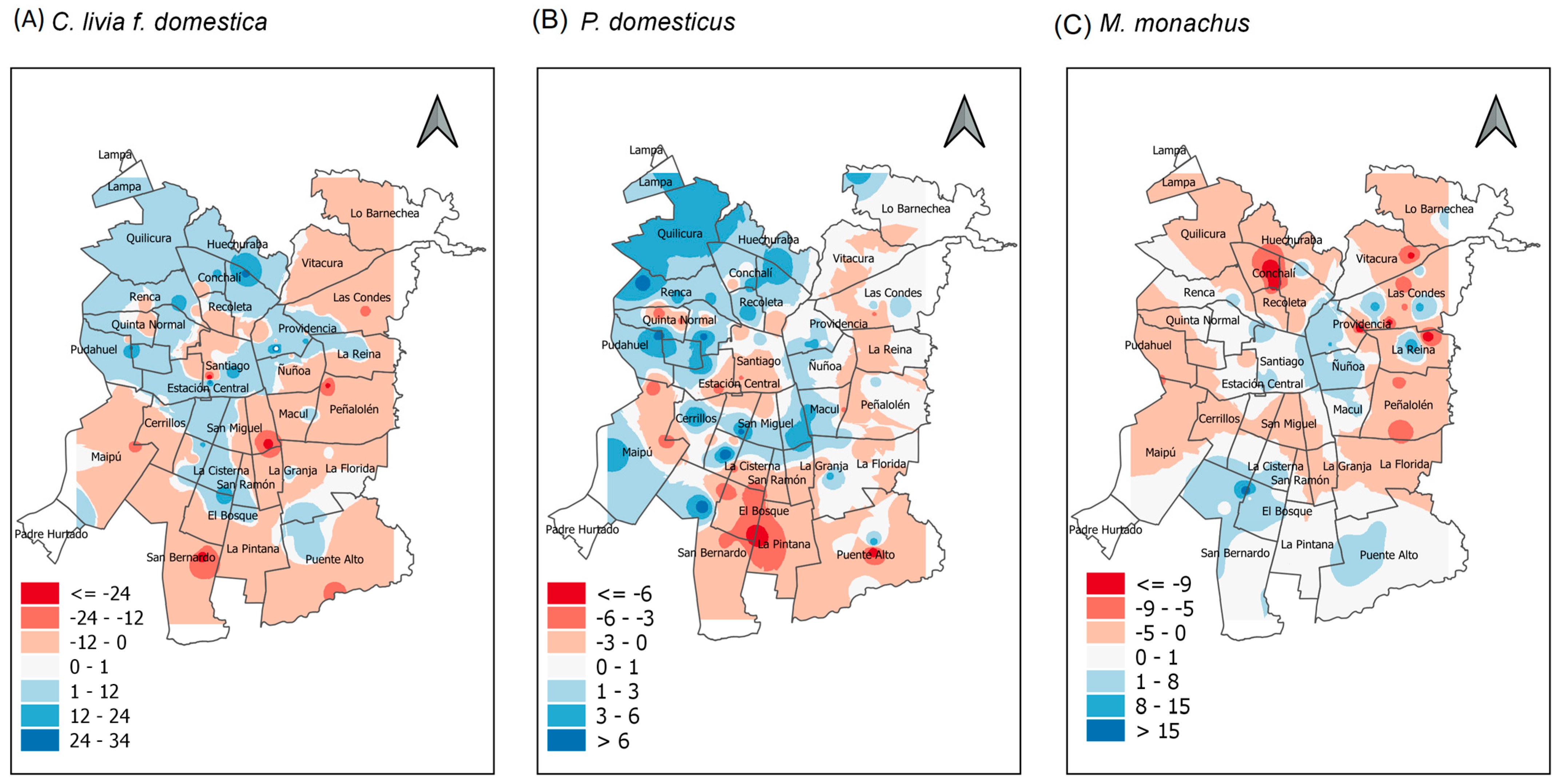
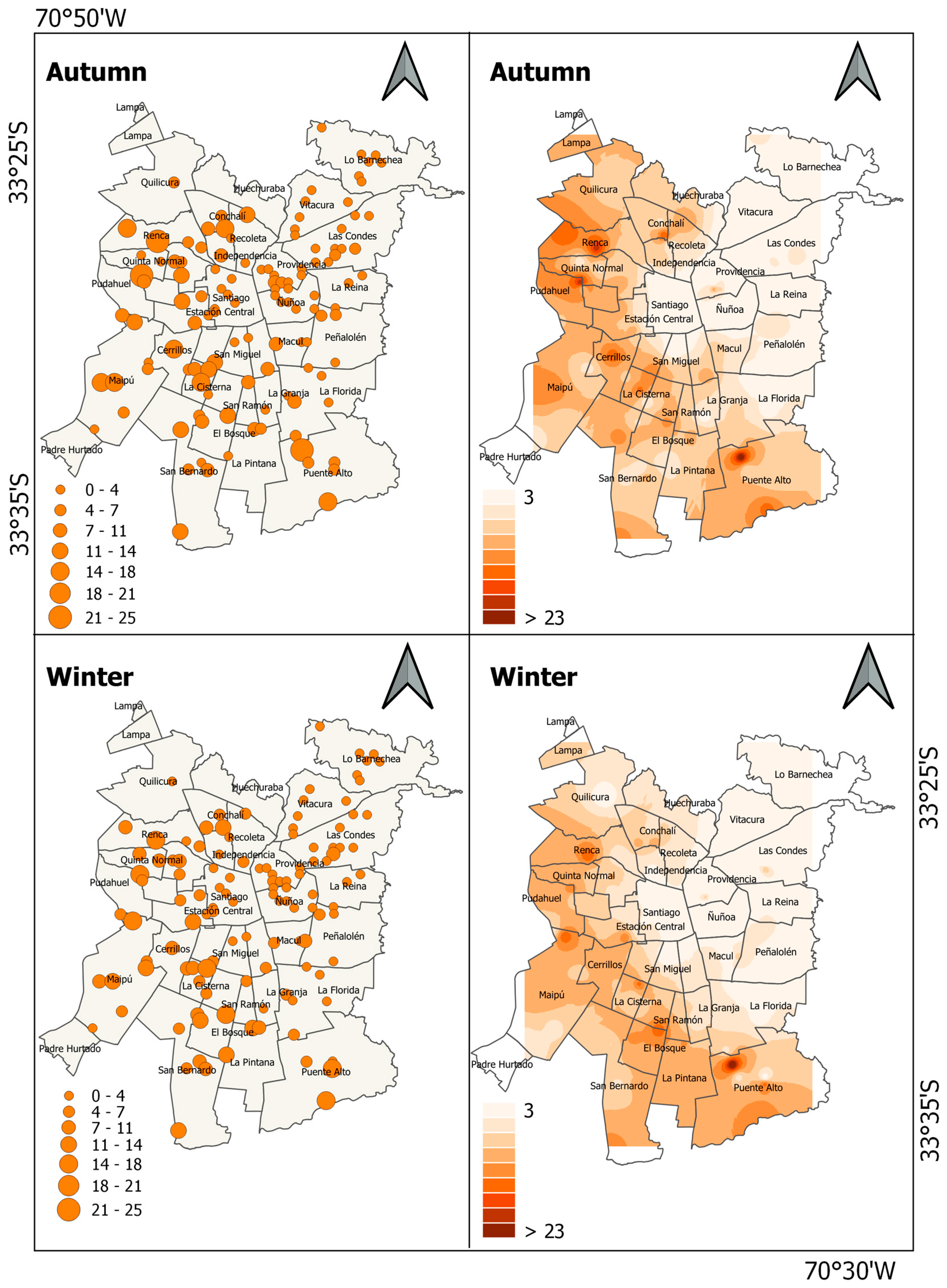
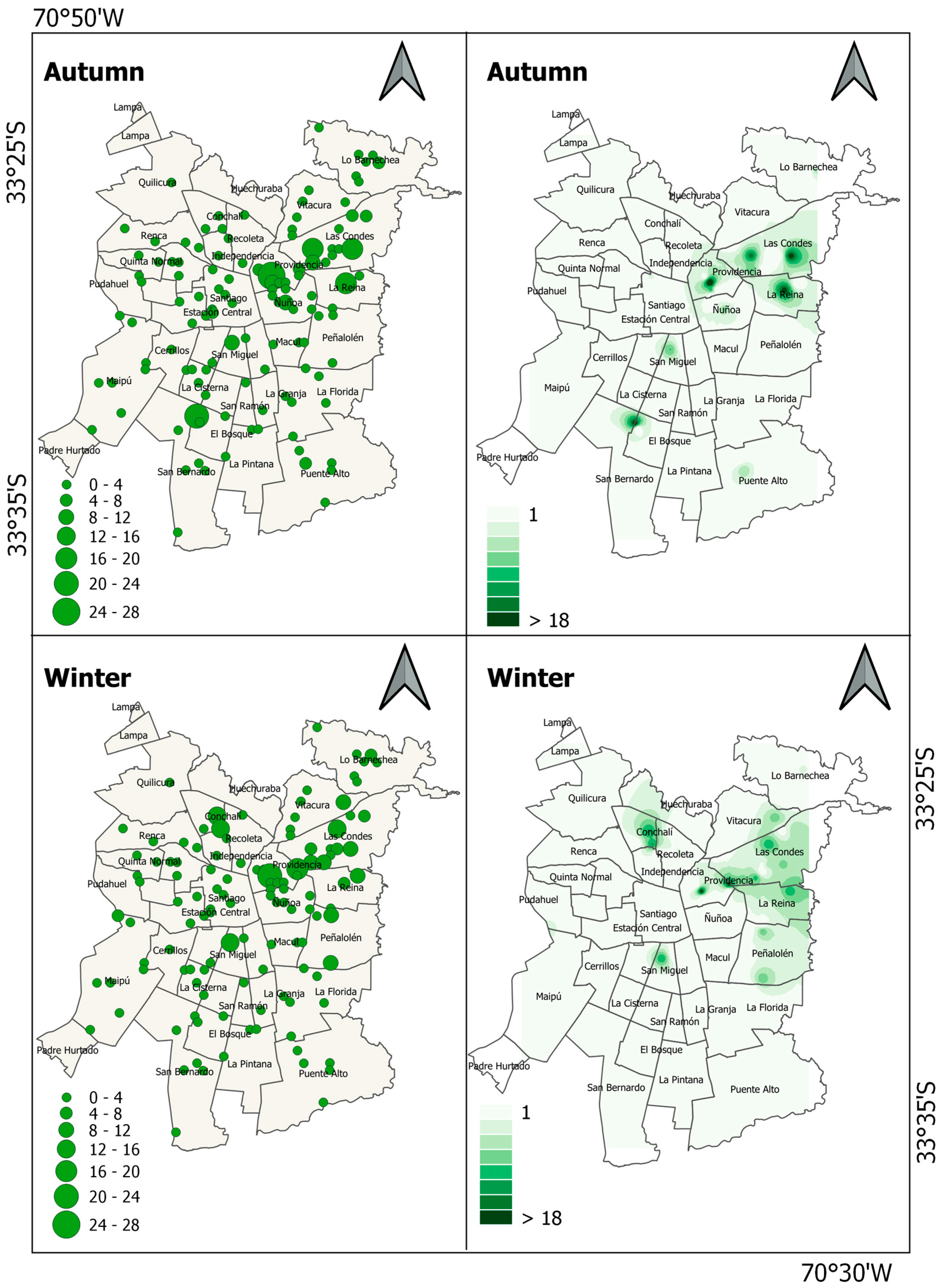
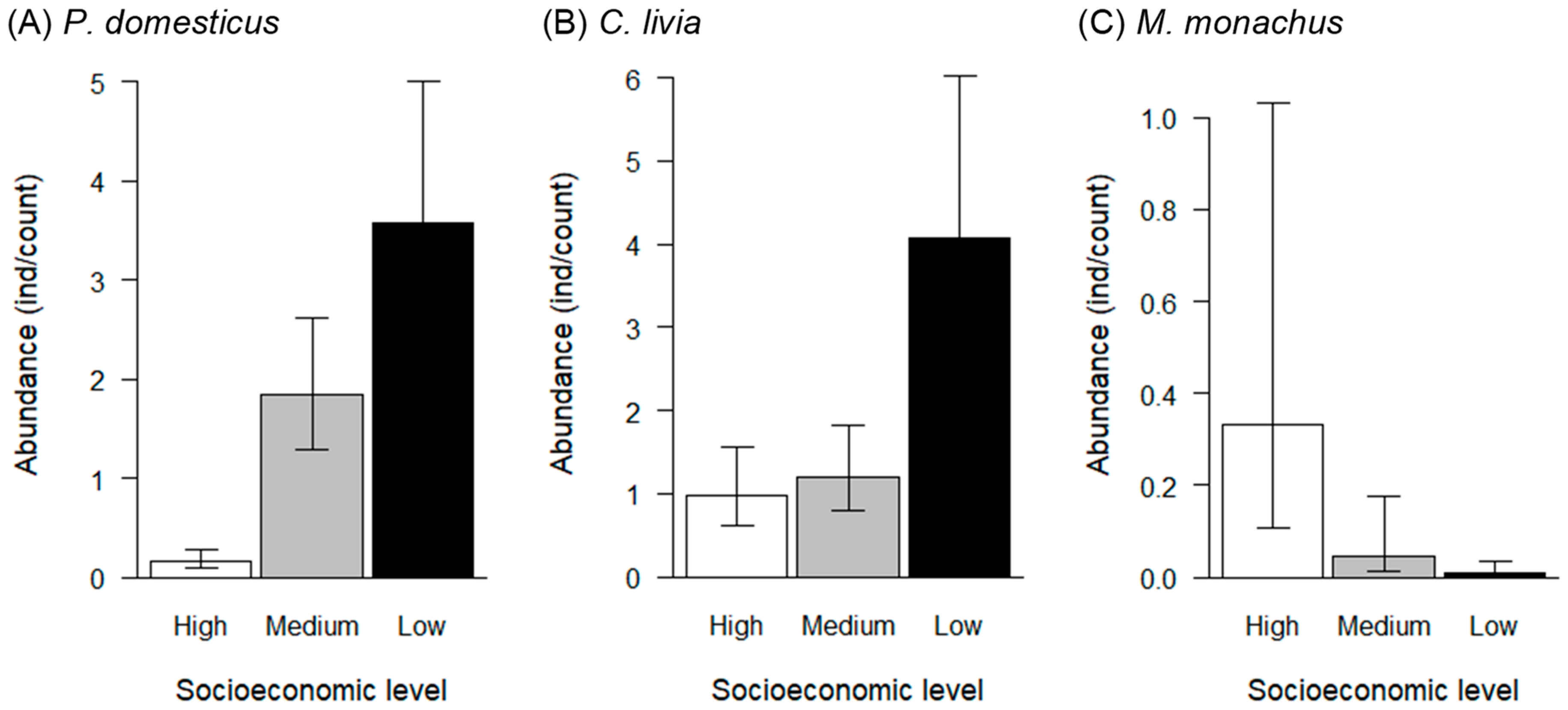
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonacic, C.; Almuna, R.; Ibarra, J.T. Biodiversity Conservation Requires Management of Feral Domestic Animals. Trends Ecol. Evol. 2019, 34, 683–686. [Google Scholar] [CrossRef]
- Manchester, S.J.; Bullock, J.M. The impacts of non-native species on UK biodiversity and the effectiveness of control. J. Appl. Ecol. 2000, 37, 845–864. [Google Scholar] [CrossRef]
- Clout, M.N.; Williams, P.A. Invasive Species Management: A Handbook of Principles and Techniques; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Przybylska, K.; Haidt, A.; Myczko, Ł.; Ekner-Grzyb, A.; Rosin, Z.M.; Kwieciński, Z.; Tryjanowski, P.; Suchodolska, J.; Takacs, V.; Jankowiak, Ł.; et al. Local and Landscape-Level Factors Affecting the Density and Distribution of the Feral Pigeon Columba livia var. domestica in an Urban Environment. Acta Ornithol. 2012, 47, 37–45. [Google Scholar] [CrossRef]
- Hart, L.; Downs, C. Winged invaders: Bird introductions. Quest 2015, 11, 38–41. Available online: https://hdl.handle.net/10520/EJC172338 (accessed on 10 May 2023).
- Ruwandeniya, C.; Lill, A. Do exotic birds dominate feeding at garden food stations in Melbourne in winter? Corella 2016, 40, 91–97. [Google Scholar]
- Barua, M. Feral ecologies: The making of postcolonial nature in London. J. R. Anthr. Inst. 2022, 28, 896–919. [Google Scholar] [CrossRef]
- Gering, E.; Incorvaia, D.; Henriksen, R.; Conner, J.; Getty, T.; Wright, D. Getting Back to Nature: Feralization in Animals and Plants. Trends Ecol. Evol. 2019, 34, 1137–1151. [Google Scholar] [CrossRef]
- Hobson, E.A.; Smith-Vidaurre, G.; Salinas-Melgoza, A. History of nonnative Monk Parakeets in Mexico. PLoS ONE 2017, 12, e0184771. [Google Scholar] [CrossRef]
- Codesido, M.; Drozd, A. Alien birds in Argentina: Pathways, characteristics and ecological roles. Biol. Invasions 2021, 23, 1329–1338. [Google Scholar] [CrossRef]
- Liebl, A.L.; Schrey, A.W.; Andrew, S.C.; Sheldon, E.L.; Griffith, S.C. Invasion genetics: Lessons from a ubiquitous bird, the house sparrow Passer domesticus. Curr. Zool. 2015, 61, 465–476. [Google Scholar] [CrossRef]
- Del Hoyo, J.; Del Hoyo, J.; Elliott, A.; Sargatal, J. Handbook of the Birds of the World; Lynx Edicions: Barcelona, Spain, 1992; Volume 1. [Google Scholar]
- Spennemann, D.H.R.; Watson, M.J. Dietary habits of urban pigeons (Columba livia) and implications of excreta pH—A review. Eur. J. Ecol. 2017, 3, 27–41. [Google Scholar] [CrossRef]
- González-Acuña, D.; Silva, F.; Moreno, L.; Cerda, F.; Donoso, S.; Cabello, J.; López, J. Detección de algunos agentes zoonóticos en la paloma doméstica (Columba livia) en la ciudad de Chillán, Chile. Rev. Chil. Infectología 2007, 24, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, J.; Monsalve-Arcila, D.; Márquez-Villegas, C. Presencia de parásitos y enterobacterias en palomas ferales (Columba livia) en áreas urbanas en Envigado, Colombia. Rev. Fac. Nac. Salud Pública 2015, 33, 370–376. [Google Scholar] [CrossRef]
- MacGregor-Fors, I.; Morales-Pérez, L.; Quesada, J.; Schondube, J.E. Relationship between the presence of House Sparrows (Passer domesticus) and Neotropical bird community structure and diversity. Biol. Invasions 2010, 12, 87–96. [Google Scholar] [CrossRef]
- Caraboni, A.; Barrientos, C.; Fuentes, P. Registro de una colonia reproductiva de la cotorra argentina (Myiopsitta monachus) en la ciudad de Coronel, Región del Biobío, sur de Chile. Rev. Chil. Ornitol. 2020, 26, 96–100. [Google Scholar]
- Briceño, C.; Larraechea, M.; Alvarado, S. Monk Parakeet’s (Myiopsitta monachus) Ecological Parameters after Five Decades of Invasion in Santiago Metropolis, Chile. Birds 2022, 3, 341–358. [Google Scholar] [CrossRef]
- Briceño, C.; Sandoval-Rodríguez, A.; Yévenes, K.; Larraechea, M.; Morgado, A.; Chappuzeau, C.; Muñoz, V.; Dufflocq, P.; Olivares, F. Interactions between Invasive Monk Parakeets (Myiopsitta monachus) and Other Bird Species during Nesting Seasons in Santiago, Chile. Animals 2019, 9, 923. [Google Scholar] [CrossRef]
- Iriarte, J.A.; Lobos, G.A.; Jaksic, F. Invasive vertebrate species in Chile and their control and monitoring by governmental agencies. Rev. Chil. Hist. Nat. 2005, 78, 143–151. [Google Scholar] [CrossRef]
- Leong, M.; Dunn, R.R.; Trautwein, M.D. Biodiversity and socioeconomics in the city: A review of the luxury effect. Biol. Lett. 2018, 14, 20180082. [Google Scholar] [CrossRef]
- Lerman, S.B.; Warren, P.S. The conservation value of residential yards: Linking birds and people. Ecol. Appl. 2011, 21, 1327–1339. [Google Scholar] [CrossRef]
- Villaseñor, N.R.; Escobar, M.A.H. Linking Socioeconomics to Biodiversity in the City: The Case of a Migrant Keystone Bird Species. Front. Ecol. Evol. 2022, 10, 210. [Google Scholar] [CrossRef]
- Loss, S.R.; Ruiz, M.O.; Brawn, J.D. Relationships between avian diversity, neighborhood age, income, and environmental characteristics of an urban landscape. Biol. Conserv. 2009, 142, 2578–2585. [Google Scholar] [CrossRef]
- INE. Estimaciones y Proyecciones de la Población de Chile 2002–2035, Totales Regionales, Población Urbana y Rural; Instituto Nacional de Estadísticas: Santiago, Chile, 2019.
- Arroyo, M.T.; Marquet, P.; Marticorena, C.; Simonetti, J.; Cavieres, L.; Saqueo, F.; Massardo, F. El hotspot chileno, prioridad para la Biodiversidad mundial. In CONAMA. (2008). Biodiversidad de CHILE, Patrimonio y Desafíos; Ocho Libros Editores: Santiago, Chile, 2008; p. 640. [Google Scholar]
- Corcuera, E. Especies Exóticas Invasoras en Áreas Protegidas de Chile, Memoria Primer Encuentro Reserva Biológica Huilo Huilo, octubre—2014. In Proyecto GEF/MMA/PNUD: Fortalecimiento de los Marcos Nacionales para la Gobernabilidad de las Especies Exóticas Invasoras (2013–2017); Santiago, Chile, 2016; p. 100. Available online: https://especies-exoticas.mma.gob.cl/wp-content/uploads/2018/12/7.-Libro_final_EEI_encuentro_huilo_huilo_final_baja.pdf (accessed on 10 May 2023).
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Castri, F.D.; Hajek, E.R. Bioclimatología de Chile; University Católica de Chile: Santiago, Chile, 1976. [Google Scholar]
- GfK Chile. Estilo de Vida de Los Nuevos Grupos Socioeconómicos de Chile. 2019. Available online: https://www.gfk.com/fileadmin/user_upload/country_one_pager/CL/GfK_GSE_190502_FINAL.pdf (accessed on 18 January 2023).
- Lionello, P.; Malanotte-Rizzoli, P.; Boscolo, R.; Alpert, P.; Artale, V.; Li, L.; Luterbacher, J.; May, W.; Trigo, R.; Tsimplis, M. The Mediterranean climate: An overview of the main characteristics and issues. Dev. Earth Environ. Sci. 2006, 4, 1–26. [Google Scholar] [CrossRef]
- Tellería, J.L.; Fernández-López, J.; Fandos, G. Effect of Climate Change on Mediterranean Winter Ranges of Two Migratory Passerines. PLoS ONE 2016, 11, e0146958. [Google Scholar] [CrossRef]
- Herrera, C. A Study of Avian Frugivores, Bird-Dispersed Plants, and Their Interaction in Mediterranean Scrublands. Ecol. Monogr. 1984, 54, 1–23. [Google Scholar] [CrossRef]
- Muñoz-Pacheco, C.B.; Villaseñor, N.R. Avian species richness in cities: A review of the Spanish-language literature from the Southern Cone of South America. Urban Ecosyst. 2021, 25, 601–616. [Google Scholar] [CrossRef]
- Benito, J.F.; Escobar, M.A.H.; Villaseñor, N.R. Conservación en la ciudad: ¿Cómo influye la estructura del hábitat sobre la abundancia de especies de aves en una metrópoli latinoamericana? Gayana 2019, 83, 114–125. [Google Scholar] [CrossRef]
- Bartier, P.M.; Keller, C. Multivariate interpolation to incorporate thematic surface data using inverse distance weighting (IDW). Comput. Geosci. 1996, 22, 795–799. [Google Scholar] [CrossRef]
- Castro, S.A.; Leiva, N.G.; Bolados, M.; Figueroa, J.A. Riqueza y distribución de la flora urbana de Santiago de Chile: Una aproximación basada en interpolación IDW. Cad. Pesqui. 2018, 30, 41–54. [Google Scholar] [CrossRef]
- Gutiérrez-Tapia, P.; Azócar, M.I.; Castro, S.A. A citizen-based platform reveals the distribution of functional groups inside a large city from the Southern Hemisphere: E-Bird and the urban birds of Santiago (Central Chile). Rev. Chil. Hist. Nat. 2018, 91, 3. [Google Scholar] [CrossRef]
- García-Arroyo, M.; Gómez-Martínez, M.A.; MacGregor-Fors, I. The queen of the island: On the density and distribution of the Eurasian Collared-Dove (Streptopelia decaocto) in Cozumel. Urban Ecosyst. 2022, 25, 285–290. [Google Scholar] [CrossRef]
- Roberts, E.A.; Sheley, R.L.; Lawrence, R.L. Using sampling and inverse distance weighted modeling for mapping invasive plants. West. N. Am. Nat. 2004, 64, 312–323. [Google Scholar]
- Escobedo, F.J.; Palmas-Perez, S.; Dobbs, C.; Gezan, S.; Hernandez, J. Spatio-Temporal Changes in Structure for a Mediterranean Urban Forest: Santiago, Chile 2002 to 2014. Forests 2016, 7, 121. [Google Scholar] [CrossRef]
- Moran, P.A. The interpretation of statistical maps. J. R. Stat. Soc. Ser. B (Methodol.) 1948, 10, 243–251. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S.; Christensen, R.H.B.; Singmann, H.; Grothendieck, G. Package ‘lme4’. 2009. Available online: https://github.com/lme4/lme4/ (accessed on 2 December 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 5 April 2023).
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Muñoz-Pedreros, A.; González-Urrutia, M.; Encina-Montoya, F.; Norambuena, H.V. Effects of vegetation strata and human disturbance on bird diversity in green areas in a city in southern Chile. Avian Res. 2018, 9, 38. [Google Scholar] [CrossRef]
- Silva, C.P.; Sepúlveda, R.D.; Barbosa, O. Nonrandom filtering effect on birds: Species and guilds response to urbanization. Ecol. Evol. 2016, 6, 3711–3720. [Google Scholar] [CrossRef]
- Leveau, L.M.; Leveau, C.M. Comunidades de aves en un gradiente urbano de la ciudad de Mar del Plata, Argentina. El Hornero 2004, 19, 13–21. [Google Scholar] [CrossRef]
- Almazán-Núñez, R.C.; Hinterholzer-Rodríguez, A. Dinámica temporal de la avifauna en un parque urbano de la ciudad de Puebla, México. Huitzil 2010, 11, 26–32. [Google Scholar] [CrossRef]
- Sacco, A.G.; Bergmann, F.B.; Rui, A.M. Bird assemblages in the urban area in the city of Pelotas, Rio Grande do Sul, Brazil. Biota Neotrop. 2013, 13, 153–162. [Google Scholar] [CrossRef]
- Rico-Silva, J.F.; Cruz-Trujillo, E.J.; Colorado, Z.G.J. Influence of environmental factors on bird diversity in greenspaces in an Amazonian city. Urban Ecosyst. 2021, 24, 365–374. [Google Scholar] [CrossRef]
- Hager, S.B.; Cosentino, B.J.; McKay, K.J.; Monson, C.; Zuurdeeg, W.; Blevins, B. Window Area and Development Drive Spatial Variation in Bird-Window Collisions in an Urban Landscape. PLoS ONE 2013, 8, e53371. [Google Scholar] [CrossRef] [PubMed]
- Clergeau, P.; Savard, J.-P.L.; Mennechez, G.; Falardeau, G. Bird Abundance and Diversity along an Urban-Rural Gradient: A Comparative Study between Two Cities on Different Continents. Condor 1998, 100, 413–425. [Google Scholar] [CrossRef]
- Giunchi, D.; Albores-Barajas, Y.V.; Baldaccini, N.E.; Vanni, L.; Soldatini, C. Feral pigeons: Problems, dynamics and control methods. In Integrated Pest Management and Pest Control. Current and Future Tactics; InTechOpen: London, UK, 2012; pp. 215–240. [Google Scholar]
- De Laet, J.; Summers-Smith, J.D. The status of the urban house sparrow Passer domesticus in north-western Europe: A review. J. Ornithol. 2007, 148 (Suppl. S2), 275–278. [Google Scholar] [CrossRef]
- Jokimäki, J.; Suhonen, J.; Kaisanlahti-Jokimäki, M.-L. Differential Long-Term Population Responses of Two Closely Related Human-Associated Sparrow Species with Respect to Urbanization. Birds 2021, 2, 230–249. [Google Scholar] [CrossRef]
- Johnston, R.F. Evolution in the rock dove: Skeletal morphology. Auk 1992, 109, 530–542. [Google Scholar] [CrossRef]
- Shaw, L.M.; Chamberlain, D.; Evans, M.R. The House Sparrow Passer domesticus in urban areas: Reviewing a possible link between post-decline distribution and human socioeconomic status. J. Ornithol. 2008, 149, 293–299. [Google Scholar] [CrossRef]
- Strubbe, D.; Matthysen, E. Establishment success of invasive ring-necked and monk parakeets in Europe. J. Biogeogr. 2009, 36, 2264–2278. [Google Scholar] [CrossRef]
- Rodríguez-Pastor, R.; Barcelona, S.M.D.C.N.D.; Senar, J.C.; Ortega, A.; Faus, J.; Uribe, F.; Montalvo, T. Distribution patterns of invasive Monk parakeets (Myiopsitta monachus) in an urban habitat. Anim. Biodivers. Conserv. 2012, 35, 107–117. [Google Scholar] [CrossRef]
- Senar, J.C.; Carrillo-Ortiz, J.G.; Ortega-Segalerva, A.; Pell, F.S.E.D.; Pascual, J.; Arroyo, L.; Mazzoni, D.; Montalvo, T.; Hatchwell, B.J. The reproductive capacity of Monk Parakeets Myiopsitta monachus is higher in their invasive range. Bird Study 2019, 66, 136–140. [Google Scholar] [CrossRef]
- Martin-Etchegaray, A.; Esquivel, A.; Weiler, A. Estructura de las comunidades de aves de cuatro áreas verdes de la ciudad de Asunción, Paraguay. Rev. Cienc. Ambient. 2018, 52, 184–207. [Google Scholar] [CrossRef]
- Aramburú, R.y.V. Corbalán. Dieta de pichones de cotorra Myiopsitta monachus (Aves: Psittacidae) en una población silvestre. Ornitol. Neotrop. 2000, 11, 241–245. [Google Scholar]
- Yzurieta, D.; Naroski, T. Guía para la Identificación de las Aves de Argentina y Uruguay; Mazzini, V., Ed.; Zagier & Urruty Pubns: Buenos Aires, Argentina, 2003; 345p. [Google Scholar]
- Leger, E.A.; Rice, K.J. Invasive California poppies (Eschscholzia californica Cham.) grow larger than native individuals under reduced competition. Ecol. Lett. 2003, 6, 257–264. [Google Scholar] [CrossRef]
- Sacchi, R.; Gentilli, A.; Razzetti, E.; Barbieri, F. Effects of building features on density and flock distribution of feral pigeons Columba livia var. domestica in an urban environment. Can. J. Zool. 2002, 80, 48–54. [Google Scholar] [CrossRef]
- Buijs, J.A.; Van Wijnen, J.H. Survey of feral rock doves (Columba livia) in Amsterdam, a bird-human association. Urban Ecosyst. 2001, 5, 235–241. [Google Scholar] [CrossRef]
- De La Ossa, J.; La Ossa-Lacayo, D.; Monroy-Pineda, M. Abundance of domestic dove (Columbia livia domestica Gmelin, 1789) in Santiago de Tolú, Sucre, Colombia. Rev. MVZ Córdoba 2017, 22, 5718–5727. [Google Scholar] [CrossRef]
- Haag-Wackernagel, D.; Geigenfeind, I. Protecting buildings against feral pigeons. Eur. J. Wildl. Res. 2008, 54, 715–721. [Google Scholar] [CrossRef]
- Zarricueta, R.T. Vivienda social en Santiago de Chile: Análisis de su comportamiento locacional, período 1980–2002. Rev. INVI 2011, 26, 105–131. [Google Scholar] [CrossRef]
- Borsdorf, A.; Hidalgo, R. New dimensions of social exclusion in Latin America: From gated communities to gated cities, the case of Santiago de Chile. Land Use Policy 2008, 25, 153–160. [Google Scholar] [CrossRef]
- Escobedo, F.J.; Nowak, D.J.; Wagner, J.E.; De la Maza, C.L.; Rodríguez, M.; Crane, D.E.; Hernández, J. The socioeconomics and management of Santiago de Chile’s public urban forests. Urban For. Urban Green 2006, 4, 105–114. [Google Scholar] [CrossRef]
- Hernández, H.J.; Villaseñor, N.R. Twelve-year change in tree diversity and spatial segregation in the Mediterranean city of Santiago, Chile. Urban For. Urban Green 2018, 29, 10–18. [Google Scholar] [CrossRef]
- Figueroa, J.A.; Saldías, M.G.; Lagos, D.; Teillier, S.; Castro, S.A. Soil seed banks are short-lived and triggered by the first effective rainfalls in the vacant lots of Santiago, Chile. Urban For. Urban Green 2022, 67, 127418. [Google Scholar] [CrossRef]
- Whittingham, M.J.; Evans, K.L. The effects of habitat structure on predation risk of birds in agricultural landscapes. Ibis 2004, 146, 210–220. [Google Scholar] [CrossRef]
- Foncea, J.F.; Villaseñor, N.R.; Escobar, M.A. Respuestas de la comunidad de aves a las variables de hábitat local y del paisaje en la ciudad de Santiago de Chile. Ecol. Austral. 2023, 33, 455–468. [Google Scholar]
- Bucher, E.H.; Aramburú, R.M. Land-use changes and monk parakeet expansion in the Pampas grasslands of Argentina. J. Biogeogr. 2014, 41, 1160–1170. [Google Scholar] [CrossRef]
- Sol, D.; Santos, D.M.; Clavell, J. Habitat Selection by the Monk Parakeet during Colonization of a New Area in Spain. Condor 1997, 99, 39–46. [Google Scholar] [CrossRef]
- Fernández, I.C.; Wu, J. A GIS-based framework to identify priority areas for urban environmental inequity mitigation and its application in Santiago de Chile. Appl. Geogr. 2018, 94, 213–222. [Google Scholar] [CrossRef]
- Buitrago-Medina, D.A.; Méndez-Mancera, V.; Jimenez, L.C.V.; Soler-Tovar, D. La paloma (Columba livia) en la transmisión de enfermedades de importancia en salud pública. Rev. Cienc. Anim. 2013, 6, 177–194. [Google Scholar]
- Vasconcelos, R.H.; Teixeira, R.S.D.C.; Da Silva, I.N.G.; Lopes, E.D.S.; Maciel, W.C. Feral pigeons (Columba livia) as potential reservoirs of Salmonella sp. and Escherichia coli. Arq. Do Inst. Biológico 2018, 85, 1–6. [Google Scholar] [CrossRef]
- Cong, W.; Huang, S.-Y.; Zhou, N.-H.; Zhang, X.-X.; Zhang, N.-Z.; Zhao, Q.; Zhu, X.-Q. Prevalence and Genetic Characterization of Toxoplasma gondii in House Sparrows (Passer domesticus) in Lanzhou, China. Korean J. Parasitol. 2013, 51, 363–367. [Google Scholar] [CrossRef]
- Abdoli, A.; Dalimi, A.; Soltanghoraee, H.; Ghaffarifar, F. Molecular detection of Toxoplasma gondii in house sparrow (Passer domesticus) by LAMP and PCR methods in Tehran, Iran. J. Parasit. Dis. 2016, 40, 1317–1321. [Google Scholar] [CrossRef]
- Dubey, J.P. The History of Toxoplasma gondii—The First 100 Years. J. Eukaryot. Microbiol. 2008, 55, 467–475. [Google Scholar] [CrossRef]
- Briceño, C.; Surot, D.; González-Acuña, D.; Martinez-Moreno, F.; Fredes, F. Parasitic survey on introduced monk parakeets (Myiopsitta monachus) in Santiago, Chile. Rev. Bras. Parasitol. Vet. 2017, 26, 129–135. [Google Scholar] [CrossRef]
- Di Santo, M.; Battisti, C.; Bologna, M.A. Interspecific interactions in nesting and feeding urban sites among introduced Monk Parakeet (Myiopsitta monachus) and syntopic bird species. Ethol. Ecol. Evol. 2017, 29, 138–148. [Google Scholar] [CrossRef]
- Villaseñor, N.R.; Escobar, M.A.H. Cemeteries and biodiversity conservation in cities: How do landscape and patch-level attributes influence bird diversity in urban park cemeteries? Urban Ecosyst. 2019, 22, 1037–1046. [Google Scholar] [CrossRef]
- Amaya-Espinel, J.D.; Hostetler, M.E. The value of small forest fragments and urban tree canopy for Neotropical migrant birds during winter and migration seasons in Latin American countries: A systematic review. Landsc. Urban Plan. 2019, 190, 103592. [Google Scholar] [CrossRef]
- Bednarczuk, E.; Feare, C.J.; Lovibond, S.; Tatayah, V.; Jones, C.G. Attempted eradication of house sparrows Passer domesticus from Round Island (Mauritius), Indian Ocean. Conserv. Evid. 2010, 7, 75–86. [Google Scholar]
- Ramírez-Cruz, G.A.; Ortega-Álvarez, R. Identifying management guidelines to control the invasive House Sparrow (Passer domesticus) within natural protected areas through the estimation of local colonization and extinction probabilities. Biol. Invasions 2021, 23, 3767–3776. [Google Scholar] [CrossRef]
- Farfán, M.; Díaz-Ruiz, F.; Duarte, J.; Real, R. Feral pigeon (Columba livia var. domestica) management in low-density urban areas: Prevention is better than cure. Urban Ecosyst. 2019, 22, 1027–1035. [Google Scholar] [CrossRef]
- Albonetti, P.; Marletta, A.; Repetto, I.; Sasso, E. Efficacy of nicarbazin (Ovistop®) in the containment and reduction of the populations of feral pigeons (Columba livia var. domestica) in the city of Genoa, Italy: A retrospective evaluation. Vet. Ital. 2015, 51, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Dobeic, M.; Pintarič, Š.; Vlahović, K.; Dovč, A. Feral pigeon (Columba livia) population management in Ljubljana. Vet. Arh. 2011, 81, 285–298. [Google Scholar]
- Avery, M.L.; Yoder, C.A.; Tillman, E.A. Diazacon Inhibits Reproduction in Invasive Monk Parakeet Populations. J. Wildl. Manag. 2008, 72, 1449–1452. [Google Scholar] [CrossRef]
- Yoder, C.A.; Avery, M.L.; Keacher, K.L.; Tillman, E.A. Use of DiazaCon™ as a reproductive inhibitor for monk parakeets (Myiopsitta monachus). Wildl. Res. 2007, 34, 8–13. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Common Name | Scientific Name | Autumn | Winter |
|---|---|---|---|---|
| Columbidae | Domestic pigeon | Columba livia f. domestica | 3.7 ± 0.5 | 3.1 ± 0.4 |
| Passeridae | House sparrow | Passer domesticus | 2.7 ± 0.2 | 2.4 ± 0.2 |
| Psittacidae | Monk parakeet | Myiopsitta monachus | 0.9 ± 0.2 | 0.9 ± 0.2 |
| Species | Predictive Variables | Estimated Coefficient | Standard Error | p-Value | |
|---|---|---|---|---|---|
| C. livia f. domestica | Intercept | −0.02 | 0.23 | 0.940 | |
| Socioeconomic Medium | 0.20 | 0.31 | 0.510 | ||
| Socioeconomic Low | 1.42 | 0.30 | <0.001 | *** | |
| Season Winter | 0.01 | 0.04 | 0.868 | ||
| P. domesticus | Intercept | −1.87 | 0.30 | <0.001 | *** |
| Socioeconomic Medium | 2.48 | 0.33 | <0.001 | *** | |
| Socioeconomic Low | 3.15 | 0.34 | <0.001 | *** | |
| Season Winter | −0.11 | 0.06 | 0.056 | . | |
| M. monachus | Intercept | −1.10 | 0.58 | 0.057 | . |
| Socioeconomic Medium | −2.01 | 0.79 | 0.011 | * | |
| Socioeconomic Low | −3.76 | 0.85 | <0.001 | *** | |
| Season Winter | 0.03 | 0.09 | 0.778 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva-Ortega, M.; Muñoz-Pacheco, C.B.; Villaseñor, N.R. Abundance of Non-Native Birds in the City: Spatial Variation and Relationship with Socioeconomics in a South American City. Animals 2023, 13, 1737. https://doi.org/10.3390/ani13111737
Silva-Ortega M, Muñoz-Pacheco CB, Villaseñor NR. Abundance of Non-Native Birds in the City: Spatial Variation and Relationship with Socioeconomics in a South American City. Animals. 2023; 13(11):1737. https://doi.org/10.3390/ani13111737
Chicago/Turabian StyleSilva-Ortega, Macarena, Catalina B. Muñoz-Pacheco, and Nélida R. Villaseñor. 2023. "Abundance of Non-Native Birds in the City: Spatial Variation and Relationship with Socioeconomics in a South American City" Animals 13, no. 11: 1737. https://doi.org/10.3390/ani13111737