
Improving Rabbit Doe Metabolism and Whole Reproductive Cycle Outcomes via Fatty Acid-Rich Moringa oleifera Leaf Extract Supplementation in Free and Nano-Encapsulated Forms



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Moringa oleifera Leaf Extraction and Nanofabrication
2.2. Nano-Encapsulated ME Physicochemical Properties and Fatty Acid Profile
2.3. Animal Management and Experimental Design
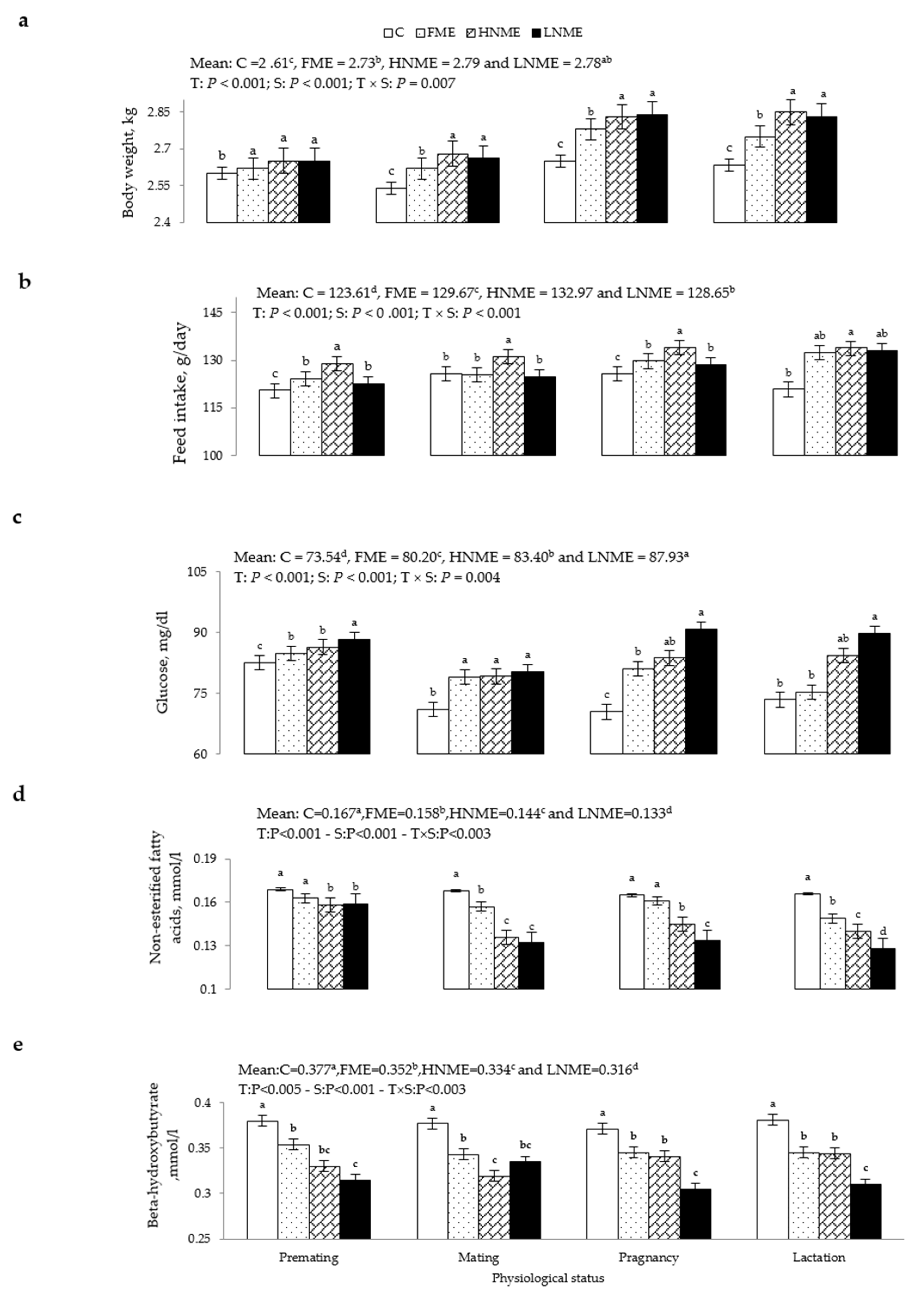
2.4. Physiological Variables
2.5. Blood Sampling and Analysis
2.5.1. Blood Biochemical Attributes
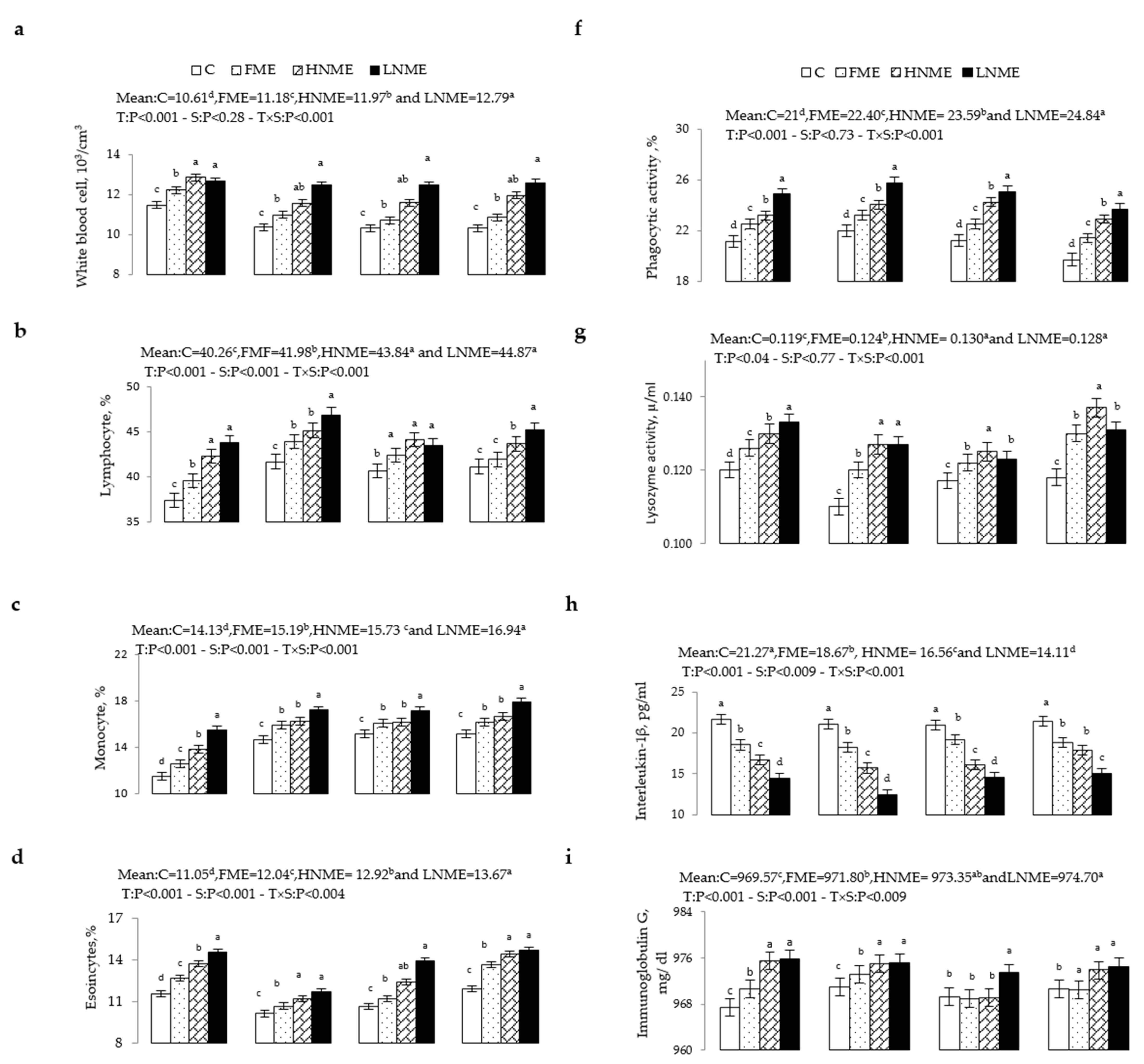
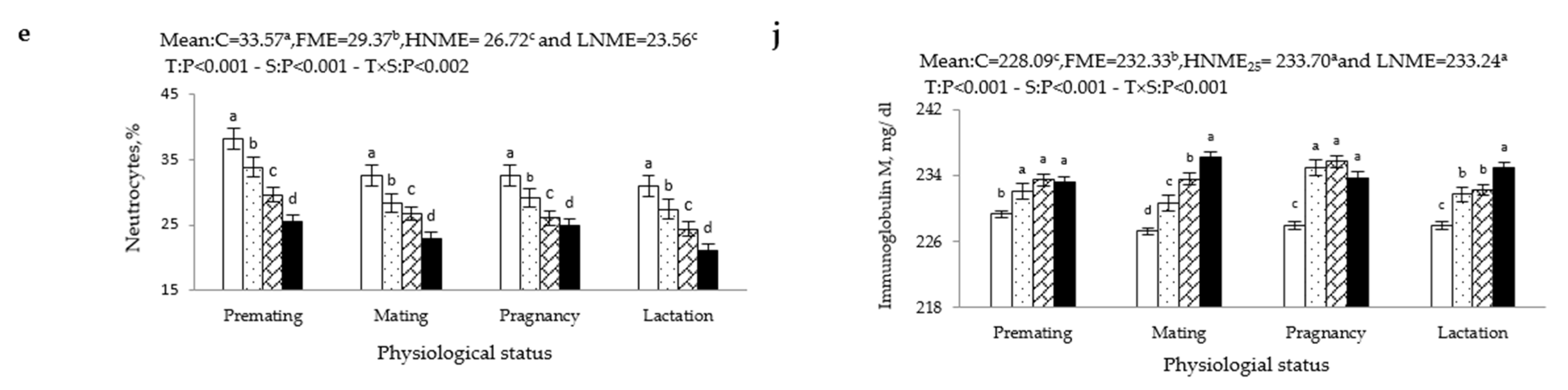
2.5.2. Immune Variables
2.6. Colostrum and Milk Analysis
2.6.1. Colostrum Collection and Analysis
2.6.2. Milk Collection and Analysis
2.7. Productive and Reproductive Performance
2.8. Statistical Analysis
3. Results
3.1. Fatty Acid Profile of ME and EE
3.2. BW, Feed Intake, and Energy-Related Metabolites
3.3. Blood Immune Variables
3.4. Colostrum Immunoglobulin, Milk Yield, and Milk Composition
3.5. Milk Fatty Acid Profile and Fatty Acid Health Indices
3.6. Reproductive Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maertens, L.; Lebas, F.; Szendro, Z. Rabbit milk: A review of quantity, quality and non-dietary affecting factors. World Rabbit. Sci. 2006, 14, 205–230. [Google Scholar] [CrossRef]
- Castellini, C.; Dal Bosco, A.; Arias-Álvarez, M.; Lorenzo, P.L.; Cardinali, R.; Rebollar, P.G. The main factors affecting the reproductive performance of rabbit does: A review. Anim. Reprod. Sci. 2010, 122, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Menchetti, L.; Barbato, O.; Sforna, M.; Vigo, D.; Mattioli, S.; Curone, G.; Tecilla, M.; Riva, F.; Brecchia, G. Effects of diets enriched in linseed and fish oil on the expression pattern of toll-like receptors 4 and proinflammatory cytokines on gonadal axis and reproductive organs in rabbit buck. Oxidative Med. Cell. Longev. 2020, 2020, 4327470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellini, C. Reproductive activity and welfare of rabbit does. Ital. J. Anim. Sci. 2007, 6 (Suppl. 1), 743–747. [Google Scholar] [CrossRef]
- Martínez-Paredes, E.; Ródenas, L.; Martínez-Vallespín, B.; Cervera, C.; Blas, E.; Brecchia, G.; Boiti, C.; Pascual, J. Effects of feeding programme on the performance and energy balance of nulliparous rabbit does. Animal 2012, 6, 1086–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchiani, S.; Vignozzi, L.; Filippi, S.; Gurrieri, B.; Comeglio, P.; Morelli, A.; Danza, G.; Bartolucci, G.; Maggi, M.; Baldi, E. Metabolic syndrome-associated sperm alterations in an experimental rabbit model: Relation with metabolic profile, testis and epididymis gene expression and effect of tamoxifen treatment. Mol. Cell. Endocrinol. 2015, 401, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.; Vallenari, A.; Prusti, T.; de Bruijne, J.; Babusiaux, C.; Biermann, M.; Creevey, O.; Evans, D.; Eyer, L.; Hutton, A. Gaia Early Data Release 3-Summary of the contents and survey properties (Corrigendum). Astron. Astrophys. 2021, 650, C3. [Google Scholar]
- Fortun-Lamothe, L. Energy balance and reproductive performance in rabbit does. Anim. Reprod. Sci. 2006, 93, 1–15. [Google Scholar] [CrossRef]
- Mateo, R.; Carroll, J.; Hyun, Y.; Smith, S.; Kim, S. Effect of dietary supplementation of n-3 fatty acids and elevated concentrations of dietary protein on the performance of sows. J. Anim. Sci. 2009, 87, 948–959. [Google Scholar] [CrossRef]
- Al-Khalaifah, H. Modulatory effect of dietary polyunsaturated fatty acids on immunity, represented by phagocytic activity. Front. Vet. 2020, 7, 672. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Karpe, F. Determinants of VLDL-triglycerides production. Curr. Opin. Lipidol. 2012, 23, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Hadjadj, I.; Hankele, A.-K.; Armero, E.; Argente, M.-J.; de la Luz García, M. Fatty Acid Profile of Blood Plasma at Mating and Early Gestation in Rabbit. Animals 2021, 11, 3200. [Google Scholar] [CrossRef] [PubMed]
- Rooke, J.; Sinclair, A.; Edwards, S. Feeding tuna oil to the sow at different times during pregnancy has different effects on piglet long-chain polyunsaturated fatty acid composition at birth and subsequent growth. Br. J. Nutr. 2001, 86, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betancourt López, C.A.; Bernal Santos, M.G.; Vázquez Landaverde, P.A.; Bauman, D.E.; Harvatine, K.J.; Srigley, C.T.; Becerra Becerra, J. Both Dietary Fatty Acids and Those Present in the Cecotrophs Contribute to the Distinctive Chemical Characteristics of New Zealand Rabbit Milk Fat. Lipids 2018, 53, 1085–1096. [Google Scholar] [CrossRef] [PubMed]
- Rebollar, P.; García-García, R.; Arias-Álvarez, M.; Millán, P.; Rey, A.I.; Rodríguez, M.; Formoso-Rafferty, N.; De la Riva, S.; Masdeu, M.; Lorenzo, P. Reproductive long-term effects, endocrine response and fatty acid profile of rabbit does fed diets supplemented with n-3 fatty acids. Anim. Reprod. Sci. 2014, 146, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, M.; García-García, R.; Arias-Álvarez, M.; Formoso-Rafferty, N.; Millán, P.; López-Tello, J.; Lorenzo, P.; González-Bulnes, A.; Rebollar, P. A diet supplemented with n-3 polyunsaturated fatty acids influences the metabomscic and endocrine response of rabbit does and their offspring. J. Anim. Sci. 2017, 95, 2690–2700. [Google Scholar] [PubMed]
- Rodríguez, M.; García-García, R.; Arias-Álvarez, M.; Millán, P.; Febrel, N.; Formoso-Rafferty, N.; López-Tello, J.; Lorenzo, P.; Rebollar, P. Improvements in the conception rate, milk composition and embryo quality of rabbit does after dietary enrichment with n-3 polyunsaturated fatty acids. Animal 2018, 12, 2080–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sri, W.; Sri, W.; Hendrawan, S. The effects of adding Moringa oleifera leaves extract on rabbit does’ milk production and mammary gland histology. Russ. J. Agric. Socio-Econ. Sci. 2019, 8, 296–304. [Google Scholar]
- Saini, R.; Shetty, N.; Prakash, M.; Giridhar, P. Effect of dehydration methods on retention of carotenoids, tocopherols, ascorbic acid and antioxidant activity in Moringa oleifera leaves and preparation of a RTE product. J. Food Sci. Technol. 2014, 51, 2176–2182. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, L.; Doriya, K.; Kumar, D.S. Moringa oleifera: A review on nutritive importance and its medicinal application. Food Sci. Hum. Wellness 2016, 5, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, P.A.; Morón, B.; Becker, H.M.; Lang, S.; Atrott, K.; Spalinger, M.R.; Scharl, M.; Wojtal, K.A.; Fischbeck-Terhalle, A.; Frey-Wagner, I. Titanium dioxide nanoparticles exacerbate DSS-induced colitis: Role of the NLRP3 inflammasome. Gut 2017, 66, 1216–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Desoky, N.I.; Hashem, N.M.; Gonzalez-Bulnes, A.; Elkomy, A.G.; Abo-Elezz, Z.R. Effects of a Nanoencapsulated Moringa Leaf Ethanolic Extract on the Physiology, Metabolism and Reproductive Performance of Rabbit Does during Summer. Antioxidants 2021, 10, 1326. [Google Scholar] [CrossRef]
- Egan, W.F. Frequency Synthesis by Phase Lock; Wiley: New York, NY, USA, 1981. [Google Scholar]
- Tsiplakou, E.; Mountzouris, K.; Zervas, G. Concentration of conjugated linoleic acid in grazing sheep and goat milk fat. Livest. Sci. 2006, 103, 74–84. [Google Scholar] [CrossRef]
- Estany, J.; Baselga, M.; Blasco, A.; Camacho, J. Mixed model methodology for the estimation of genetic response to selection in litter size of rabbits. Livest. Prod. Sci. 1989, 21, 67–75. [Google Scholar] [CrossRef]
- Carmichael, W.; Gorham, P.; Biggs, D. Two laboratory case studies on the oral toxicity to calves of the freshwater cyanophyte (blue-green alga) Anabaena flos-aquae NRC-44-1. Can. Vet. J. 1977, 18, 71. [Google Scholar] [PubMed]
- Hashem, N.; Aboul-Ezz, Z. Effects of a single administration of different gonadotropins on day 7 post-insemination on pregnancy outcomes of rabbit does. Theriogenology 2018, 105, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hosny, N.S.; Hashem, N.M.; Morsy, A.S.; Abo-Elezz, Z.R. Effects of organic selenium on the physiological response, blood metabolites, redox status, semen quality, and fertility of rabbit bucks kept under natural heat stress conditions. Front. Vet. Sci. 2020, 7, 290. [Google Scholar] [CrossRef] [PubMed]
- Hashem, N.M.; Shehata, M.G. Antioxidant and Antimicrobial Activity of Cleomedroserifolia (Forssk.) Del. and Its Biological Effects on Redox Status, Immunity, and Gut Microflora. Animals 2021, 11, 1929. [Google Scholar] [CrossRef] [PubMed]
- Jimidar, M.; Hartmann, C.; Cousement, N.; Massart, D. Determination of nitrate and nitrite in vegetables by capillary electrophoresis with indirect detection. J. Chromatogr. A 1995, 706, 479–492. [Google Scholar] [CrossRef]
- Xiccato, G.; Parigi-Bini, R.; Dalle Zotte, A.; Carazzolo, A.; Cossu, M. Effect of dietary energy level, addition of fat and physiological state on performance and energy balance of lactating and pregnant rabbit does. Anim. Sci. 1995, 61, 387–398. [Google Scholar] [CrossRef]
- Rijo, P.; Matias, D.; Fernandes, A.S.; Simões, M.F.; Nicolai, M.; Reis, C.P. Antimicrobial plant extracts encapsulated into polymeric beads for potential application on the skin. Polymers 2014, 6, 479–490. [Google Scholar] [CrossRef]
- El-Desoky, N.; Hashem, N.; Elkomy, A.; Abo-Elezz, Z. Physiological response and semen quality of rabbit bucks supplemented with Moringa leaves ethanolic extract during summer season. Animal 2017, 11, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Sun, X.; Guo, X.; Zhu, W.; Wu, J.; Chen, L.; Wang, J.; Chen, M.; Cheng, C.; Zhang, Q. Green synthesis, characterization and in vitro release of cinnamaldehyde/sodium alginate/chitosan nanoparticles. Food Hydrocoll. 2019, 90, 515–522. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Lee, W.; Han, E.J.; Ahn, G. Alginate-based nanomaterials: Fabrication techniques, properties, and applications. Chem. Eng. J. 2020, 391, 123823. [Google Scholar] [CrossRef]
- Hashem, N.M.; Hosny, N.S.; El-Desoky, N.I.; Shehata, M.G. Effect of Nanoencapsulated Alginate-Synbiotic on Gut Microflora Balance, Immunity, and Growth Performance of Growing Rabbits. Polymers 2021, 13, 4191. [Google Scholar] [CrossRef] [PubMed]
- Chankuang, P.; Linlawan, A.; Junda, K.; Kuditthalerd, C.; Suwanprateep, T.; Kovitvadhi, A.; Chundang, P.; Sanyathitiseree, P.; Yinharnmingmongkol, C. Comparison of Rabbit, Kitten and Mammal Milk Replacer Efficiencies in Early Weaning Rabbits. Animals 2020, 10, 1087. [Google Scholar] [CrossRef] [PubMed]
- Hashem, N.; Soltan, Y.; El-Desoky, N.; Morsy, A.; Sallam, S. Effects of Moringa oleifera extracts and monensin on performance of growing rabbits. Livest. Sci. 2019, 228, 136–143. [Google Scholar] [CrossRef]
- Filly, A.; Fernandez, X.; Minuti, M.; Visinoni, F.; Cravotto, G.; Chemat, F. Solvent-free microwave extraction of essential oil from aromatic herbs: From laboratory to pilot and industrial scale. Food Chem. 2014, 150, 193–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, I.U.; Sajid, S.; Javed, A.; Sajid, S.; Shah, S.U. Comparative diagnosis of typhoid fever by polymerase chain reaction and widal test in Southern Districts (Bannu, Lakki Marwat and DI Khan) of Khyber Pakhtunkhwa, Pakistan. Acta Sci. Malays. 2017, 1, 12–15. [Google Scholar] [CrossRef]
- Khalid, A.R.; Yasoob, T.B.; Zhang, Z.; Yu, D.; Feng, J.; Zhu, X.; Hang, S. Supplementation of Moringa oleifera leaf powder orally improved productive performance by enhancing the intestinal health in rabbits under chronic heat stress. J. Therm. Biol. 2020, 93, 102680. [Google Scholar] [CrossRef] [PubMed]
- Lennarz, W.J.; Lane, M.D. Encyclopedia of Biological Chemistry; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- El-Gazzar, H.I.; Gad, S.S.; Mubarak, M.H.; El-Deep, G.S.; GabAlla, A.A. Effect of Nitrogen fertilizer Levels and Six Canola Genotypes on Physicochemical Properties of (Brassica napus L.) Cultivated under North Sinai Conditions. Sinai J. Appl. Sci. 2019, 8, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Mousa, S. Boosting Refugee Outcomes: Evidence from Policy, Academia, and social Innovation; Academia, and Social Innovation (2 October 2018); Stanford Immigration Policy Lab: Stanford, CA, USA, 2018. [Google Scholar]
- Isitua, C.; Ibeh, I. Toxicological assessment of aqueous extract of Moringa oleifera and Caulis bambusae leaves in rabbits. J. Clin. Toxicol. S 2013, 12, 4. [Google Scholar]
- Rodríguez, M.; Rebollar, P.G.; Mattioli, S.; Castellini, C. n-3 PUFA sources (precursor/products): A review of current knowledge on rabbit. Animals 2019, 9, 806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Li, Y.; Pan, X.; Nguyen, D.N.; Brunse, A.; Bojesen, A.M.; Rudloff, S.; Mortensen, M.S.; Burrin, D.G.; Sangild, P.T. Human Milk Fortification with Bovine Colostrum Is Superior to Formula-Based Fortifiers to Prevent Gut Dysfunction, Necrotizing Enterocolitis, and Systemic Infection in Preterm Pigs. J. Parenter. Enter. Nutr. 2019, 43, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Declerck, I.; Dewulf, J.; Piepers, S.; Decaluwé, R.; Maes, D. Sow and litter factors influencing colostrum yield and nutritional composition. J. Anim. Sci. 2015, 93, 1309–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alameda-Pineda, X.; Horaud, R. A geometric approach to sound source localization from time-delay estimates. IEEE/ACM Trans. Audio Speech Lang. Process. 2014, 22, 1082–1095. [Google Scholar] [CrossRef] [Green Version]
- Adesina, O.; Oladapo, O.; Adeniran, O.; Obehioye, O.; Iyabode, O. Blood transfusion in obstetrics: Attitude and perceptions of pregnant women. Trop. J. Obstet. Gynaecol. 2014, 31, 38–44. [Google Scholar]
- Hewitson, J.P.; Grainger, J.R.; Maizels, R.M. Helminth immunoregulation: The role of parasite secreted proteins in modulating host immunity. Mol. Biochem. Parasitol. 2009, 167, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Benayad, A.; Taghouti, M.; Benali, A.; Aboussaleh, Y.; Benbrahim, N. Nutritional and technological assessment of durum wheat-faba bean enriched flours, and sensory quality of developed composite bread. Saudi J. Biol. Sci. 2021, 28, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Balthazar, C.F.; Guimarães, J.T.; Rocha, R.S.; Pimentel, T.C.; Neto, R.P.; Tavares, M.I.B.; Graça, J.S.; Alves Filho, E.G.; Freitas, M.Q.; Esmerino, E.A. Nuclear magnetic resonance as an analytical tool for monitoring the quality and authenticity of dairy foods. Trends Food Sci. Technol. 2021, 108, 84–91. [Google Scholar] [CrossRef]
- De Mattos, A.M.; Olyaei, A.J.; Bennett, W.M. Nephrotoxicity of immunosuppressive drugs: Long-term consequences and challenges for the future. Am. J. Kidney Dis. 2000, 35, 333–346. [Google Scholar] [CrossRef]
- Cerri, R.L.; Rutigliano, H.M.; Chebel, R.C.; Santos, J.E. Period of dominance of the ovulatory follicle influences embryo quality in lactating dairy cows. Reproduction 2009, 137, 813–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpato, S.; Cavalieri, M.; Guerra, G.; Sioulis, F.; Ranzini, M.; Maraldi, C.; Fellin, R.; Guralnik, J.M. Performance-based functional assessment in older hospitalized patients: Feasibility and clinical correlates. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2008, 63, 1393–1398. [Google Scholar] [CrossRef] [PubMed]
- Escamilla, A.; Bautista, M.; Zafra, R.; Pacheco, I.; Ruiz, M.T.; Martínez-Cruz, S.; Méndez, A.; Martínez-Moreno, A.; Molina-Hernández, V.; Pérez, J. Fasciola hepatica induces eosinophil apoptosis in the migratory and biliary stages of infection in sheep. Vet. Parasitol. 2016, 216, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Mark, P.J.; Mori, T.A.; Keelan, J.A.; Waddell, B.J. Maternal dietary omega-3 fatty acid supplementation reduces placental oxidative stress and increases fetal and placental growth in the rat. Biol. Reprod. 2013, 88, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Herrera, E.; Ortega-Senovilla, H. Lipid metabolism during pregnancy and its implications for fetal growth. Curr. Pharm. Biotechnol. 2014, 15, 24–31. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| FAs | FA, g/100 g FA Methyl Esters |
|---|---|
| Caprylic acid methyl ester, C8:0 | 5.30 |
| Capric acid methyl ester, C10:0 | 0.61 |
| Undecanoic acid methyl ester, C11:0 | 1.01 |
| Lauric acid methyl ester, C12:0 | 0.63 |
| Tridecanoic acid methyl ester, C13:0 | 0.64 |
| Myristic acid methyl ester, C14:0 | 1.18 |
| Pentadecanoic acid methyl ester, C15:0 | 0.81 |
| Palmitic acid methyl ester, C16:0 | 11.45 |
| Heptadecanoic acid methyl ester, C17:0 | 1.45 |
| Stearic acid methyl ester, C18:0 | 3.72 |
| Arachidic acid methyl ester, C20:0 | 1.97 |
| Heneicosanoic acid methyl ester, C21:0 | 2.55 |
| Behenoic acid methyl ester, C22:0 | 5.14 |
| Tricosanoic acid methyl ester, C23:0 | 3.26 |
| Lignoceric acid methyl ester, C24:0 | 6.02 |
| Myristoleic acid methyl ester, C14:1n-9 | 1.53 |
| Pentadecenoic acid methyl ester, C15:1n-5 | 1.71 |
| Palmitoleic acid methyl ester, C16:1n-7 | 2.12 |
| Heptadecenoic acid methyl ester, C17:1n-7 | 2.00 |
| Oleic acid methyl ester, C18:1n-9 | 9.04 |
| Elaidic acid methyl ester, C18:1n-9t | 3.82 |
| Eicosenoic acid methyl ester, C20:1n-9 | 2.62 |
| Erucic acid methyl ester, C22:1n-9 | 2.57 |
| Nervonic acid methyl ester, C24:1n-9 | 3.93 |
| Linolenic acid methyl ester (LA), C18:2n-6 | 3.68 |
| Gama-Linolenic acid methyl ester (GLA), C18:3n-6 | 5.98 |
| Dihomo-gamma-linolenic acid (DGLA), C20:4n-6 | 3.89 |
| Eicosatrienoic acid methyl ester (ETE), C20:3n-3 | 3.53 |
| Eicosapentaenoic acid methyl ester (EPA), C20:5n-3 | 3.40 |
| Docosahexaenoic acid methyl ester (DHA), C22:6n-3 | 4.45 |
| Saturated fatty acid | 45.73 |
| Unsaturated fatty acid | 54.27 |
| Mono-unsaturated fatty acid | 29.34 |
| Poly-unsaturated fatty acid | 24.93 |
| Poly-unsaturated fatty acid/Saturated fatty acid | 0.54 |
| Total odd FAs | 13.43 |
| Omega-3 FAs | 11.38 |
| Omega-6 FAs | 13.55 |
| Omega-9 FAs | 21.98 |
| Omega-6 FAs/Omega-3 FAs | 1.19 |
| FAs Category | Encapsulation Efficiency 1, % |
|---|---|
| Saturated FAs | 71.03 |
| Unsaturated FAs | 69.62 |
| Mono-unsaturated FAs | 77.64 |
| Poly-unsaturated FAs | 60.26 |
| Total odd FAs | 80.45 |
| Omega-3 FAs | 71.4 |
| Omega-6 FAs | 49.1 |
| Omega-9 FAs | 68.70 |
| Variable | Treatment 1 | SEM | p Value | |||
|---|---|---|---|---|---|---|
| C | FME | HNME | LNME | |||
| Colostrum immunoglobulin, mg/dL | ||||||
| Immunoglobulin M | 225.32 c | 230.85 b | 237.15 a | 232.14 b | 2.27 | 0.001 |
| Immunoglobulin A | 73.22 b | 75.23 a | 76.89 a | 75.56 a | 0.91 | 0.001 |
| Immunoglobulin G | 964.20 c | 968.21 b | 972.54 a | 968.23 b | 2.35 | 0.001 |
| Immunoglobulin E | 12.99 | 13.22 | 13.52 | 13.39 | 0.07 | 0.160 |
| Immunoglobulin D | 28.86 c | 31.83 b | 35.52 a | 35.53 a | 0.82 | 0.001 |
| Milk yield and composition | ||||||
| Milk yield, g/day | 117.50 c | 159.44 b | 161.52 ab | 169.86 a | 2.46 | 0.001 |
| Milk Composition, % | ||||||
| Protein | 11.79 d | 12.33 c | 12.68 b | 13.39 a | 0.02 | 0.001 |
| Fat | 13.38 | 13.50 | 14.12 | 14.69 | 0.33 | 0.07 |
| Total solids | 27.91 c | 29.54 bc | 31.77 ab | 32.51 a | 2.23 | 0.001 |
| Energy, MJ/kg | 8.50 c | 8.56 bc | 9.04 ab | 9.41 a | 0.06 | 0.005 |
| FAs, g/100 g FA Methyl Esters | Treatment 1 | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| C | FME | HNME | LNME | |||
| Butyric acid, C4:0 | 0.101 | 0.112 | 0.110 | 0.108 | 0.002 | 0.616 |
| Caproic acid methyl ester, C6:0 | 0.410 | 0.410 | 0.411 | 0.403 | 0.001 | 0.378 |
| Caprylic acid methyl ester, C8:0 | 25.59 | 25.97 | 26.27 | 26.13 | 0.11 | 0.167 |
| Capric acid methyl ester, C10:0 | 21.76 | 21.59 | 21.74 | 22.18 | 0.12 | 0.476 |
| Lauric acid methyl ester, C12:0 | 2.66 | 2.67 | 2.77 | 2.79 | 0.08 | 0.938 |
| Myristic acid methyl ester, C14:0 | 1.55 | 1.54 | 1.57 | 1.57 | 0.01 | 0.729 |
| Pentadecanoic cid methyl ester, C15:0 | 0.827 a | 0.757 b | 0.816 a | 0.773 ab | 0.01 | 0.039 |
| Palmitic acid methyl ester, C16:0 | 12.94 ab | 13.30 a | 12.56 b | 12.79 b | 0.01 | 0.030 |
| Heptadecanoic acid methyl ester, C17:0 | 0.713 c | 0.755 b | 0.744 bc | 0.789 a | 0.009 | 0.006 |
| Stearic acid methyl ester, C18:0 | 2.66 b | 3.00 a | 2.78 b | 2.80 b | 0.04 | 0.005 |
| Arachidic acid methyl ester, C20:0 | 0.125 b | 0.182 ab | 0.179 ab | 0.221 a | 0.01 | 0.013 |
| Myristoleic acid methyl ester, C14:1 n-9 | 0.125 b | 0.159 a | 0.164 a | 0.161 a | 0.01 | 0.021 |
| Palmitoleic acid methyl ester, C16:1 n-7 | 1.52 c | 1.70 a | 1.66 ab | 1.63 b | 0.02 | 0.001 |
| Oleic acid methyl ester, C18:1 n-9 | 11.39 b | 11.39 b | 11.42 ab | 11.43 a | 0.007 | 0.004 |
| Conjugated Linoleic Acid (CLA), C18:2 n-3 | 0.074 c | 0.080 b | 0.085 a | 0.086 a | 0.002 | 0.001 |
| Linolenic acid methyl ester (LA), C18:2 n-6 | 12.21 b | 12.49 a | 12.60 a | 12.56 a | 0.05 | 0.030 |
| α-Linolenic acid methyl ester (ALA), C18:3 n-3 | 2.47 c | 2.50 b | 2.51 b | 2.55 a | 0.01 | 0.007 |
| Arachidonic acid methyl ester(ARA), C20:4 n-6 | 0.537 ab | 0.523 b | 0.534 b | 0.548 a | 0.003 | 0.015 |
| Eicosapentaenoic acid methyl ester(EPA), C20:5n-3 | 0.049 c | 0.068 b | 0.074 a | 0.073 a | 0.004 | 0.001 |
| Docosahexaenoic acid(DHA), C22:6n-3 | 0.061 b | 0.065 b | 0.070 a | 0.076 a | 0.004 | 0.001 |
| Degree of FAs saturation, g/100 g FA methyl ester | ||||||
| Saturated FAs | 72.27 | 71.72 | 71.62 | 71.70 | 0.22 | 0.282 |
| Unsaturated FAs | 27.73 b | 28.28 a | 28.38 a | 28.30 a | 0.08 | 0.007 |
| Monounsaturated FAs | 13.04 c | 13.28 a | 13.26 ab | 13.19 b | 0.03 | 0.001 |
| Polyunsaturated FAs | 14.69 b | 15.00 a | 15.12 a | 15.11 a | 0.06 | 0.014 |
| Unsaturated FAs / Saturated FAs | 0.400 | 0.402 | 0.405 | 0.401 | 0.001 | 0.552 |
| Variable | Treatment 1 | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| C | FME | HNME | LNME | |||
| Conception rate, % | 76.9 c (20/26) | 84.61 bc (22/26) | 92.3 b (24/26) | 96.15 a (25/26) | - | 0.004 |
| Parturition rate, % | 69.23 c (18/26) | 84.61 b (22/26) | 88.46 ab (23/26) | 92.3 a (24/26) | - | 0.003 |
| Litter size at birth | 6.33 bc | 5.95 c | 7.17 b | 7.86 a | 2.4 | 0.004 |
| No. live litter sizes | 5.16 c | 5.90 ab | 6.65 b | 7.34 a | 3.7 | 0.003 |
| No. dead litter sizes | 1.17 a | 0.05 b | 0.52 ab | 0.52 ab | 2.2 | 0.130 |
| Litter weight at birth, g | 298.36 c | 324.17 b | 340.65 b | 409.30 a | 50.08 | 0.005 |
| Litter size at weaning | 5.27 c | 5.92 bc | 6.64 b | 7.21 a | 0.63 | < 0.001 |
| Litter weight at weaning, g | 1699.1 c | 2376.2 b | 2796.8 ab | 3144.6 a | 174.46 | < 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Desoky, N.I.; Hashem, N.M.; Elkomy, A.G.; Abo-Elezz, Z.R. Improving Rabbit Doe Metabolism and Whole Reproductive Cycle Outcomes via Fatty Acid-Rich Moringa oleifera Leaf Extract Supplementation in Free and Nano-Encapsulated Forms. Animals 2022, 12, 764. https://doi.org/10.3390/ani12060764
El-Desoky NI, Hashem NM, Elkomy AG, Abo-Elezz ZR. Improving Rabbit Doe Metabolism and Whole Reproductive Cycle Outcomes via Fatty Acid-Rich Moringa oleifera Leaf Extract Supplementation in Free and Nano-Encapsulated Forms. Animals. 2022; 12(6):764. https://doi.org/10.3390/ani12060764
Chicago/Turabian StyleEl-Desoky, Nagwa I., Nesrein M. Hashem, Ahmed G. Elkomy, and Zahraa R. Abo-Elezz. 2022. "Improving Rabbit Doe Metabolism and Whole Reproductive Cycle Outcomes via Fatty Acid-Rich Moringa oleifera Leaf Extract Supplementation in Free and Nano-Encapsulated Forms" Animals 12, no. 6: 764. https://doi.org/10.3390/ani12060764