
Effects of Dietary Supplementation with Honeybee Pollen and Its Supercritical Fluid Extract on Immune Response and Fillet’s Quality of Farmed Gilthead Seabream (Sparus aurata)

 , ,
, ,  , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement and Experimental Diets
2.2. Fish Rearing and Sampling
2.3. Gene Expression and Quantitative Real Time Determination
2.4. Fish Sampling and Processing
2.5. Analyses of Physical Parameters
2.6. Analyses of Chemical Parameters
2.6.1. Moisture, Total Lipids, and Fatty Acid Composition of Fillets
2.6.2. Fatty Acid Composition of HBP and HBP_SFE
2.6.3. Fillet Oxidative Status
2.6.4. Antioxidant Capacity
2.7. Statistical Analysis
3. Results
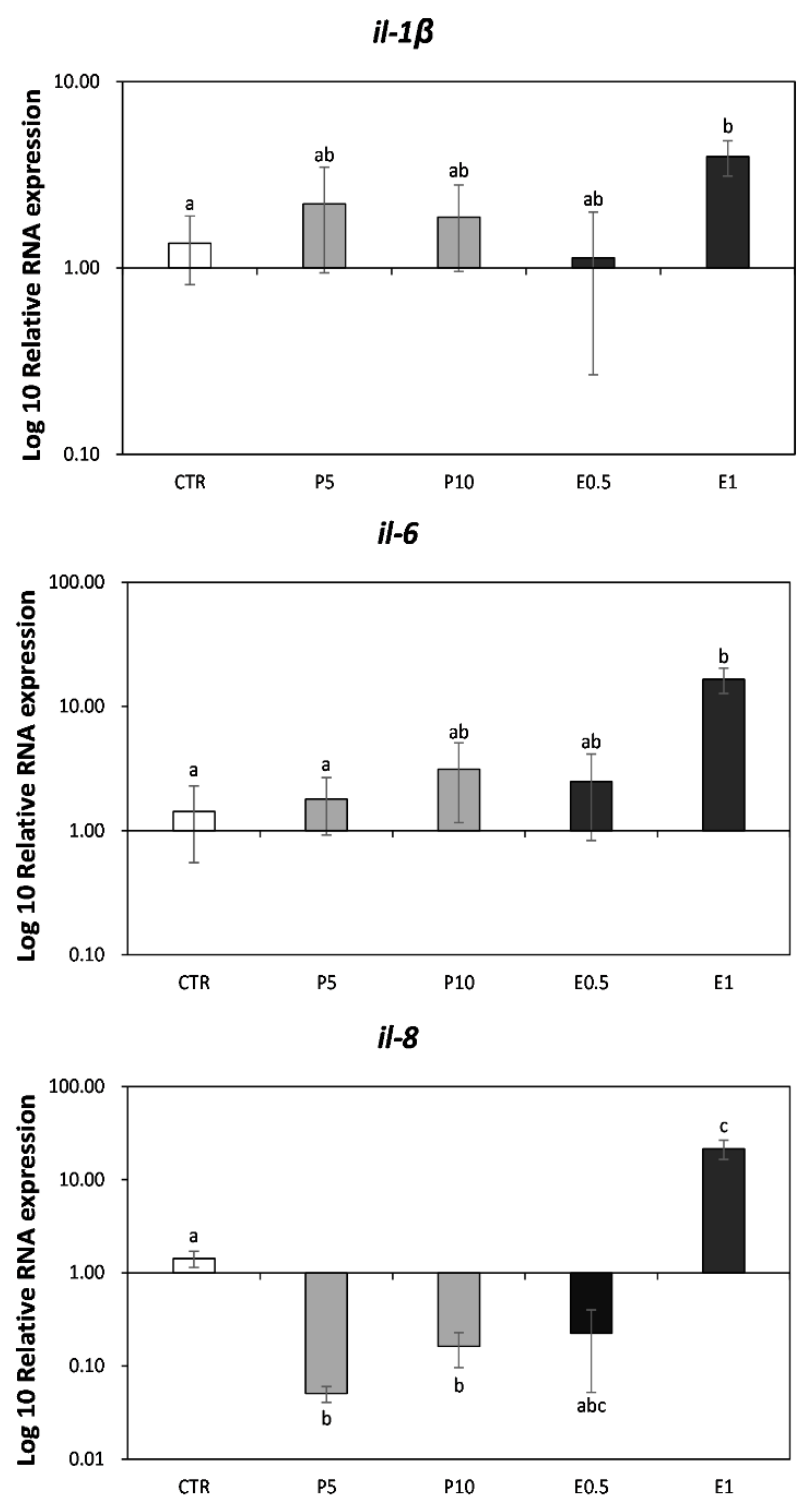
3.1. Gene Expression on Seabream Liver
3.2. Fillet Physical Parameters
3.3. Fillet Chemical Parameters
3.3.1. Chemical and Fatty Acid Composition
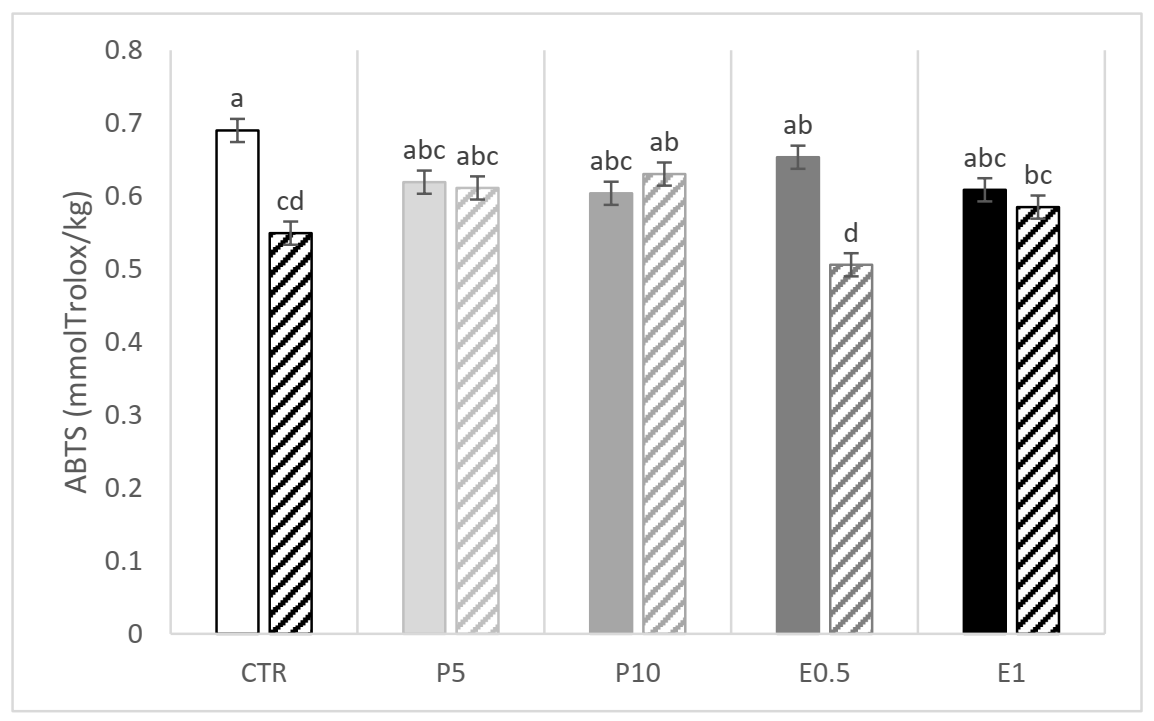
3.3.2. Fillet Oxidative Status
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Craig, S.; Kuhn, D.; Schwarz, M. Understanding Fish Nutrition, Feeds, and Feeding. Va. Coop. Extention 2017, 1–6. Available online: https://fisheries.tamu.edu/files/2019/01/FST-269.pdf (accessed on 24 December 2021).
- Hua, K.; Cobcroft, J.M.; Cole, A.; Condon, K.; Jerry, D.R.; Mangott, A.; Praeger, C.; Vucko, M.J.; Zeng, C.; Zenger, K.; et al. The Future of Aquatic Protein: Implications for Protein Sources in Aquaculture Diets. One Earth 2019, 1, 316–329. [Google Scholar] [CrossRef] [Green Version]
- Pulido-Rodriguez, L.F.; Cardinaletti, G.; Secci, G.; Randazzo, B.; Bruni, L.; Cerri, R.; Olivotto, I.; Tibaldi, E.; Parisi, G. Appetite Regulation, Growth Performances and Fish Quality Are Modulated by Alternative Dietary Protein Ingredients in Gilthead Sea Bream (Sparus aurata) Culture. Animals 2021, 11, 1919. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz-Cervantes, J.A.; Benavides-González, F.; Sánchez-Martínez, J.G.; Vázquez-Sauceda, M.d.L.L.; Ruiz-Uribe, A.J. Propolis in Aquaculture: A Review of Its Potential. Rev. Fish. Sci. Aquac. 2018, 26, 337–349. [Google Scholar] [CrossRef]
- Ushakova, N.A.; Pravdin, V.G.; Kravtsova, L.Z.; Ponomarev, S.V.; Gridina, T.S.; Ponomareva, E.N.; Rudoy, D.V.; Chikindas, M.L. Complex Bioactive Supplements for Aquaculture—Evolutionary Development of Probiotic Concepts. Probiotics Antimicrob. Proteins 2021, 13, 1696–1708. [Google Scholar] [CrossRef] [PubMed]
- Reverter, M.; Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Use of Plant Extracts in Fish Aquaculture as an Alternative to Chemotherapy: Current Status and Future Perspectives. Aquaculture 2014, 433, 50–61. [Google Scholar] [CrossRef]
- Bulfon, C.; Volpatti, D.; Galeotti, M. Current Research on the Use of Plant-Derived Products in Farmed Fish. Aquac. Res. 2015, 46, 513–551. [Google Scholar] [CrossRef]
- Messina, C.M.; Panettieri, V.; Arena, R.; Renda, G.; Espinosa Ruiz, C.; Morghese, M.; Piccolo, G.; Santulli, A.; Bovera, F. The Inclusion of a Supercritical Fluid Extract, Obtained From Honey Bee Pollen, in the Diet of Gilthead Sea Bream (Sparus aurata), Improves Fish Immune Response by Enhancing Anti-Oxidant, and Anti-Bacterial Activities. Front. Vet. Sci. 2020, 7, 95. [Google Scholar] [CrossRef] [Green Version]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.S. Impact of Plant Products on Innate and Adaptive Immune System of Cultured Finfish and Shellfish. Aquaculture 2011, 317, 1–15. [Google Scholar] [CrossRef]
- Santos, H.M.C.; Méndez, L.; Secci, G.; Parisi, G.; Martelli, R.; Medina, I. Pathway-Oriented Action of Dietary Essential Oils to Prevent Muscle Protein Oxidation and Texture Deterioration of Farmed Rainbow Trout. Animal 2019, 13, 2080–2091. [Google Scholar] [CrossRef]
- Hernández, A.; García García, B.; Jordán, M.J.; Hernández, M.D. Study of the Dose of Thyme Essential Oil in Feed to Prolong the Shelf Life of Gilthead Seabream (Sparus aurata). Aquac. Nutr. 2015, 21, 740–749. [Google Scholar] [CrossRef]
- Pascoal, A.; Rodrigues, S.; Teixeira, A.; Feás, X.; Estevinho, L.M. Biological Activities of Commercial Bee Pollens: Antimicrobial, Antimutagenic, Antioxidant and Anti-Inflammatory. Food Chem. Toxicol. 2014, 63, 233–239. [Google Scholar] [CrossRef]
- Ares, A.M.; Valverde, S.; Bernal, J.L.; Nozal, M.J.; Bernal, J. Extraction and Determination of Bioactive Compounds from Bee Pollen. J. Pharm. Biomed. Anal. 2018, 147, 110–124. [Google Scholar] [CrossRef]
- Xu, X.; Sun, L.; Dong, J.; Zhang, H. Breaking the Cells of Rape Bee Pollen and Consecutive Extraction of Functional Oil with Supercritical Carbon Dioxide. Innov. Food Sci. Emerg. Technol. 2009, 10, 42–46. [Google Scholar] [CrossRef]
- Panettieri, V.; Chatzifotis, S.; Messina, C.M.; Olivotto, I.; Manuguerra, S.; Randazzo, B.; Ariano, A.; Bovera, F.; Santulli, A.; Severino, L.; et al. Honey Bee Pollen in Meagre (Argyrosomus regius) Juvenile Diets: Effects on Growth, Diet Digestibility, Intestinal Traits, and Biochemical Markers Related to Health and Stress. Animals 2020, 10, 231. [Google Scholar] [CrossRef] [Green Version]
- Morais, M.; Moreira, L.; Feás, X.; Estevinho, L.M. Honeybee-Collected Pollen from Five Portuguese Natural Parks: Palynological Origin, Phenolic Content, Antioxidant Properties and Antimicrobial Activity. Food Chem. Toxicol. 2011, 49, 1096–1101. [Google Scholar] [CrossRef] [Green Version]
- Mistry, J.; Kennedy, J.F. Antioxidants in Food: Practical Applications. Carbohydr. Polym. 2003, 52, 88. [Google Scholar] [CrossRef]
- di Chiacchio, I.M.; Paiva, I.M.; de Abreu, D.J.M.; Carvalho, E.E.N.; Martínez, P.J.; Carvalho, S.M.; Mulero, V.; Murgas, L.D.S. Bee Pollen as a Dietary Supplement for Fish: Effect on the Reproductive Performance of Zebrafish and the Immunological Response of Their Offspring. Fish Shellfish. Immunol. 2021, 119, 300–307. [Google Scholar] [CrossRef]
- Sánchez, E.G.T.; Fuenmayor, C.A.; Vásquez Mejía, S.M.; Díaz-Moreno, C.; Suárez Mahecha, H. Effect of Bee Pollen Extract as a Source of Natural Carotenoids on the Growth Performance and Pigmentation of Rainbow Trout (Oncorhynchus mykiss). Aquaculture 2020, 514, 734490. [Google Scholar] [CrossRef]
- Attia, Y.A.; Al-Hanoun, A.; Tag El-Din, A.E.; Bovera, F.; Shewika, Y.E. Effect of Bee Pollen Levels on Productive, Reproductive and Blood Traits of NZW Rabbits. J. Anim. Physiol. Anim. Nutr. 2011, 95, 294–303. [Google Scholar] [CrossRef]
- Abbass, A.A.; El-Asely, A.M.; Kandiel, M.M.M. Effects of Dietary Propolis and Pollen on Growth Performance, Fecundity and Some Hematological Parameters of Oreochromis niloticus. Turk. J. Fish. Aquat. Sci. 2012, 12, 917–924. [Google Scholar] [CrossRef]
- El-Asely, A.M.; Abbass, A.A.; Austin, B. Honey Bee Pollen Improves Growth, Immunity and Protection of Nile Tilapia (Oreochromis niloticus) against Infection with Aeromonas hydrophila. Fish Shellfish. Immunol. 2014, 40, 500–506. [Google Scholar] [CrossRef]
- Ren, H.; Huang, Y.; Lin, L. Effects of Dietary Supplementation with Peony Pollen on Growth, Intestinal Function, Fillet Quality and Fatty Acids Profiles of Common Carp. Aquac. Nutr. 2021, 27, 908–917. [Google Scholar] [CrossRef]
- Cuesta, A.; Rodríguez, A.; Esteban, M.Á.; Meseguer, J. In Vivo Effects of Propolis, a Honeybee Product, on Gilthead Seabream Innate Immune Responses. Fish Shellfish. Immunol. 2005, 18, 71–80. [Google Scholar] [CrossRef]
- Prakatur, I.; Miskulin, M.; Pavic, M.; Marjanovic, K.; Blazicevic, V.; Miskulin, I.; Domacinovic, M. Intestinal Morphology in Broiler Chickens Supplemented with Propolis and Bee Pollen. Animals 2019, 9, 301. [Google Scholar] [CrossRef] [Green Version]
- Picoli, F.; Lopes, D.L.d.A.; Zampar, A.; Serafini, S.; Freccia, A.; Veronezi, L.O.; Kowalski, M.W.; Ghizzo, J.B.; Emerenciano, M.G.C. Dietary Bee Pollen Affects Hepatic–Intestinal Histomorphometry of Nile Tilapia Fingerlings. Aquac. Res. 2019, 50, 3295–3304. [Google Scholar] [CrossRef]
- Herrero, M.; Cifuentes, A.; Ibañez, E. Sub-and Supercritical Fluid Extraction of Functional Ingredients from Different Natural Sources: Plants, Food-by-Products, Algae and Microalgae: A Review. Food Chem. 2006, 98, 136–148. [Google Scholar] [CrossRef] [Green Version]
- Messina, C.M.; Troia, A.; Arena, R.; Manuguerra, S.; Ioannou, T.; Curcuraci, E.; Renda, G.; Hellio, C.; Santulli, A. Species-Specific Antioxidant Power and Bioactive Properties of the Extracts Obtained from Wild Mediterranean Calendula spp. (Asteraceae). Appl. Sci. 2019, 9, 4627. [Google Scholar] [CrossRef] [Green Version]
- Castro-Vargas, H.I.; Rodríguez-Varela, L.I.; Ferreira, S.R.S.; Parada-Alfonso, F. Extraction of Phenolic Fraction from Guava Seeds (Psidium guajava L.) Using Supercritical Carbon Dioxide and Co-Solvents. J. Supercrit. Fluids 2010, 51, 319–324. [Google Scholar] [CrossRef]
- Gómez-Requeni, P.; Mingarro, M.; Calduch-Giner, J.A.; Médale, F.; Martin, S.A.M.; Houlihan, D.F.; Kaushik, S.; Pérez-Sánchez, J. Protein Growth Performance, Amino Acid Utilisation and Somatotropic Axis Responsiveness to Fish Meal Replacement by Plant Protein Sources in Gilthead Sea Bream (Sparus aurata). Aquaculture 2004, 232, 493–510. [Google Scholar] [CrossRef]
- Peres, H.; Oliva-Teles, A. The Optimum Dietary Essential Amino Acid Profile for Gilthead Seabream (Sparus aurata) Juveniles. Aquaculture 2009, 296, 81–86. [Google Scholar] [CrossRef]
- Chomczynski, P. A Reagent for the Single-Step Simultaneous Isolation of RNA, DNA and Proteins from Cell and Tissue Samples. BioTechniques 1993, 15, 532–537. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- CIE. Colorimetry, 3rd ed.; Commission Internationale de l’Eclairage, CIE Central Bureau: Vienna, Austria, 2004. [Google Scholar]
- Eide, O.; Børresen, T.; Strøm, T. Minced Fish Production from Capelin (Mallotus villosus). A New Method for Gutting, Skinning and Removal of Fat from Small Fatty Fish Species. J. Food Sci. 1982, 47, 347–349. [Google Scholar] [CrossRef]
- Hultmann, L.; Rustad, T. Textural Changes during Iced Storage of Salmon (Salmo salar) and Cod (Gadus morhua). J. Aquat. Food Prod. Technol. 2002, 11, 105–123. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 19th ed.; Latimer, G.W., Jr., Ed.; AOAC: Washington, DC, USA, 2012. [Google Scholar]
- Folch, J.; Lees, M.; Sloane, G.H. A Simple Method for the Isolation and Purification of Total Lipids from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Morrison, W.R.; Smith, L.M. Preparation of Fatty Acid Methyl Esters and Dimethylacetals From Lipids. J. Lipid Res. 1964, 5, 600–608. [Google Scholar] [CrossRef]
- Secci, G.; Mancini, S.; Iaconisi, V.; Gasco, L.; Basto, A.; Parisi, G. Can the Inclusion of Black Soldier Fly (Hermetia illucens) in Diet Affect the Flesh Quality/Nutritional Traits of Rainbow Trout (Oncorhynchus mykiss) after Freezing and Cooking? Int. J. Food Sci. Nutr. 2019, 70, 161–171. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved Recovery of Fatty Acid through Direct Transesterification without Prior Extraction or Purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef]
- Messina, C.M.; Renda, G.; La Barbera, L.; Santulli, A. By-Products of Farmed European Sea Bass (Dicentrarchus labrax L.) as a Potential Source of n-3 PUFA. Biologia 2013, 68, 288–293. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Xiong, Y.L.; Decker, E.A. Inhibition of Protein and Lipid Oxidation in Beef Heart Surimi-like Material by Antioxidants and Combinations of pH, NaCl, and Buffer Type in the Washing Media. J. Agric. Food Chem. 1996, 44, 119–125. [Google Scholar] [CrossRef]
- Vyncke, W. Direct Determination of the Thiobarbituric Acid Value in Trichloracetic Acid Extracts of Fish as a Measure of Oxidative Rancidity. Fette Seifen Anstrichm. 1970, 72, 1084–1087. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decoloration Assay. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Mancini, S.; Preziuso, G.; Dal Bosco, A.; Roscini, V.; Szendro, Z.; Fratini, F.; Paci, G. Effect of Turmeric Powder (Curcuma longa L.) and Ascorbic Acid on Physical Characteristics and Oxidative Status of Fresh and Stored Rabbit Burgers. Meat Sci. 2015, 110, 93–100. [Google Scholar] [CrossRef]
- SAS. SAS/STAT 9.3 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2011. [Google Scholar]
- Abdelnour, S.A.; Abd El-Hack, M.E.; Alagawany, M.; Farag, M.R.; Elnesr, S.S. Beneficial Impacts of Bee Pollen in Animal Production, Reproduction and Health. J. Anim. Physiol. Anim. Nutr. 2019, 103, 477–484. [Google Scholar] [CrossRef]
- Liu, J.; Li, N.; Wang, C. Study on Extraction and Antioxidant Activity of Flavonoids from Peony Pollen. Food Res. Dev. 2012, 33, 39–44. [Google Scholar]
- Yuan, Y.; Wang, C.; Rao, H. Research Progress of Flavonoid in Peony Flowers. J. Qilu Univ. Technol. 2013, 4, 31–34. [Google Scholar]
- Lepetsos, P.; Papavassiliou, A.G. ROS/Oxidative Stress Signaling in Osteoarthritis. Biochim. Biophys. Acta-Mol. Basis Dis. 2016, 1862, 576–591. [Google Scholar] [CrossRef]
- Buonocore, F.; Forlenza, M.; Randelli, E.; Benedetti, S.; Bossù, P.; Meloni, S.; Secombes, C.J.; Mazzini, M.; Scapigliati, G. Biological Activity of Sea Bass (Dicentrarchus labrax L.) Recombinant Interleukin-1β. Mar. Biotechnol. 2005, 7, 609–617. [Google Scholar] [CrossRef]
- Hong, S.; Peddie, S.; Campos-Pérez, J.J.; Zou, J.; Secombes, C.J. The Effect of Intraperitoneally Administered Recombinant IL-1β on Immune Parameters and Resistance to Aeromonas salmonicida in the Rainbow Trout (Oncorhynchus mykiss). Dev. Comp. Immunol. 2003, 27, 801–812. [Google Scholar] [CrossRef]
- Secombes, C.J.; Wang, T.; Hong, S.; Peddie, S.; Crampe, M.; Laing, K.J.; Cunningham, C.; Zou, J. Cytokines and Innate Immunity of Fish. Dev. Comp. Immunol. 2001, 25, 713–723. [Google Scholar] [CrossRef]
- Fu, X.; Ding, Z.; Fan, J.; Wang, H.; Zhou, F.; Cui, L.; Boxiang, C.; Wang, W.; Liu, H. Characterization, Promoter Analysis and Expression of the Interleukin-6 Gene in Blunt Snout Bream, Megalobrama amblycephala. Fish Physiol. Biochem. 2016, 42, 1527–1540. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.; Zou, J.; Savan, R.; Kono, T.; Sakai, M.; Woo, J.; Secombes, C. Characterisation and Expression Analysis of an Interleukin 6 Homologue in the Japanese Pufferfish, Fugu rubripes. Dev. Comp. Immunol. 2005, 29, 775–789. [Google Scholar] [CrossRef]
- Nam, B.H.; Byon, J.Y.; Kim, Y.O.; Park, E.M.; Cho, Y.C.; Cheong, J.H. Molecular Cloning and Characterisation of the Flounder (Paralichthys olivaceus) Interleukin-6 Gene. Fish Shellfish. Immunol. 2007, 23, 231–236. [Google Scholar] [CrossRef]
- Castellana, B.; Iliev, D.B.; Sepulcre, M.P.; MacKenzie, S.; Goetz, F.W.; Mulero, V.; Planas, J.V. Molecular Characterization of Interleukin-6 in the Gilthead Seabream (Sparus aurata). Mol. Immunol. 2008, 45, 3363–3370. [Google Scholar] [CrossRef]
- Wang, T.T.; Song, X.H.; Bao, G.M.; Zhao, L.X.; Yu, X.; Zhao, J. Molecular Characterization, Expression Analysis, and Biological Effects of Interleukin-8 in Grass Carp Ctenopharyngodon idellus. Fish Shellfish. Immunol. 2013, 35, 1421–1432. [Google Scholar] [CrossRef]
- Muangnoi, C.; Chingsuwanrote, P.; Praengamthanachoti, P.; Svasti, S.; Tuntipopipat, S. Moringa oleifera Pod Inhibits Inflammatory Mediator Production by Lipopolysaccharide-Stimulated RAW 264.7 Murine Macrophage Cell Lines. Inflammation 2012, 35, 445–455. [Google Scholar] [CrossRef]
- Praengam, K.; Muangnoi, C.; Dawilai, S.; Awatchanawong, M.; Tuntipopipat, S. Digested Moringa oleifera Boiled Pod Exhibits Anti-Inflammatory Activity in CACO-2 Cells. J. Herbs Spices Med. Plants 2015, 21, 148–160. [Google Scholar] [CrossRef]
- Christybapita, D.; Divyagnaneswari, M.; Dinakaran Michael, R. Oral Administration of Eclipta alba Leaf Aqueous Extract Enhances the Non-Specific Immune Responses and Disease Resistance of Oreochromis mossambicus. Fish Shellfish. Immunol. 2007, 23, 840–852. [Google Scholar] [CrossRef]
- Chu, W.H. Adjuvant Effect of Propolis on Immunisation by Inactivated Aeromonas hydrophila in Carp (Carassius auratus Gibelio). Fish Shellfish. Immunol. 2006, 21, 113–117. [Google Scholar] [CrossRef]
- Mansour, A.T.; Miao, L.; Espinosa, C.; García-Beltrán, J.M.; Ceballos Francisco, D.C.; Esteban, M.Á. Effects of Dietary Inclusion of Moringa oleifera Leaves on Growth and Some Systemic and Mucosal Immune Parameters of Seabream. Fish Physiol. Biochem. 2018, 44, 1223–1240. [Google Scholar] [CrossRef]
- Yin, G.; Ardó, L.; Thompson, K.D.; Adams, A.; Jeney, Z.; Jeney, G. Chinese Herbs (Astragalus radix and Ganoderma lucidum) Enhance Immune Response of Carp, Cyprinus carpio, and Protection against Aeromonas hydrophila. Fish Shellfish. Immunol. 2009, 26, 140–145. [Google Scholar] [CrossRef]
- Zhang, G.; Gong, S.; Yu, D.; Yuan, H. Propolis and Herba epimedii Extracts Enhance the Non-Specific Immune Response and Disease Resistance of Chinese Sucker, Myxocyprinus asiaticus. Fish Shellfish. Immunol. 2009, 26, 467–472. [Google Scholar] [CrossRef]
- Bruni, L.; Randazzo, B.; Cardinaletti, G.; Zarantoniello, M.; Mina, F.; Secci, G.; Tulli, F.; Olivotto, I.; Parisi, G. Dietary Inclusion of Full-Fat Hermetia illucens Prepupae Meal in Practical Diets for Rainbow Trout (Oncorhynchus mykiss): Lipid Metabolism and Fillet Quality Investigations. Aquaculture 2020, 529, 735678. [Google Scholar] [CrossRef]
- Ha, N.C.; Hiep, L.T.B.; van Toan, L.; Nho, L.T.H.; Tuyet, N.T.N.; le Ngoc, N.T. Ergothioneine Accumulated in the Catfish Muscle by Supplementation of Flammulina velutipes Extract Prevents Lipid Oxidation, Improves Color Stability, and Limits Soft Structure of the Fillet during Frozen Storage. J. Food Processing Preserv. 2021, 45, e15340. [Google Scholar] [CrossRef]
- Ünal Şengör, G.F.; Balaban, M.O.; Topaloğlu, B.; Ayvaz, Z.; Ceylan, Z.; Doğruyol, H. Color Assessment by Different Techniques of Gilthead Seabream (Sparus aurata) during Cold Storage. Food Sci. Technol. 2018, 39, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, A.; Fontanillas, R.; Hernández-Contreras, A.; Hernández, M.D. Partial Replacement of Fish Oil with Vegetal Oils in Commercial Diets: The Effect on the Quality of Gilthead Seabream (Sparus aurata). Anim. Feed. Sci. Technol. 2020, 265, 114504. [Google Scholar] [CrossRef]
- Ayala, M.D.; Abdel, I.; Santaella, M.; Martínez, C.; Periago, M.J.; Gil, F.; Blanco, A.; Albors, O.L. Muscle Tissue Structural Changes and Texture Development in Sea Bream, Sparus aurata L., during Post-Mortem Storage. LWT-Food Sci. Technol. 2010, 43, 465–475. [Google Scholar] [CrossRef]
- Hematyar, N.; Mraz, J.; Stejskal, V.; Sampels, S.; Linhartová, Z.; Prokesova, M.; Vacha, F.; Krizek, M.; Dadakova, E.; Søndergård Møller, H.; et al. Comparison of Quality Changes in Eurasian Perch (Perca fluviatilis L.) Fillets Originated from Two Different Rearing Systems during Frozen and Refrigerated Storage. Foods 2021, 10, 1405. [Google Scholar] [CrossRef]
- Burgaard, M.G.; Jørgensen, B.M. Effect of Temperature on Quality-Related Changes in Cod (Gadus morhua) during Short- and Long-Term Frozen Storage. J. Aquat. Food Prod. Technol. 2010, 19, 249–263. [Google Scholar] [CrossRef]
- Makri, M. Biochemical and Textural Properties of Frozen Stored −22 °C Gilthead Seabream Sparus aurata Fillets. Afr. J. Biotechnol. 2009, 8, 1287–1299. [Google Scholar] [CrossRef]
- Sáez, M.I.; Suárez, M.D.; Cárdenas, S.; Martínez, T.F. Freezing and Freezing-Thawing Cycles on Textural and Biochemical Changes of Meagre (Argyrosomus regius, L.) Fillets during Further Cold Storage. Int. J. Food Prop. 2015, 18, 1635–1647. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.P.; Reddy, G.V.S.; Dhanapal, K.; Babu, P.H. Effect of Frozen Storage on the Quality and Shelflife of Mrigal (Cirrhinus mrigala). J. Entomol. Zool. Stud. 2021, 9, 1071–1076. [Google Scholar]
- Caballero, M.J.; Betancor, M.; Escrig, J.C.; Montero, D.; Espinosa de los Monteros, A.; Castro, P.; Ginés, R.; Izquierdo, M. Post Mortem Changes Produced in the Muscle of Sea Bream (Sparus aurata) during Ice Storage. Aquaculture 2009, 291, 210–216. [Google Scholar] [CrossRef]
- Cai, L.; Nian, L.; Zhao, G.; Zhang, Y.; Sha, L.; Li, J. Effect of Herring Antifreeze Protein Combined with Chitosan Magnetic Nanoparticles on Quality Attributes in Red Sea Bream (Pagrosomus major). Food Bioprocess Technol. 2019, 12, 409–421. [Google Scholar] [CrossRef]
- Nian, L.; Cao, A.; Cai, L.; Ji, H.; Liu, S. Effect of Vacuum Impregnation of Red Sea Bream (Pagrosomus major) with Herring AFP Combined with CS@Fe3O4 Nanoparticles during Freeze-Thaw Cycles. Food Chem. 2019, 291, 139–148. [Google Scholar] [CrossRef]
- Sathivel, S. Chitosan and Protein Coatings Affect Yield, Moisture Loss, and Lipid Oxidation of Pink Salmon (Oncorhynchus gorbuscha) Fillets During Frozen Storage. J. Food Sci. 2005, 70, e455–e459. [Google Scholar] [CrossRef]
- Gandotra, R. Change In Proximate Composition And Microbial Count By Low Temperaturepreservation In Fish Muscle Of Labeo rohita (HamBuch). IOSR J. Pharm. Biol. Sci. 2012, 2, 13–17. [Google Scholar] [CrossRef]
- Arannilewa, S.T.; Salawu, S.O.; Sorungbe, A.A.; Ola-Salawu, B.B. Effect of Frozen Period on the Chemical, Microbiological and Sensory Quality of Frozen Tilapia Fish (Sarotherodun galiaenus). Afr. J. Biotechnol. 2005, 4, 852–855. [Google Scholar] [CrossRef]
- Hassanin, S.I.A.; El-Daly, E.-S.A. Effect of Propolis and Garlic on Nile Tilapia Oreochromis niloticus Fillets during Frozen Storage. J. Arab. Aquac. Soc. 2013, 8, 237–248. [Google Scholar]
- Obemeata, O.; Christopher, N. Organoleptic Assessment and Proximate Analysis of Stored Tilapia guineensis. Annu. Rev. Res. Biol. 2012, 2, 46–52. [Google Scholar]
- Gill, I.; Valivety, R. Polyunsaturated Fatty Acids, Part 1: Occurrence, Biological Activities and Applications. Trends Biotechnol. 1997, 15, 401–409. [Google Scholar] [CrossRef]
- Dave, D.; Routray, W. Current Scenario of Canadian Fishery and Corresponding Underutilized Species and Fishery Byproducts: A Potential Source of Omega-3 Fatty Acids. J. Clean. Prod. 2018, 180, 617–641. [Google Scholar] [CrossRef]
- Bordoni, A.; Di Nunzio, M.; Danesi, F.; Biagi, P.L. Polyunsaturated Fatty Acids: From Diet to Binding to Ppars and Other Nuclear Receptors. Genes Nutr. 2006, 1, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, M.; Montero, D.; Torrecillas, S.; Castro, P.; Zamorano, M.J.; Izquierdo, M. Hepatic Biochemical, Morphological and Molecular Effects of Feeding Microalgae and Poultry Oils to Gilthead Sea Bream (Sparus aurata). Aquaculture 2021, 532, 736073. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CTR | P5 | P10 | E0.5 | E1 | |
|---|---|---|---|---|---|
| Dry matter | 88.94 | 88.19 | 87.63 | 88.43 | 88.62 |
| Ash | 5.50 | 4.99 | 4.35 | 4.71 | 4.51 |
| Crude protein | 39.78 | 39.92 | 38.26 | 38.91 | 39.46 |
| Ether extract | 17.85 | 17.25 | 17.15 | 17.75 | 17.15 |
| Crude fiber | 7.17 | 7.54 | 8.63 | 7.37 | 7.62 |
| Gene | Accession Number | F/R Primer Sequence (5′–3′) |
|---|---|---|
| il1-β | AJ277166 | F-GGGCTGAACAACAGCACTCTC |
| R-TTAACACTCTCCACCCTCCA | ||
| il-6 | AM749958 | F-AGGCAGGAGTTTGAAGCTGA |
| R-ATGCTGAAGTTGGTGGAAGG | ||
| il-8 | AM765841 | F-GCCACTCTGAAGAGGACAGG |
| R-TTTGGTTGTCTTTGGTCGAA | ||
| ef1α | AF184170 | F-CTTCAACGCTCAGGTCATCAT |
| R-GCACAGCGAAACGACCAAGGGGA | ||
| 18S | AM490061 | F-CTTCAACGCTCAGGTCATCAT |
| R-AGTTGGCACCGTTTATGGTC |
| Treatment, T | Storage, S | p-Value | RMSE | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CTR | P5 | P10 | E0.5 | E1 | 0 | 110 | T | S | ||
| Weight, g | 74.91 | 72.45 | 67.42 | 67.12 | 71.56 | 71.70 | 69.68 | 0.835 | 0.688 | 19.347 |
| L* | 53.11 | 54.10 | 53.87 | 53.88 | 54.47 | 48.65 | 59.13 | 0.725 | <0.0001 | 2.391 |
| a* | −1.75 | −1.73 | −1.51 | −1.89 | −1.84 | −0.48 | −3.01 | 0.790 | <0.0001 | 0.764 |
| b* | 0.20 | 0.78 | −0.06 | −0.29 | −0.66 | −0.53 | 0.52 | 0.172 | 0.0072 | 1.450 |
| pH | 6.06 | 6.06 | 6.14 | 6.11 | 6.06 | 6.04 | 6.14 | 0.163 | 0.0003 | 0.098 |
| Shear force, N | 43.12 | 41.33 | 49.18 | 40.57 | 44.12 | 40.94 | 46.39 | 0.360 | 0.064 | 11.127 |
| WHC, % | 73.31 | 77.93 | 76.17 | 78.43 | 78.48 | 86.87 | 66.85 | 0.608 | <0.0001 | 9.165 |
| Treatment, T | Storage, S | p-Value | RMSE | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CTR | P5 | P10 | E0.5 | E1 | 0 | 110 | T | S | ||
| Moisture | 61.35 | 60.17 | 63.05 | 62.61 | 61.27 | 67.58 | 55.81 | 0.788 | <0.0001 | 6.100 |
| Total lipids | 9.31 | 9.79 | 8.75 | 8.81 | 9.16 | 10.11 | 8.22 | 0.841 | 0.003 | 2.26 |
| Fatty acids | ||||||||||
| C14:0 | 2.18 | 2.19 | 2.18 | 2.21 | 2.16 | 2.10 | 2.27 | 0.878 | <0.0001 | 0.11 |
| C16:0 | 12.43 | 12.48 | 12.25 | 12.52 | 12.35 | 12.07 | 12.74 | 0.461 | <0.0001 | 0.41 |
| C16:1n-7 | 3.56 | 3.53 | 3.53 | 3.60 | 3.49 | 3.46 | 3.62 | 0.587 | 0.0003 | 0.16 |
| C18:0 | 3.10 | 3.10 | 3.15 | 3.03 | 3.15 | 3.09 | 3.12 | 0.245 | 0.422 | 0.15 |
| C18:1n-9 | 32.41 | 32.14 | 32.34 | 32.56 | 32.10 | 32.35 | 32.28 | 0.144 | 0.604 | 0.50 |
| C18:1n-7 | 2.69 | 2.69 | 2.66 | 2.71 | 2.68 | 2.69 | 2.68 | 0.212 | 0.535 | 0.06 |
| C18:2n-6 | 22.01 | 22.02 | 22.17 | 21.91 | 22.07 | 22.10 | 21.98 | 0.872 | 0.007 | 0.01 |
| C18:3n-3 | 3.86 ab | 3.90 ab | 3.84 b | 3.85 b | 3.99 a | 3.93 | 3.84 | 0.026 | 0.008 | 0.12 |
| C20:1n-9 | 1.55 ab | 1.62 a | 1.47 b | 1.53 ab | 1.61 a | 1.59 | 1.52 | 0.001 | 0.004 | 0.08 |
| C20:5n-3 | 2.39 | 2.35 | 2.43 | 2.38 | 2.47 | 2.44 | 2.36 | 0.185 | 0.183 | 0.13 |
| C22:5n-3 | 1.23 | 1.24 | 1.27 | 1.20 | 1.23 | 1.29 | 1.18 | 0.373 | <0.0001 | 0.08 |
| C22:6n-3 | 5.58 | 5.42 | 5.70 | 5.43 | 5.59 | 5.64 | 5.45 | 0.574 | 0.104 | 0.46 |
| Σ SFA | 18.56 | 18.64 | 18.46 | 18.59 | 18.53 | 18.13 | 18.98 | 0.869 | <0.0001 | 0.47 |
| Σ MUFA | 42.42 | 42.36 | 42.13 | 42.66 | 42.21 | 42.37 | 42.34 | 0.396 | 0.891 | 0.67 |
| Σ n-6PUFA | 23.86 | 23.93 | 24.12 | 23.81 | 23.86 | 24.08 | 23.75 | 0.722 | 0.405 | 0.58 |
| Σ n-3PUFA | 14.41 | 14.29 | 14.54 | 14.20 | 14.64 | 14.68 | 14.16 | 0.399 | 0.002 | 0.61 |
| Treatment, T | Storage, S | p-Value | RMSE | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CTR | P5 | P10 | E0.5 | E1 | 0 | 110 | T | S | ||
| CD | 0.262 | 0.274 | 0.263 | 0.237 | 0.230 | 0.243 | 0.263 | 0.628 | 0.322 | 0.079 |
| TBARS | 0.863 | 0.979 | 0.762 | 0.913 | 0.858 | 0.627 | 1.123 | 0.409 | <0.0001 | 0.271 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arena, R.; de Medeiros, A.C.L.; Secci, G.; Mancini, S.; Manuguerra, S.; Bovera, F.; Santulli, A.; Parisi, G.; Messina, C.M.; Piccolo, G. Effects of Dietary Supplementation with Honeybee Pollen and Its Supercritical Fluid Extract on Immune Response and Fillet’s Quality of Farmed Gilthead Seabream (Sparus aurata). Animals 2022, 12, 675. https://doi.org/10.3390/ani12060675
Arena R, de Medeiros ACL, Secci G, Mancini S, Manuguerra S, Bovera F, Santulli A, Parisi G, Messina CM, Piccolo G. Effects of Dietary Supplementation with Honeybee Pollen and Its Supercritical Fluid Extract on Immune Response and Fillet’s Quality of Farmed Gilthead Seabream (Sparus aurata). Animals. 2022; 12(6):675. https://doi.org/10.3390/ani12060675
Chicago/Turabian StyleArena, Rosaria, Adja Cristina Lira de Medeiros, Giulia Secci, Simone Mancini, Simona Manuguerra, Fulvia Bovera, Andrea Santulli, Giuliana Parisi, Concetta Maria Messina, and Giovanni Piccolo. 2022. "Effects of Dietary Supplementation with Honeybee Pollen and Its Supercritical Fluid Extract on Immune Response and Fillet’s Quality of Farmed Gilthead Seabream (Sparus aurata)" Animals 12, no. 6: 675. https://doi.org/10.3390/ani12060675