
The Animal-Human Interface in Farm Animal Production: Animal Fear, Stress, Reproduction and Welfare


Abstract
:Simple Summary
Abstract
1. Introduction
2. Emotions Associated with the Responses of Animals to Humans
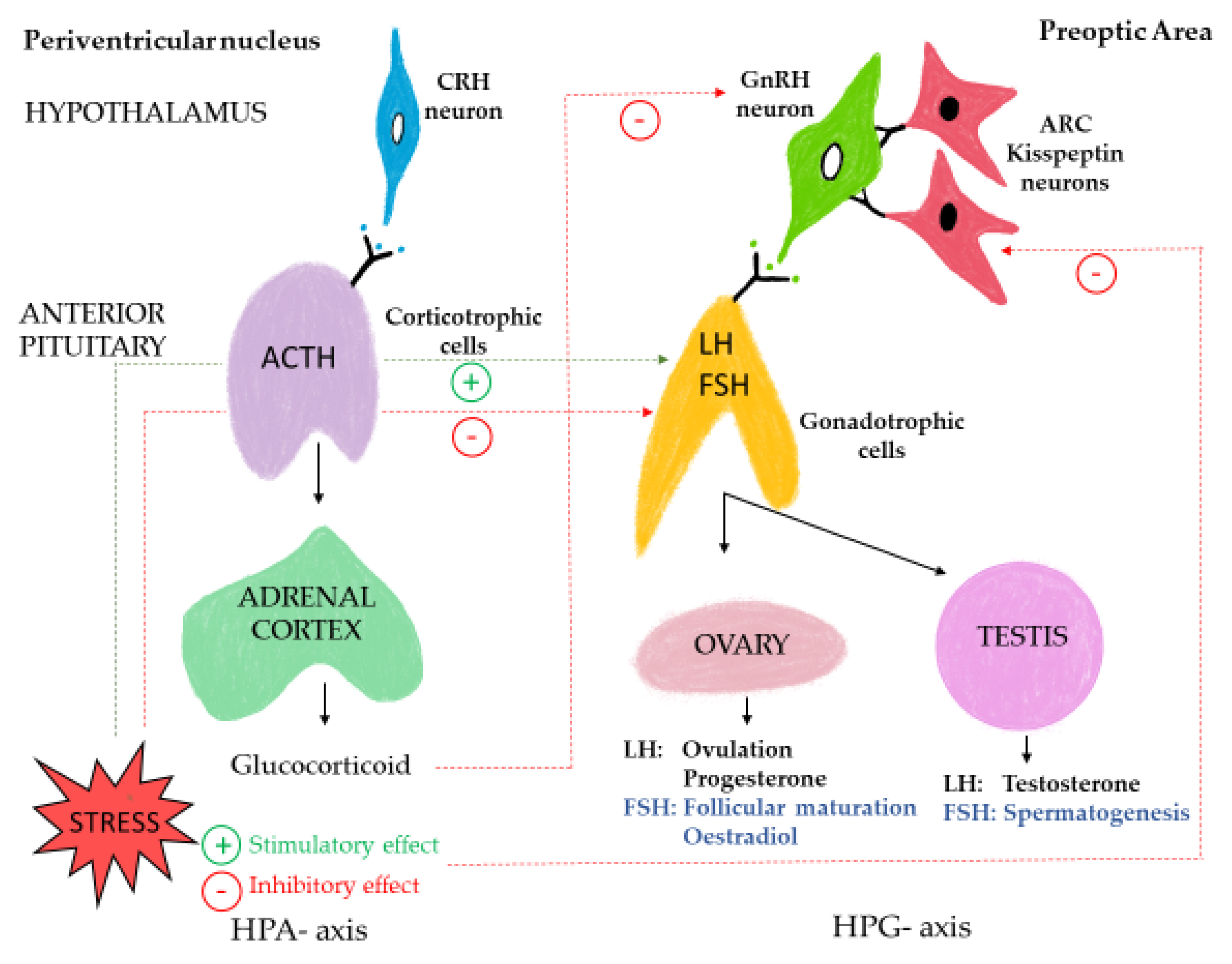
3. Fear, Stress, and Productivity: Interplay between HPA and HPG Axes
3.1. Evidence from Experimental Studies
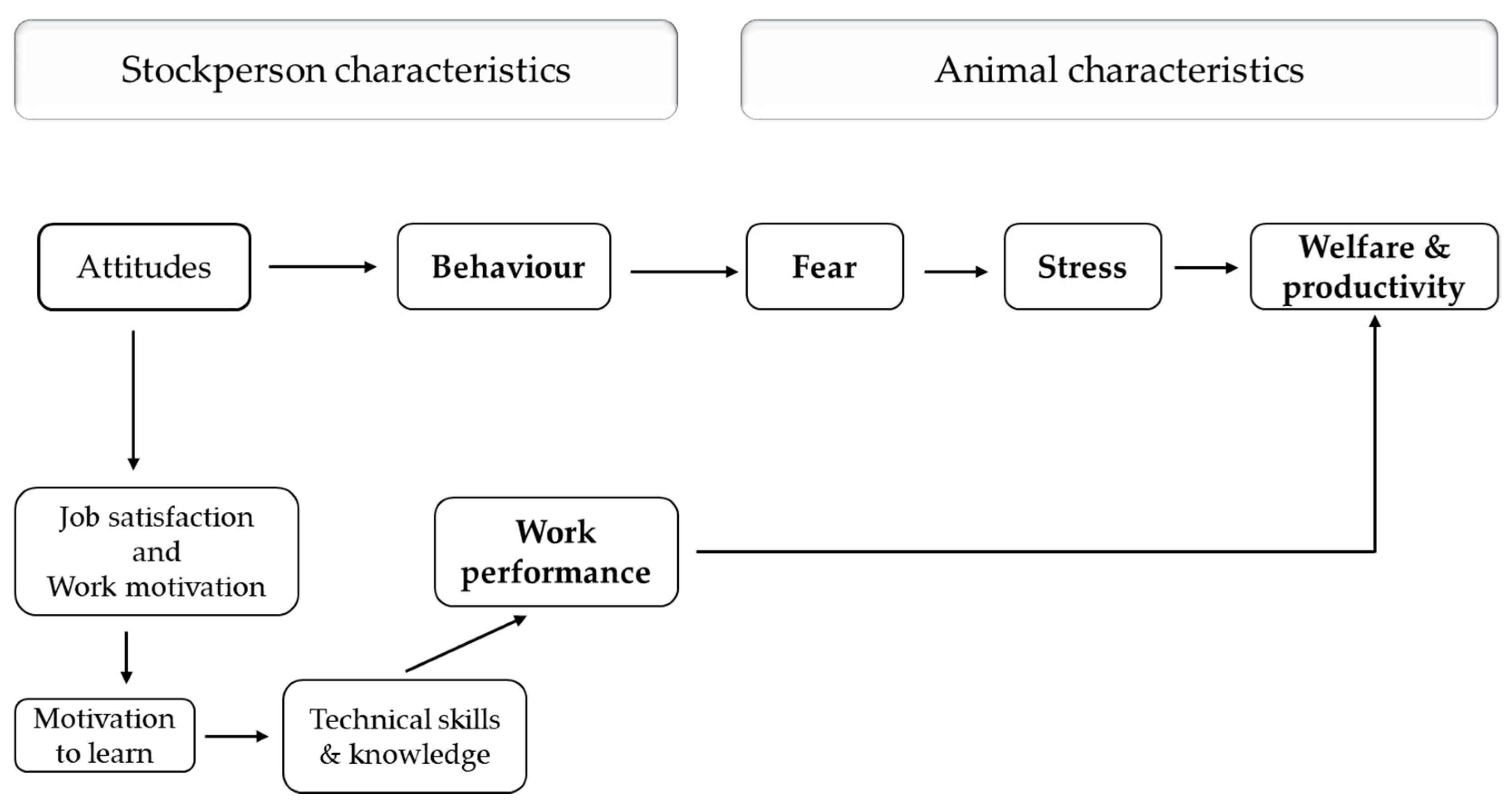
3.2. Evidence from On-Farm Studies

{kind=link}
{kind=link}
| Variables | Change Following Training (Relative to Control) | p Value |
|---|---|---|
| Stockperson attitudes | ||
| +ve Beliefs about ‘effort’ | 16% ↑ | 0.001 |
| +ve Beliefs about ‘petting’ | 25% ↑ | 0.01 |
| Stockperson behaviour | ||
| −ve behaviour | 50% ↓ | 0.001 |
| Cow behaviour Flight distance (m) | 7% ↓ | 0.05 |
| Cow physiology Milk cortisol (nM/L) | 32% ↓ | 0.06 |
| Cow productivity Milk yield (L/cow/month) | 5% ↑ | 0.02 |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fraser, D.G. Animal Welfare and the Intensification of Animal Production: An Alternative Interpretation; FAO, Ed.; FAO Readings in Ethics: Rome, Italy, 2005. [Google Scholar]
- Cronin, G.; Rault, J.; Glatz, P. Lessons Learned from Past Experience with Intensive Livestock Management Systems. Rev. Sci. Tech. 2014, 33, 139–151. [Google Scholar] [CrossRef]
- Tallet, C.; Brajon, S.; Devillers, N.; Lensink, J. Pig–human interactions: Creating a positive perception of humans to ensure pig welfare. In Advances in Pig Welfare; Elsevier: Amsterdam, The Netherlands, 2018; pp. 381–398. [Google Scholar]
- Boissy, A.; Fisher, A.; Bouix, J.; Hinch, G.; Le Neindre, P. Genetics of fear in ruminant livestock. Livest. Prod. Sci. 2005, 93, 23–32. [Google Scholar] [CrossRef]
- Hemsworth, P.; Coleman, G. Human-animal interactions and animal productivity and welfare. In Human-Livestock Interactions: The Stockperson and the Productivity and Welfare of Intensively Farmed Animals, 2nd ed.; CABI: Wallingford, England, 2011; pp. 47–83. [Google Scholar]
- Boivin, X.; Bensoussan, S.; L’Hotellier, N.; Bignon, L.; Brives, H.; Brule, A.; Godet, J.; Grannec, M.-L.; Hausberger, M.; Kling-Eveillard, F. Humans and livestock animals at work: Toward a multidisciplinary approach of relational practices. INRA Prod. Anim. 2012, 25, 159–167. [Google Scholar] [CrossRef]
- Hemsworth, P. Ethical stockmanship. Aust. Vet. J. 2007, 85, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Pinillos, R.G.; Appleby, M.C.; Manteca, X.; Scott-Park, F.; Smith, C.; Velarde, A. One welfare—A platform for improving human and animal welfare. Vet. Rec. 2016, 179, 412–413. [Google Scholar] [CrossRef] [Green Version]
- Blumberg, M.; Pringle, C.D. The missing opportunity in organizational research: Some implications for a theory of work performance. Acad. Manag. Rev. 1982, 7, 560–569. [Google Scholar] [CrossRef]
- Coleman, G.; Hemsworth, P. Training to improve stockperson beliefs and behaviour towards livestock enhances welfare and productivity. Rev. Sci. Tech. 2014, 33, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Désiré, L.; Boissy, A.; Veissier, I. Emotions in farm animals:: A new approach to animal welfare in applied ethology. Behav. Process. 2002, 60, 165–180. [Google Scholar] [CrossRef]
- Hemsworth, P.; Sherwen, S.; Coleman, G. Human Contact. In Animal Welfare, 3rd ed.; Appleby, M., Olsson, I., Galindo, F., Eds.; CAB International: Oxford, UK, 2018. [Google Scholar]
- Rushen, J.; de Passillé, A.M. The motivation of non-nutritive sucking in calves, Bos taurus. Anim. Behav. 1995, 49, 1503–1510. [Google Scholar] [CrossRef]
- Panksepp, J. Affective consciousness: Core emotional feelings in animals and humans. Conscious. Cogn. 2005, 14, 30–80. [Google Scholar] [PubMed]
- Panksepp, J. The basic emotional circuits of mammalian brains: Do animals have affective lives? Neurosci. Biobehav. Rev. 2011, 35, 1791–1804. [Google Scholar] [PubMed]
- Panksepp, J. The philosophical implications of affective neuroscience. J. Conscious. Stud. 2012, 19, 6–48. [Google Scholar]
- Alcaro, A.; Carta, S.; Panksepp, J. The affective core of the self: A neuro-archetypical perspective on the foundations of human (and animal) subjectivity. Front. Psychol. 2017, 8, 1424. [Google Scholar]
- Mellor, D. Animal Emotions, Behaviour and the Promotion of Positive Welfare States; Taylor & Francis: Abingdon, UK, 2012. [Google Scholar]
- Alcaro, A.; Panksepp, J. The SEEKING mind: Primal neuro-affective substrates for appetitive incentive states and their pathological dynamics in addictions and depression. Neurosci. Biobehav. Rev. 2011, 35, 1805–1820. [Google Scholar] [PubMed]
- Waiblinger, S.; Boivin, X.; Pedersen, V.; Tosi, M.-V.; Janczak, A.M.; Visser, E.K.; Jones, R.B. Assessing the human–animal relationship in farmed species: A critical review. Appl. Anim. Behav. Sci. 2006, 101, 185–242. [Google Scholar]
- Hemsworth, P. Key determinants of pig welfare: Implications of animal management and housing design on livestock welfare. Anim. Prod. Sci. 2018, 58, 1375–1386. [Google Scholar] [CrossRef]
- Rault, J.-L.; Waiblinger, S.; Boivin, X.; Hemsworth, P. The power of a positive human–animal relationship for animal welfare. Front. Vet. Sci. 2020, 7, 857. [Google Scholar]
- Sapolsky, R.M. Endocrinology of the stress-response. In Behavioral Endocrinology; Becker, J.B., Breedlove, S.M., Crews, D., McCarthy, M.M., Eds.; MIT Press: Cambridge, MA, USA, 2002; pp. 409–450. [Google Scholar]
- Moberg, G.P. Biological response to stress: Implications for animal welfare. Biol. Anim. Stress Basic Princ. Implic. Anim. Welf. 2000, 1, 21. [Google Scholar]
- Turner, A.I.; Keating, C.L.; Tilbrook, A.J. Sex differences and the role of sex steroids in sympatho-adrenal medullary system and hypothalamo-pituitary adrenal axis responses to stress. In Sex Steroids; IntechOpen: London, UK, 2012. [Google Scholar]
- Goldstein, D.S. Stress-induced activation of the sympathetic nervous system. Bailliere’s Clin. Endocrinol. Metab. 1987, 1, 253–278. [Google Scholar]
- Heller, H. Inactivation of vasopressin and oxytocin in vivo. In Polypeptides Which Affect Smooth Muscles and Blood Vessels; Pergamon Press: London, UK, 1960; p. 59. [Google Scholar]
- Minton, J.E.; Parsons, K. Adrenocorticotropic hormone and cortisol response to corticotropin-releasing factor and lysine vasopressin in pigs. J. Anim. Sci. 1993, 71, 724–729. [Google Scholar] [CrossRef] [Green Version]
- Howlett, T.A.; Rees, L.H. Endogenous opioid peptides and hypothalamo-pituitary function. Annu. Rev. Physiol. 1986, 48, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Owens, P.C.; Smith, R. Opioid peptides in blood and cerebrospinal fluid during acute stress. Bailliere’s Clin. Endocrinol. Metab. 1987, 1, 415–437. [Google Scholar]
- Torpy, D.J.; Chrousos, G.P. The three-way interactions between the hypothalamic-pituitary-adrenal and gonadal axes and the immune system. Bailliere’s Clin. Rheumatol. 1996, 10, 181–198. [Google Scholar]
- Turner, A.I.; Hemsworth, P.H.; Tilbrook, A. Susceptibility of reproduction in female pigs to impairment by stress and the role of the hypothalamo–pituitary–adrenal axis. Reprod. Fertil. Dev. 2002, 14, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, P.H.; Mellor, D.; Cronin, G.; Tilbrook, A. Scientific assessment of animal welfare. N. Z. Vet. J. 2015, 63, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, P.; Barnett, J.; Hansen, C. The influence of inconsistent handling by humans on the behaviour, growth and corticosteroids of young pigs. Appl. Anim. Behav. Sci. 1987, 17, 245–252. [Google Scholar] [CrossRef]
- Hemsworth, P.; Barnett, J.; Hansen, C. The influence of handling by humans on the behaviour, reproduction and corticosteroids of male and female pigs. Appl. Anim. Behav. Sci. 1986, 15, 303–314. [Google Scholar] [CrossRef]
- Barnett, J.; Hemsworth, P. The effects of individual and group housing on sexual behaviour and pregnancy in pigs. Anim. Reprod. Sci. 1991, 25, 265–273. [Google Scholar] [CrossRef]
- Coubrough, R. Stress and Fertility. A Review. Onderstepoort J. Vet. Res. 1985, 52, 153–156. [Google Scholar]
- Moberg, G.P. A model for assessing the impact of behavioral stress on domestic animals. J. Anim. Sci. 1987, 65, 1228–1235. [Google Scholar] [CrossRef] [Green Version]
- Clarke, I.; Hemsworth, P.; Barnett, J.; Tilbrook, A. Stress and reproduction in farm-animals. In Stress and Reproduction, Port Douglas Australia, July, 1991; Raven Press: Newark, DE, USA, 1992; pp. 239–252. [Google Scholar]
- Liptrap, R.M. Stress and reproduction in domestic animals. Ann. N. Y. Acad. Sci. 1993, 697, 275–284. [Google Scholar] [CrossRef]
- Tilbrook, A.J.; Turner, A.I.; Clarke, I.J. Effects of stress on reproduction in non-rodent mammals: The role of glucocorticoids and sex differences. Rev. Reprod. 2000, 5, 105–113. [Google Scholar] [CrossRef]
- Barnett, J.; Hemsworth, P.; Newman, E. Fear of humans and its relationships with productivity in laying hens at commercial farms. Br. Poult. Sci. 1992, 33, 699–710. [Google Scholar] [CrossRef]
- Toufexis, D.; Rivarola, M.A.; Lara, H.; Viau, V. Stress and the reproductive axis. J. Neuroendocrinol. 2014, 26, 573–586. [Google Scholar]
- Seeburg, P.H.; Mason, A.J.; Stewart, T.A.; Nikolics, K. The mammalian GnRH gene and its pivotal role in reproduction. In Proceedings of the The Annual Laurentian Hormone Conference, Quebec, QC, Canada, 24–26 August 1986; pp. 69–98. [Google Scholar]
- Clarke, I. Effector mechanisms of the hypothalamus that regulate the anterior pituitary gland. Auton.-Endocr. Interact. 1996, 1, 45–88. [Google Scholar]
- Clayton, R. Gonadotrophin-releasing hormone: Its actions and receptors. J. Endocrinol. 1989, 120, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Clarke, I. The hypothalamo-pituitary axis. In Scientific Essentials of Reproductive Medicine; W.B. Saunders Company Ltd.: London, UK, 1996; pp. 120–132. [Google Scholar]
- Esbenshade, K.L.; Ziecik, A.J.; Britt, J.H. Regulation and action of gonadotrophins in pigs. J. Reprod. Fertil. Suppl. 1990, 40, 19–32. [Google Scholar]
- Calisi, R.M.; Austin, S.H.; Lang, A.S.; MacManes, M.D. Sex-biased transcriptomic response of the reproductive axis to stress. Horm. Behav. 2018, 100, 56–68. [Google Scholar]
- Whirledge, S.; Cidlowski, J.A. Glucocorticoids and reproduction: Traffic control on the road to reproduction. Trends Endocrinol. Metab. 2017, 28, 399–415. [Google Scholar] [PubMed]
- Scott, C.J.; Rose, J.L.; Gunn, A.J.; McGrath, B.M. Kisspeptin and the regulation of the reproductive axis in domestic animals. J. Endocrinol. 2019, 240, R1–R16. [Google Scholar]
- Barnett, J.; Tauson, R.; Downing, J.; Janardhana, V.; Lowenthal, J.; Butler, K.; Cronin, G. The effects of a perch, dust bath, and nest box, either alone or in combination as used in furnished cages, on the welfare of laying hens. Poult. Sci. 2009, 88, 456–470. [Google Scholar]
- Barb, C.; Kraeling, R.; Rampacek, G.; Fonda, E.; Kiser, T. Inhibition of ovulation and LH secretion in the gilt after treatment with ACTH or hydrocortisone. Reproduction 1982, 64, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.I.; Hemsworth, P.H.; Canny, B.J.; Tilbrook, A.J. Sustained but Not Repeated Acute Elevation of Cortisol Impaired the Luteinizing Hormone Surge, Estrus, and Ovulation in Gilts1. Biol. Reprod. 1999, 61, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Gonyou, H.; Hemsworth, P.; Barnett, J. Effects of frequent interactions with humans on growing pigs. Appl. Anim. Behav. Sci. 1986, 16, 269–278. [Google Scholar] [CrossRef]
- Hemsworth, P.; Barnett, J. The effects of aversively handling pigs, either individually or in groups, on their behaviour, growth and corticosteroids. Appl. Anim. Behav. Sci. 1991, 30, 61–72. [Google Scholar] [CrossRef]
- Paterson, A.; Pearce, G. Boar-induced puberty in gilts handled pleasantly or unpleasantly during rearing. Appl. Anim. Behav. Sci. 1989, 22, 225–233. [Google Scholar] [CrossRef]
- Hemsworth, P.; Barnett, J.; Hansen, C. The influence of handling by humans on the behavior, growth, and corticosteroids in the juvenile female pig. Horm. Behav. 1981, 15, 396–403. [Google Scholar] [CrossRef]
- Kunavongkrit, A.; Madej, A.; Einarsson, S. Plasma levels of cortisol in zero-weaned and lactating sows during the first two weeks post partum. Domest. Anim. Endocrinol. 1984, 1, 217–223. [Google Scholar] [CrossRef]
- Turner, A.I.; Hemsworth, P.H.; Hughes, P.; Canny, B.; Tilbrook, A.J. The effect of repeated boar exposure on cortisol secretion and reproduction in gilts. Anim. Reprod. Sci. 1998, 51, 143–154. [Google Scholar] [CrossRef]
- Andersen, I.L.; Berg, S.; Bøe, K.E.; Edwards, S. Positive handling in late pregnancy and the consequences for maternal behaviour and production in sows. Appl. Anim. Behav. Sci. 2006, 99, 64–76. [Google Scholar] [CrossRef]
- Hayes, M.E.; Hemsworth, L.M.; Morrison, R.S.; Tilbrook, A.J.; Hemsworth, P.H. Positive Human Contact and Housing Systems Impact the Responses of Piglets to Various Stressors. Animals 2021, 11, 1619. [Google Scholar] [CrossRef] [PubMed]
- Rushen, J.; De Passille, A.; Munksgaard, L. Fear of people by cows and effects on milk yield, behavior, and heart rate at milking. J. Dairy Sci. 1999, 82, 720–727. [Google Scholar] [CrossRef]
- Breuer, K.; Hemsworth, P.H.; Barnett, J.L.; Matthews, L.R.; Coleman, G.J. Behavioural response to humans and the productivity of commercial dairy cows. Appl. Anim. Behav. Sci. 2000, 66, 273–288. [Google Scholar] [CrossRef]
- Breuer, K.; Hemsworth, P.H.; Coleman, G.J. The effect of positive or negative handling on the behavioural and physiological responses of nonlactating heifers. Appl. Anim. Behav. Sci. 2003, 84, 3–22. [Google Scholar] [CrossRef]
- Bruckmaier, R.M.; Blum, J.W. Oxytocin release and milk removal in ruminants. J. Dairy Sci. 1998, 81, 939–949. [Google Scholar]
- Neumann, I.D.; Torner, L.; Wigger, A. Brain oxytocin: Differential inhibition of neuroendocrine stress responses and anxiety-related behaviour in virgin, pregnant and lactating rats. Neuroscience 1999, 95, 567–575. [Google Scholar]
- Feldman, S.; Conforti, N.; Weidenfeld, J. Limbic pathways and hypothalamic neurotransmitters mediating adrenocortical responses to neural stimuli. Neurosci. Biobehav. Rev. 1995, 19, 235–240. [Google Scholar] [CrossRef]
- Hughes, B.; Black, A. The influence of handling on egg production, egg shell quality and avoidance behaviour of hens. Br. Poult. Sci. 1976, 17, 135–144. [Google Scholar] [CrossRef]
- Barnett, J.; Hemsworth, P.; Hennessy, D.; McCallum, T.; Newman, E. The effects of modifying the amount of human contact on behavioural, physiological and production responses of laying hens. Appl. Anim. Behav. Sci. 1994, 41, 87–100. [Google Scholar] [CrossRef]
- Skyes, J.F. Animal and Fowl. In Water: Yearbook of Agriculture, 1st ed.; The United States Department of Agriculture: Washington, DC, USA, 1955; pp. 14–18. [Google Scholar]
- Pedersen, L.; Studnitz, M.; Jensen, K.; Giersing, A. Suckling behaviour of piglets in relation to accessibility to the sow and the presence of foreign litters. Appl. Anim. Behav. Sci. 1998, 58, 267–279. [Google Scholar]
- Hemsworth, P.; Barnett, J.; Coleman, G.J.; Hansen, C. A study of the relationships between the attitudinal and behavioural profiles of stockpersons and the level of fear of humans and reproductive performance of commercial pigs. Appl. Anim. Behav. Sci. 1989, 23, 301–314. [Google Scholar] [CrossRef]
- Waiblinger, S.; Menke, C.; Coleman, G. The relationship between attitudes, personal characteristics and behaviour of stockpeople and subsequent behaviour and production of dairy cows. Appl. Anim. Behav. Sci. 2002, 79, 195–219. [Google Scholar] [CrossRef]
- Edwards, L.E.; Coleman, G.J.; Butler, K.L.; Hemsworth, P.H. The human-animal relationship in Australian caged laying hens. Animals 2019, 9, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemsworth, P.; Coleman, G.; Barnett, J.; Jones, R. Behavioural responses to humans and the productivity of commercial broiler chickens. Appl. Anim. Behav. Sci. 1994, 41, 101–114. [Google Scholar] [CrossRef]
- Cransberg, P.; Hemsworth, P.H.; Coleman, G.J. Human factors affecting the behaviour and productivity of commercial broiler chickens. Br. Poult. Sci. 2000, 41, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, P.H.; Pedersen, V.; Cox, M.; Cronin, G.M.; Coleman, G.J. A note on the relationship between the behavioural response of lactating sows to humans and the survival of their piglets. Appl. Anim. Behav. Sci. 1999, 65, 43–52. [Google Scholar] [CrossRef]
- Hemsworth, P.H.; Coleman, G.J.; Barnett, J.L.; Borg, S. Relationships between human-animal interactions and productivity of commercial dairy cows. J. Anim. Sci. 2000, 78, 2821–2831. [Google Scholar] [CrossRef]
- Hemsworth, P.; Coleman, G.; Barnett, J. Improving the attitude and behaviour of stockpersons towards pigs and the consequences on the behaviour and reproductive performance of commercial pigs. Appl. Anim. Behav. Sci. 1994, 39, 349–362. [Google Scholar] [CrossRef]
- Coleman, G.; Hemsworth, P.; Hay, M.; Cox, M. Modifying stockperson attitudes and behaviour towards pigs at a large commercial farm. Appl. Anim. Behav. Sci. 2000, 66, 11–20. [Google Scholar] [CrossRef]
- Coleman, G.J.; Hemsworth, P.H.; Hay, M. Predicting stockperson behaviour towards pigs from attitudinal and job-related variables and empathy. Appl. Anim. Behav. Sci. 1998, 58, 63–75. [Google Scholar] [CrossRef]
- Hemsworth, P.; Coleman, G. Human-Livestock Interactions: The Stockperson and the Productivity and Welfare of Farmed Animals; CAB International: Wallingford, UK, 2011. [Google Scholar]
- Hemsworth, P.; Coleman, G.; Barnett, J.; Borg, S.; Dowling, S. The effects of cognitive behavioral intervention on the attitude and behavior of stockpersons and the behavior and productivity of commercial dairy cows. J. Anim. Sci. 2002, 80, 68–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemsworth, P.H. The effects of stockperson education and training on farm animal welfare. In Proceedings of the International Conference of Pig Welfare, Copenhagen, Denmark, 29–30 April 2015. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acharya, R.Y.; Hemsworth, P.H.; Coleman, G.J.; Kinder, J.E. The Animal-Human Interface in Farm Animal Production: Animal Fear, Stress, Reproduction and Welfare. Animals 2022, 12, 487. https://doi.org/10.3390/ani12040487
Acharya RY, Hemsworth PH, Coleman GJ, Kinder JE. The Animal-Human Interface in Farm Animal Production: Animal Fear, Stress, Reproduction and Welfare. Animals. 2022; 12(4):487. https://doi.org/10.3390/ani12040487
Chicago/Turabian StyleAcharya, Rutu Y., Paul H. Hemsworth, Grahame J. Coleman, and James E. Kinder. 2022. "The Animal-Human Interface in Farm Animal Production: Animal Fear, Stress, Reproduction and Welfare" Animals 12, no. 4: 487. https://doi.org/10.3390/ani12040487