
Effects of Different Ammonia Concentrations on Pulmonary Microbial Flora, Lung Tissue Mucosal Morphology, Inflammatory Cytokines, and Neurotransmitters of Broilers

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Experimental Diet and Feeding Management
2.3. Sample Collection
2.4. Morphological Observation of Lung Tissue Mucosa and Determination of Serum Immune Cytokines and Neurotransmitters
2.5. DNA Extraction and PCR Amplification 16s rDNA
2.6. Illumina MiSeq Sequencing
2.7. Statistical Analysis
3. Results
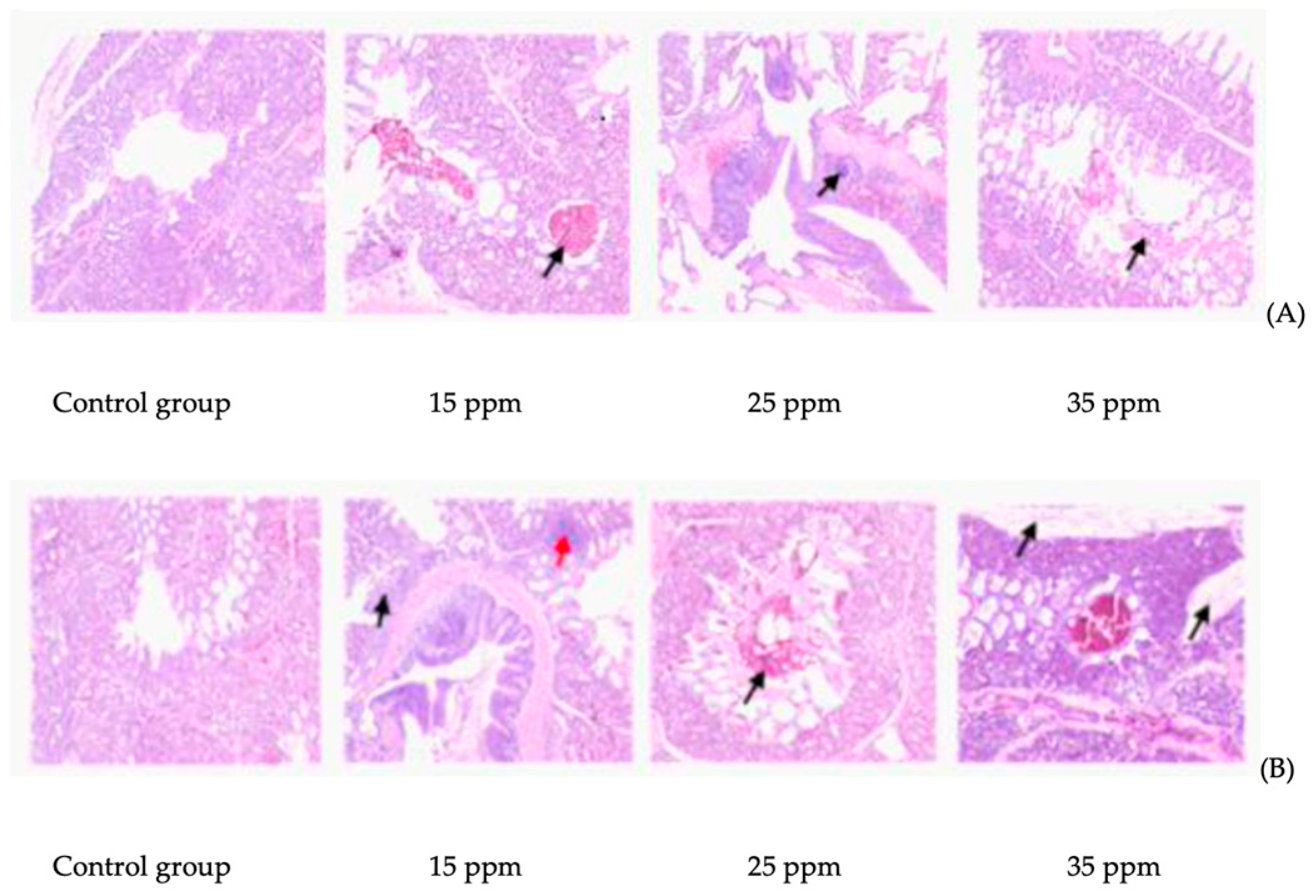
3.1. Effect of Ammonia on Lung Tissue Mucosa of Broilers
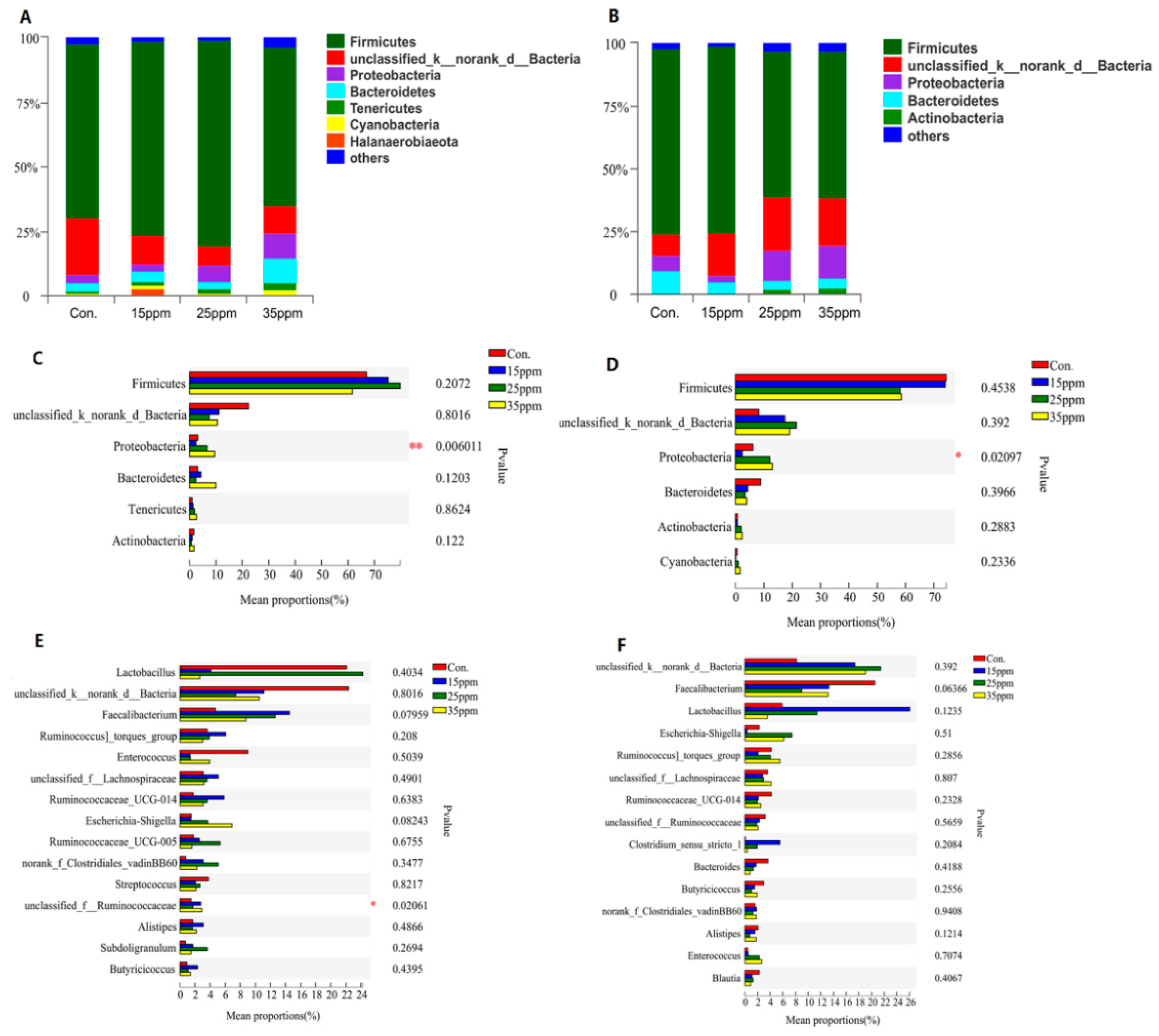
3.2. Effect of Ammonia on Pulmonary Microflora of Broilers
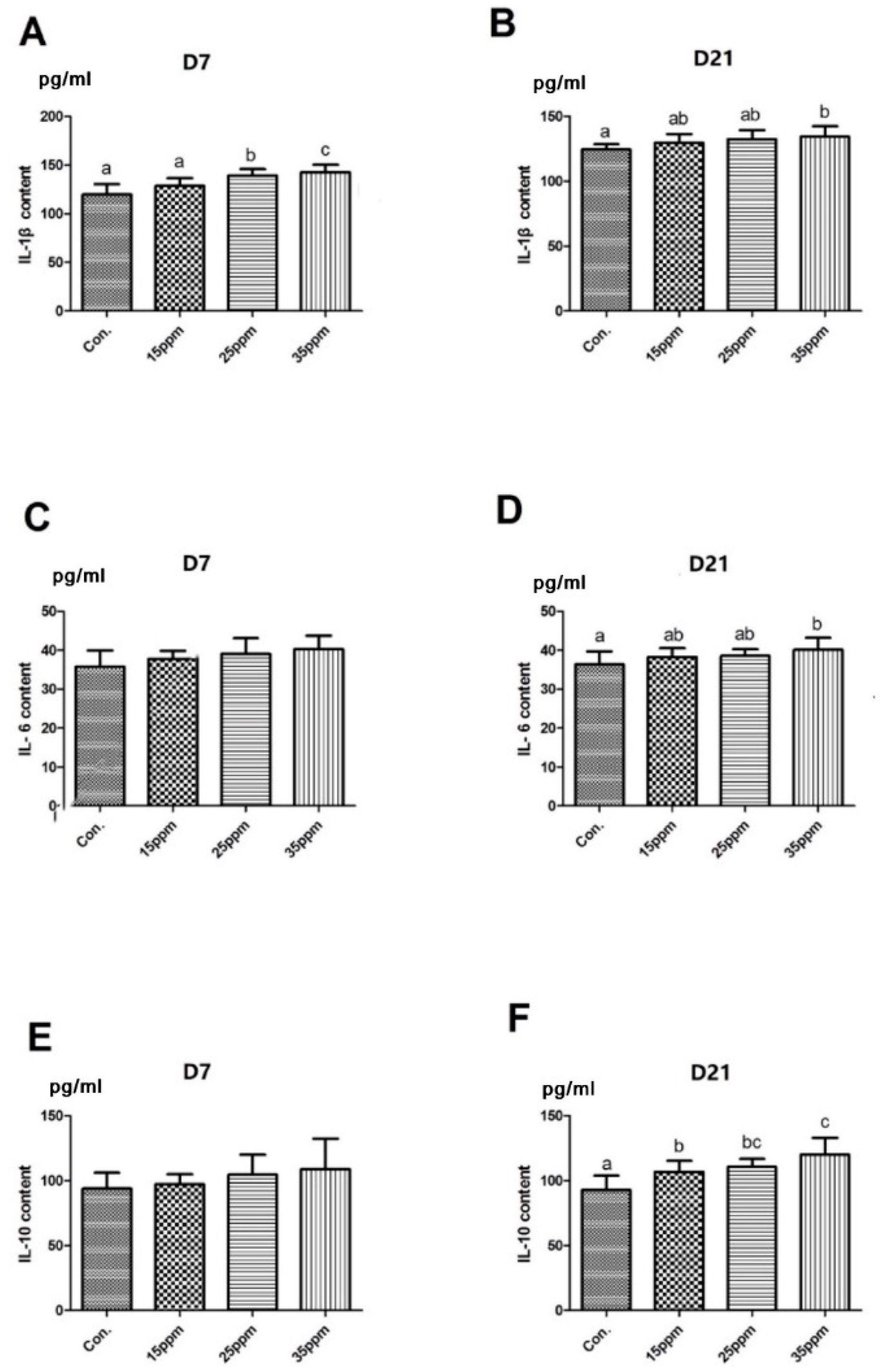
3.3. Effect of Ammonia on Inflammatory Cytokines in Lung Tissue of Broilers
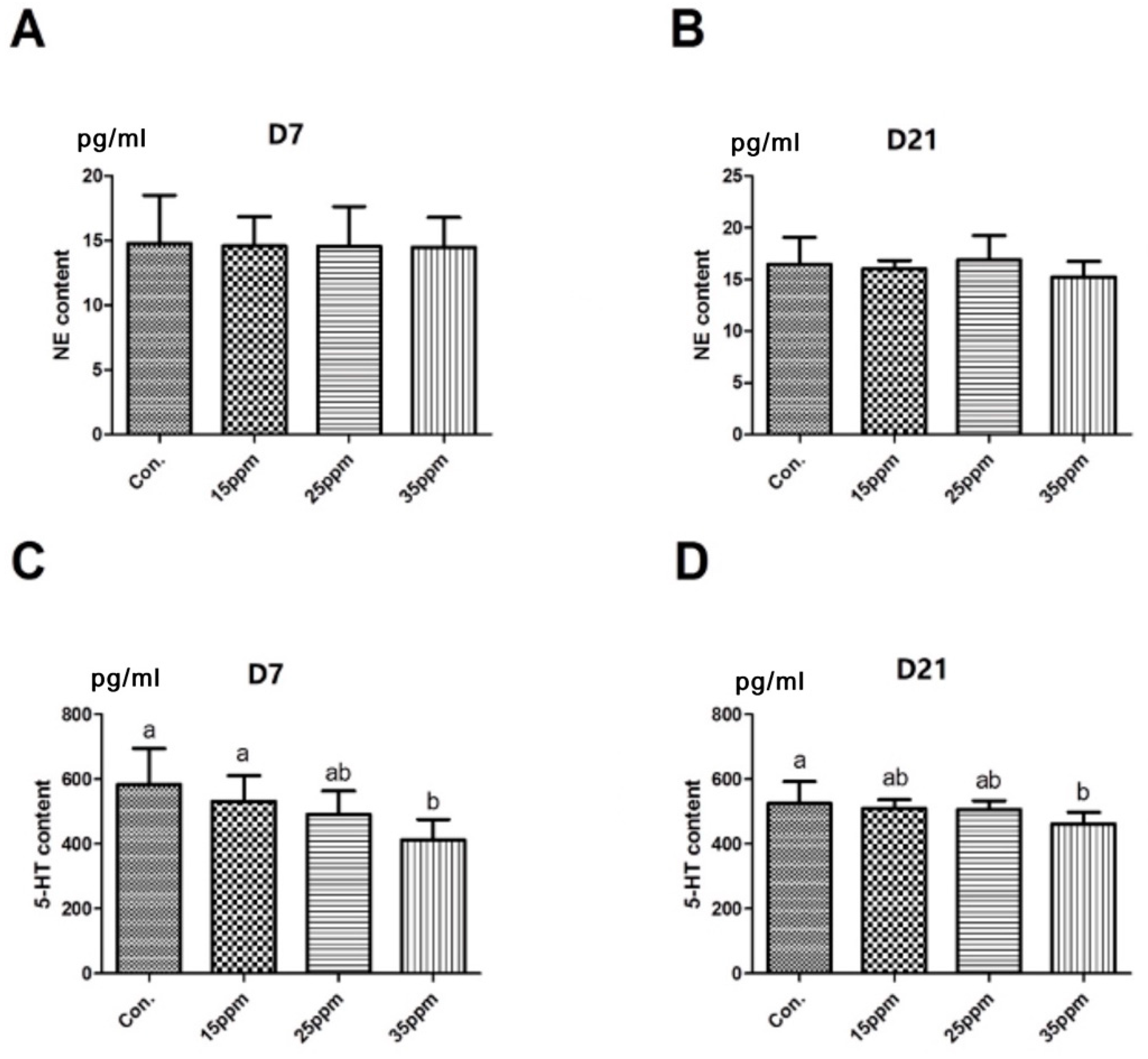
3.4. Effect of Ammonia on Neurotransmitters in Lung Tissue of Broilers
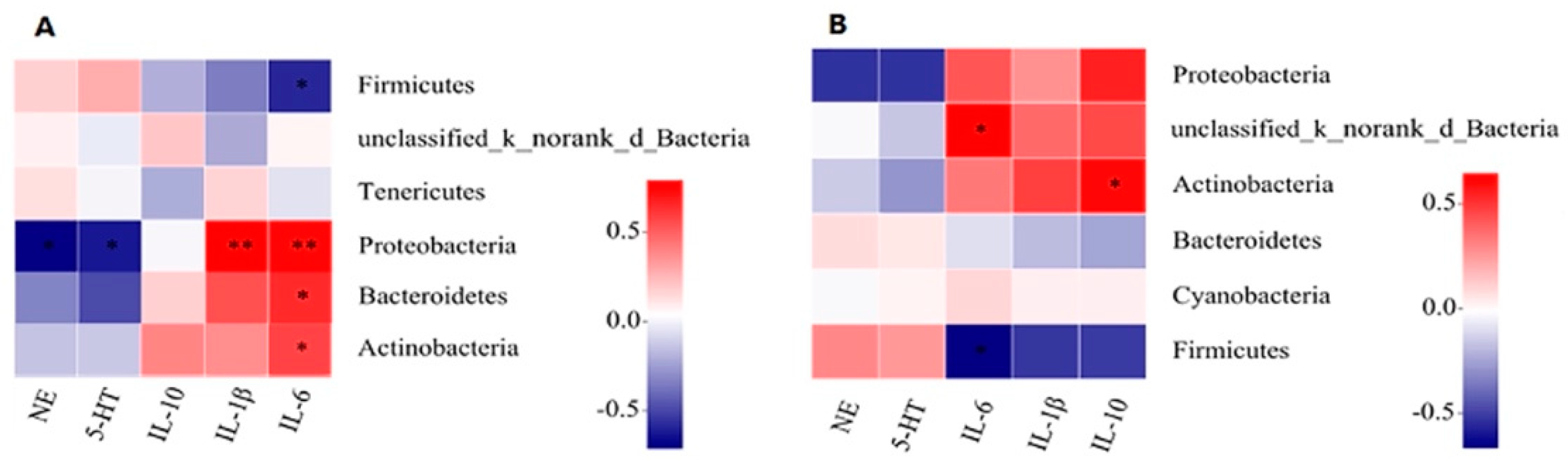
3.5. Correlation Analysis of Pulmonary Microflora with IL-1β, IL-6, IL-10, and NE, 5-HT
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koerkamp, P.G. Review on Emissions of Ammonia from Housing Systems for Laying Hens in Relation to Sources, Processes, Building Design and Manure Handling. J. Agric. Eng. Res. 1994, 59, 73–87. [Google Scholar] [CrossRef]
- Harper, L.A.; Sharpe, R.R.; Simmons, J.D. Ammonia Emissions from Swine Houses in the Southeastern United States. J. Environ. Qual. 2004, 33, 449–457. [Google Scholar] [CrossRef]
- Amon, B.; Kryvoruchko, V.; Fröhlich, M.; Amon, T.; Pöllinger, A.; Mösenbacher, I.; Hausleitner, A. Ammonia and Greenhouse Gas Emissions from a Straw Flow System for Fattening Pigs: Housing and Manure Storage. Livest. Sci. 2007, 112, 199–207. [Google Scholar] [CrossRef]
- Olanrewaju, H.; Miller, W.; Maslin, W.; Thaxton, J.; Dozier III, W.; Purswell, J.; Branton, S. Interactive Effects of Ammonia and Light Intensity on Ocular, Fear and Leg Health in Broiler Chickens. Int. J. Poult. Sci. 2007, 6, 762–769. [Google Scholar]
- Almuhanna, E.A.; Ahmed, A.S.; Yousif, Y. Effect of Air Contaminants on Poultry Immunological and Production Performance. Int. J. Poult. Sci. 2011, 10, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Yi, B.; Chen, L.; Sa, R.; Zhong, R.; Xing, H.; Zhang, H. High Concentrations of Atmospheric Ammonia Induce Alterations of Gene Expression in the Breast Muscle of Broilers (Gallus Gallus) Based on RNA-Seq. BMC Genom. 2016, 17, 598. [Google Scholar] [CrossRef] [Green Version]
- Ritz, C.; Fairchild, B.; Lacy, M. Implications of Ammonia Production and Emissions from Commercial Poultry Facilities: A Review. J. Appl. Poult. Res. 2004, 13, 684–692. [Google Scholar] [CrossRef]
- Wadhwa, R.; Paudel, K.R.; Mehta, M.; Shukla, S.D.; Sunkara, K.; Prasher, P.; Panth, N.; Goyal, R.; Chellappan, D.K.; Gupta, G.; et al. Beyond the Obvious: Smoking and Respiratory Infection Implications on Alzheimer’s Disease. CNS Neurol. Disord.-Drug Targets 2020, 19, 698–708. [Google Scholar] [CrossRef]
- Perry, V.H. The Influence of Systemic Inflammation on Inflammation in the Brain: Implications for Chronic Neurodegenerative Disease. Brain Behav. Immun. 2004, 18, 407–413. [Google Scholar] [CrossRef]
- Tan, Y.; Yang, T.; Liu, S.; Liu, H.; Xiang, Y.; Qu, F.; Li, H.; Qin, X. Infection with Respiratory Syncytial Virus Alters Peptidergic Innervation in the Lower Airways of Guinea-Pigs. Exp. Physiol. 2008, 93, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Caixia, L.; Yang, X.; Yurong, T.; Xiaoqun, Q. Involvement of Epigenetic Modification in Epithelial Immune Responses during Respiratory Syncytial Virus Infection. Microb. Pathog. 2019, 130, 186–189. [Google Scholar] [CrossRef]
- Qin, L.; Qiu, K.; Hu, C.; Wang, L.; Wu, G.; Tan, Y. Respiratory Syncytial Virus Promoted the Differentiation of Th17 Cells in Airway Microenvironment through Activation of Notch-1/Delta3. J. Med. Microbiol. 2019, 68, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P. Neurotransmitter Modulation by the Gut Microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef]
- Jameson, K.G.; Hsiao, E.Y. Linking the Gut Microbiota to a Brain Neurotransmitter. Trends Neurosci. 2018, 41, 413–414. [Google Scholar] [CrossRef] [PubMed]
- Man, W.H.; de Steenhuijsen Piters, W.A.; Bogaert, D. The Microbiota of the Respiratory Tract: Gatekeeper to Respiratory Health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Lin, X.; Jiao, H.; Uyanga, V.; Zhao, J.; Wang, X.; Li, H.; Zhou, Y.; Sun, S.; Lin, H. Mild Heat Stress Changes the Microbiota Diversity in the Respiratory Tract and the Cecum of Layer-Type Pullets. Poult. Sci. 2020, 99, 7015–7026. [Google Scholar] [CrossRef]
- Winter, S.E.; Bäumler, A.J. Dysbiosis in the Inflamed Intestine: Chance Favors the Prepared Microbe. Gut Microbes 2014, 5, 71–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beker, A.; Vanhooser, S.; Swartzlander, J.; Teeter, R. Atmospheric Ammonia Concentration Effects on Broiler Growth and Performance. J. Appl. Poult. Res. 2004, 13, 5–9. [Google Scholar] [CrossRef]
- Miles, D.; Branton, S.; Lott, B. Atmospheric Ammonia Is Detrimental to the Performance of Modern Commercial Broilers. Poult. Sci. 2004, 83, 1650–1654. [Google Scholar] [CrossRef]
- Kocaman, B.; Esenbuga, N.; Yildiz, A.; Laçin, E.; Macit, M. Effect of Environmental Conditions in Poultry Houses on the Performance of Laying Hens. Int. J. Poult. Sci. 2006, 5, 26–30. [Google Scholar]
- Anderson, D.; Beard, C.; Hanson, R. The Adverse Effects of Ammonia on Chickens Including Resistance to Infection with Newcastle Disease Virus. Avian Dis. 1964, 8, 369–379. [Google Scholar] [CrossRef]
- Brand, M.W.; Wannemuehler, M.J.; Phillips, G.J.; Proctor, A.; Overstreet, A.-M.; Jergens, A.E.; Orcutt, R.P.; Fox, J.G. The Altered Schaedler Flora: Continued Applications of a Defined Murine Microbial Community. ILAR J. 2015, 56, 169–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Asmakh, M.; Zadjali, F. Use of Germ-Free Animal Models in Microbiota-Related Research. J. Microbiol. Biotechnol. 2015, 25, 1583–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabbir, M.Z.; Malys, T.; Ivanov, Y.V.; Park, J.; Shabbir, M.A.B.; Rabbani, M.; Yaqub, T.; Harvill, E.T. Microbial Communities Present in the Lower Respiratory Tract of Clinically Healthy Birds in Pakistan. Poult. Sci. 2015, 94, 612–620. [Google Scholar] [CrossRef] [PubMed]
- NIU, J.; XIAO, C.; CHEN, D.; LI, X.; YANG, B.; YAN, S. Effects of Atmospheric Pollutants on the Respiratory Tract Microflora of SHR Rats. Chin. J. Microecol. 2016, 916–919. [Google Scholar]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial Signature of Dysbiosis in Gut Microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Reiche, E.M.V.; Nunes, S.O.V.; Morimoto, H.K. Stress, Depression, the Immune System, and Cancer. Lancet Oncol. 2004, 5, 617–625. [Google Scholar] [CrossRef]
- Li, C.; Lu, Q.; Tang, X.; Zhang, J.; Ding, Z.; Zhang, H. Influence of Ammonia Concentration on Growth Performance and Meat Quality of Broilers. Sci. Agric. Sin. 2014, 47, 4516–4523. [Google Scholar]
- Jennewein, C.; Tran, N.; Paulus, P.; Ellinghaus, P.; Eble, J.A.; Zacharowski, K. Novel Aspects of Fibrin (Ogen) Fragments during Inflammation. Mol. Med. 2011, 17, 568–573. [Google Scholar] [CrossRef]
- Liu, Q.X.; Zhou, Y.; Li, X.M.; Ma, D.D.; Xing, S.; Feng, J.H.; Zhang, M.H. Ammonia Induce Lung Tissue Injury in Broilers by Activating NLRP3 Inflammasome via Escherichia/Shigella. Poult. Sci. 2020, 99, 3402–3410. [Google Scholar] [CrossRef]
- Marc, D.T.; Ailts, J.W.; Campeau, D.C.A.; Bull, M.J.; Olson, K.L. Neurotransmitters Excreted in the Urine as Biomarkers of Nervous System Activity: Validity and Clinical Applicability. Neurosci. Biobehav. Rev. 2011, 35, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Luo, L. Effect of Air Benzo (a) Pyrene Exposure on Urine Neurotransmitter Level in 3-5 Years Old Preschool Children. Acad. J. Second Mil. Med. Univ. 2016, 37, 548–551. [Google Scholar]
- Cazzolau, M.; Matera, M.; D’amato, G.; Rossi, F. Effects of Serotonin on Airways: Recent Developments. Allergy 1995, 50, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Afergan, E.; Epstein, H.; Dahan, R.; Koroukhov, N.; Rohekar, K.; Danenberg, H.D.; Golomb, G. Delivery of Serotonin to the Brain by Monocytes Following Phagocytosis of Liposomes. J. Control. Release 2008, 132, 84–90. [Google Scholar] [CrossRef]
- Graeff, F.G.; Guimarães, F.S.; De Andrade, T.G.; Deakin, J.F. Role of 5-HT in Stress, Anxiety, and Depression. Pharmacol. Biochem. Behav. 1996, 54, 129–141. [Google Scholar] [CrossRef]
- Gaschler, G.J.; Zavitz, C.C.; Bauer, C.M.; Stämpfli, M.R. Mechanisms of Clearance of Nontypeable Haemophilus Influenzae from Cigarette Smoke-Exposed Mouse Lungs. Eur. Respir. J. 2010, 36, 1131–1142. [Google Scholar] [CrossRef] [Green Version]
- Moghaddam, S.J.; Clement, C.G.; De la Garza, M.M.; Zou, X.; Travis, E.L.; Young, H.W.; Evans, C.M.; Tuvim, M.J.; Dickey, B.F. Haemophilus Influenzae Lysate Induces Aspects of the Chronic Obstructive Pulmonary Disease Phenotype. Am. J. Respir. Cell Mol. Biol. 2008, 38, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Rizzatti, G.; Lopetuso, L.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A Common Factor in Human Diseases. BioMed Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, R.D.; Puybasset, L. The Brain–Lung–Brain Axis. Intensive Care Med. 2011, 37, 1054–1056. [Google Scholar] [CrossRef] [Green Version]
- Clarke, G.; Grenham, S.; Scully, P.; Fitzgerald, P.; Moloney, R.; Shanahan, F.; Dinan, T.; Cryan, J. The Microbiome-Gut-Brain Axis during Early Life Regulates the Hippocampal Serotonergic System in a Sex-Dependent Manner. Mol. Psychiatry 2013, 18, 666–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Content (%) | Items | Content (%) |
|---|---|---|---|
| Ingredients | Nutrient levels (2) | ||
| Corn | 56.51 | ME/(MJ/Kg) | 12.73 |
| Soybean meal | 35.52 | Crude protein | 20.07 |
| Soybean oil | 4.50 | Ca | 0.90 |
| NaCl | 0.30 | Available phosphorus | 0.40 |
| Limestone | 1.00 | Lys | 1.00 |
| CaHPO4 | 1.78 | Met | 0.42 |
| d L-Met | 0.11 | Met + Cys | 0.78 |
| Premix (1) | 0.28 | ||
| Total | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Liu, Q.; Zhou, Y.; Feng, J.; Zhang, M. Effects of Different Ammonia Concentrations on Pulmonary Microbial Flora, Lung Tissue Mucosal Morphology, Inflammatory Cytokines, and Neurotransmitters of Broilers. Animals 2022, 12, 261. https://doi.org/10.3390/ani12030261
Wang G, Liu Q, Zhou Y, Feng J, Zhang M. Effects of Different Ammonia Concentrations on Pulmonary Microbial Flora, Lung Tissue Mucosal Morphology, Inflammatory Cytokines, and Neurotransmitters of Broilers. Animals. 2022; 12(3):261. https://doi.org/10.3390/ani12030261
Chicago/Turabian StyleWang, Guangju, Qingxiu Liu, Ying Zhou, Jinghai Feng, and Minhong Zhang. 2022. "Effects of Different Ammonia Concentrations on Pulmonary Microbial Flora, Lung Tissue Mucosal Morphology, Inflammatory Cytokines, and Neurotransmitters of Broilers" Animals 12, no. 3: 261. https://doi.org/10.3390/ani12030261