

Management of Enteric Methane Emissions in Ruminants Using Feed Additives: A Review



{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
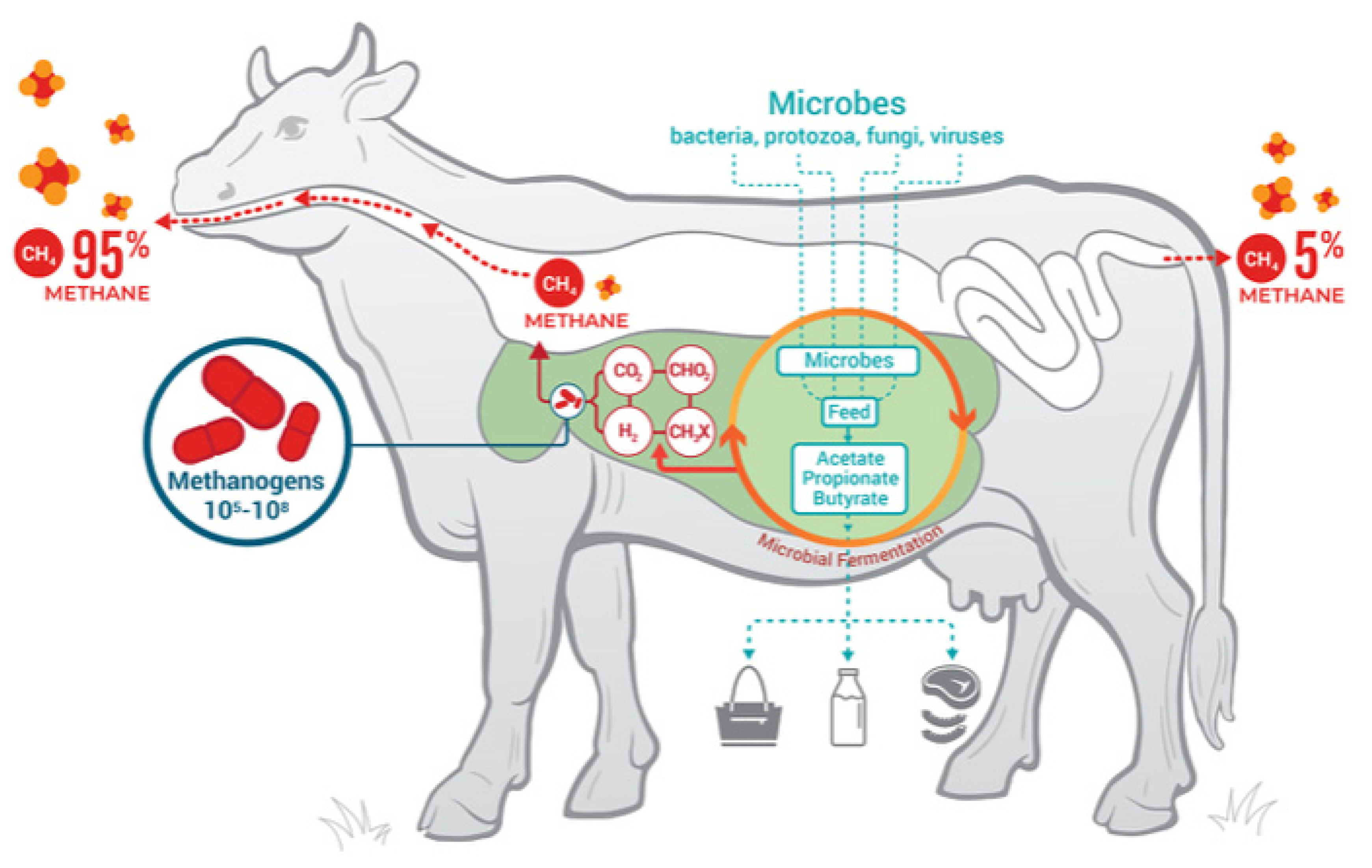
1. Introduction
2. Use of Methane Inhibitors
3. Use of Herbal Extracts
4. Use of Bee Propolis Extract
5. Use of Saponins
6. Use of Yeast
7. Use of Ionophores
8. Use of Organic Acids
9. Use of Exogenous Enzymes
10. Use of Nanoparticles
11. Use of Algae
12. Discussion
- feeding nitrates (for beef cattle)
- feeding dietary additives (for milking cows)
- possible toxicity to the ruminant
- potential environmental impacts/undesired side effects
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hook, S.E.; Wright, A.D.G.; McBride, B.W. Methanogens: Methane producers of the rumen and mitigation strategies. Archaea 2010, 2010, 945785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanbari Maman, L.; Palizban, F.; Fallah Atanaki, F.; Elmi Ghiasi, N.; Ariaeenejad, S.; Ghaffari, M.R.; Kavousi, K. Co-abundance analysis reveals hidden players associated with high methane yield phenotype in sheep rumen microbiome. Sci. Rep. 2020, 10, 4995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
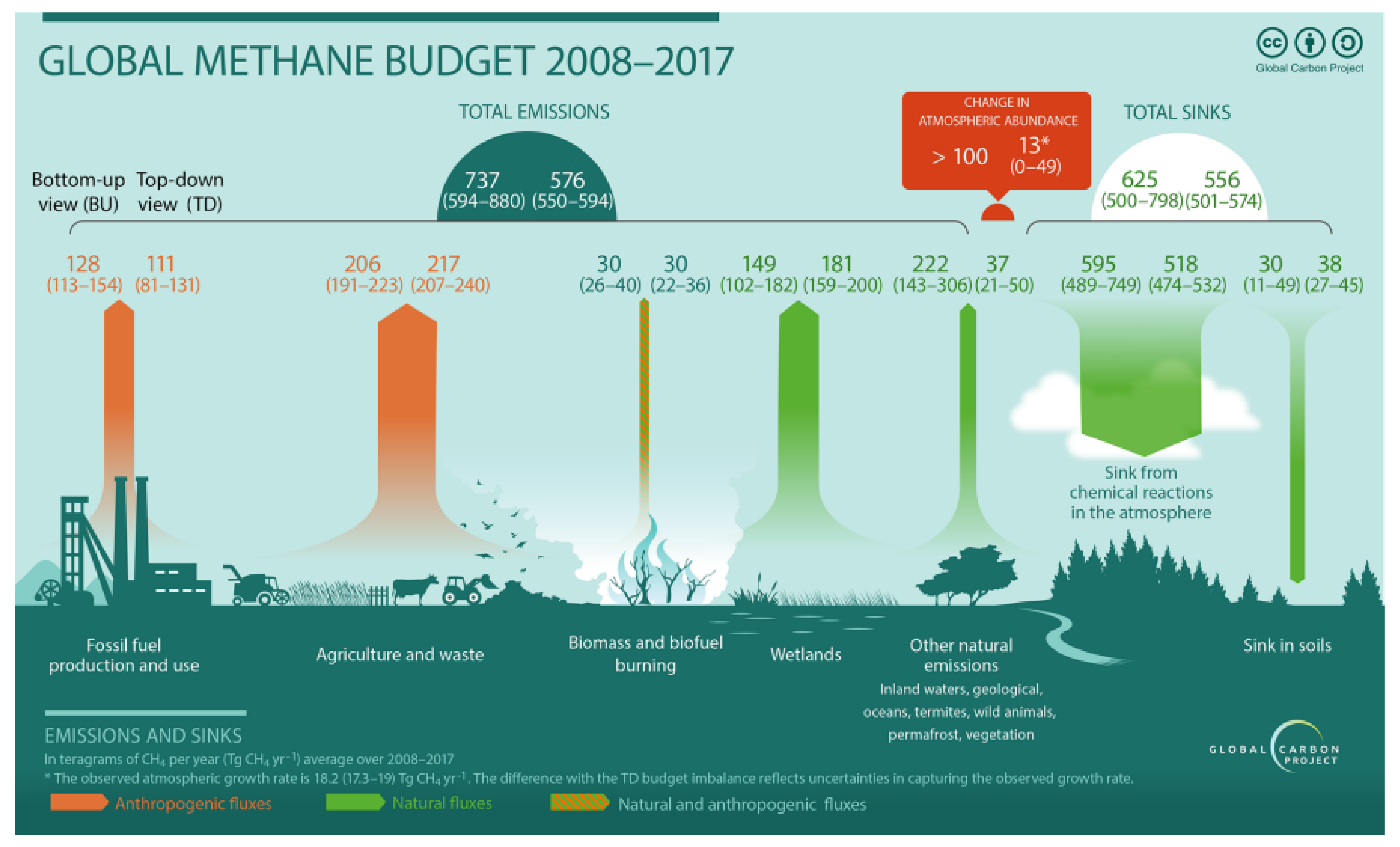
- Saunois, M.; Stavert, A.R.; Poulter, B.; Bousquet, P.; Canadell, J.G.; Jackson, R.B.; Zhuang, Q. The global methane budget 2000–2017. Earth Sys. Sci. Data 2020, 12, 1561–1623. [Google Scholar] [CrossRef]
- EPA, Overview of Greenhouse Gasses. Available online: https://www.epa.gov/ghgemissions/overview-greenhouse-gases, (accessed on 6 May 2022).
- Lackner, M.; Sajjadi, B.; Chen, W. (Eds.) Handbook of Climate Change Mitigation and Adaptation; Springer: Cham, Switzerland, 2022. [Google Scholar]
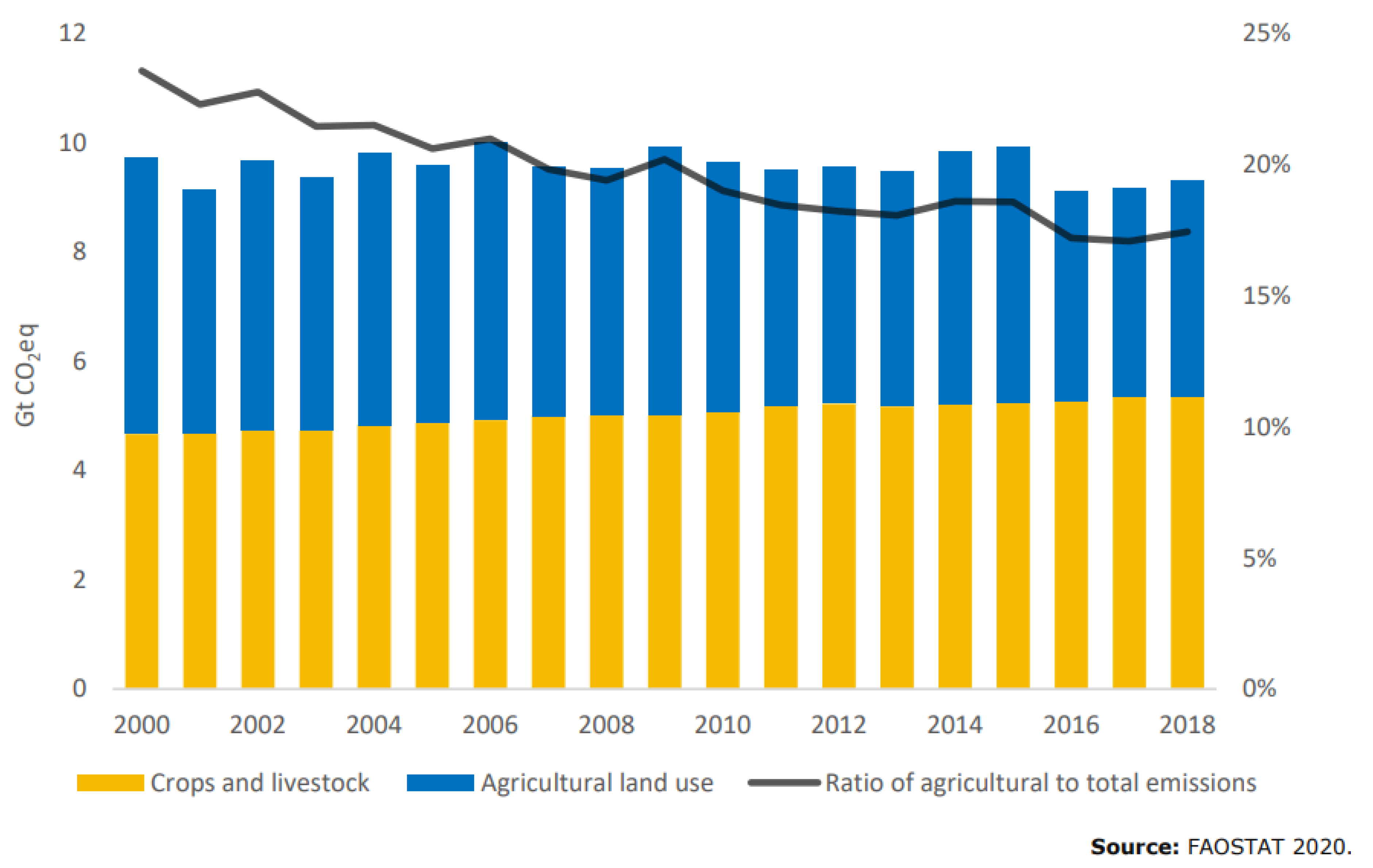
- FAO. FAO Stat Analytical Brief 18, Emissions Due to Agriculture—Global, Regional and Country Trends 2000–2018. 2018. Available online: https://www.fao.org/3/cb3808en/cb3808en.pdf (accessed on 6 May 2022).
- Department of Primary Industries and Regional Development, Government of Western Australia. 2022. Available online: https://www.agric.wa.gov.au/climate-change/carbon-farming-reducing-methane-emissions-cattle-using-feed-additives (accessed on 6 May 2022).
- Palangi, V.; Macit, M.; Nadaroglu, H.; Taghizadeh, A. Effects of green-synthesized CuO and ZnO nanoparticles on ruminal mitigation of methane emission to the enhancement of the cleaner environment. Biomass Convers. Biorefinery 2022. [Google Scholar] [CrossRef]
- Palangi, V.; Macit, M. Indictable mitigation of methane emission using some organic acids as additives towards a cleaner ecosystem. Waste Biomass Valorization 2021, 12, 4825–4834. [Google Scholar] [CrossRef]
- Eckard, R.J.; Grainger, C.; de Klein, C.A.M. Options for the abatement of methane and nitrous oxide from ruminant, production: A review. Livest. Sci. 2010, 130, 47–56. [Google Scholar] [CrossRef]
- Cottle, D.J.; Nolan, J.V.; Wiedemann, S.G. Ruminant enteric methane mitigation: A review. Anim. Prod. Sci. 2011, 51, 491–514. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Firkins, J.L.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Tricarico, J.M. Special topics—Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef] [Green Version]
- Broucek, J. Production of Methane Emissions from Ruminant Husbandry: A Review. J. Environ. Prot. 2014, 5, 51796. [Google Scholar] [CrossRef]
- Jeyanathan, J.; Martin, C.; Morgavi, D.P. The use of direct-fed microbials for mitigation of ruminant methane emissions: A review. Animal 2014, 8, 250–261. [Google Scholar] [CrossRef]
- Yáñez-Ruiz, D.R.; Bannink, A.; Dijkstra, J.; Kebreab, E.; Morgavi, D.P.; O’Kiely, P.; Hristov, A.N. Design, implementation and interpretation of in vitro batch culture experiments to assess enteric methane mitigation in ruminants—A review. Anim. Feed Sci. Technol. 2016, 216, 1–18. [Google Scholar] [CrossRef] [Green Version]
- McCauley, J.I.; Labeeuw, L.; Jaramillo-Madrid, A.C.; Nguyen, L.N.; Nghiem, L.D.; Chaves, A.V.; Ralph, P.J. Management of enteric methanogenesis in ruminants by algal-derived feed additives. Curr. Pollut. Rep. 2020, 6, 188–205. [Google Scholar] [CrossRef]
- Min, B.R.; Solaiman, S.; Waldrip, H.M.; Parker, D.; Todd, R.W.; Brauer, D. Dietary mitigation of enteric methane emissions from ruminants: A review of plant tannin mitigation options. Anim. Nut. 2020, 6, 231–246. [Google Scholar] [CrossRef]
- Cardoso-Gutierrez, E.; Aranda-Aguirre, E.; Robles-Jimenez, L.E.; Castelán-Ortega, O.A.; Chay-Canul, A.J.; Foggi, G.; González-Ronquillo, M. Effect of tannins from tropical plants on methane production from ruminants: A systematic review. Vet. Anim. Sci. 2021, 14, 100214. [Google Scholar] [CrossRef]
- Palangi, V.; Taghizadeh, A.; Abachi, S.; Lackner, M. Strategies to mitigate enteric methane emissions in ruminants: A review. Sustainability 2022, 14, 13229. [Google Scholar] [CrossRef]
- Zhenming, Z.; Meng, Q.; Yu, Z. Effects of methanogenic inhibitors on methane production and abundances of methanogens and cellulolytic bacteria in in vitro ruminal cultures. Appl. Environ. Microbiol. 2011, 77, 2634. [Google Scholar]
- Kim, H.; Lee, H.G.; Baek, Y.C.; Lee, S.; Seo, J. The effects of dietary supplementation with 3-nitrooxypropanol on enteric methane emissions, rumen fermentation, and production performance in ruminants: A meta-analysis. J. Anim. Sci. Technol. 2020, 62, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K.; Yu, Z. Combinations of nitrate, saponin, and sulfate additively reduce methane production by rumen cultures in vitro while not adversely affecting feed digestion, fermentation or microbial communities. Bioresour. Technol. 2014, 155, 129–135. [Google Scholar] [CrossRef]
- Newbold, J.R.; Van Zijderveld, S.M.; Hulshof, R.B.A.; Fokkink, W.B.; Leng, R.A.; Terencio, P.; Perdok, H.B. The effect of incremental levels of dietary nitrate on methane emissions in Holstein steers and performance in Nelore bulls. J. Anim. Sci. 2014, 92, 5032–5040. [Google Scholar] [CrossRef] [Green Version]
- Troy, S.M.; Duthie, C.A.; Hyslop, J.J.; Roehe, R.; Ross, D.W.; Wallace, R.J.; Rooke, J.A. Effectiveness of nitrate addition and increased oil content as methane mitigation strategies for beef cattle fed two contrasting basal diets. J. Anim. Sci. 2015, 93, 1815–1823. [Google Scholar] [CrossRef]
- Yang, C.; Rooke, J.A.; Cabeza, I.; Wallace, R.J. Nitrate and inhibition of ruminal methanogenesis: Microbial ecology, obstacles, and opportunities for lowering methane emissions from ruminant livestock. Front. Microbiol. 2016, 7, 132. [Google Scholar] [CrossRef] [PubMed]
- Balch, W.E.; Fox, G.E.; Magrum, L.J.; Woese, C.R.; Wolfe, R.S. Methanogens: Reevaluation of a unique biological group. Microbiol. Rev. 1979, 43, 260. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Meng, Q.; Zhou, Z.; Yu, Z. Ferric citrate, nitrate, saponin and their combinations affect in vitro ruminal fermentation, production of sulphide and methane and abundance of select microbial populations. J. Appl. Microbiol. 2019, 127, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.L.; Wolin, M.J. Inhibition of growth of methane-producing bacteria of the ruminant forestomach by hydroxymethylglutaryl∼SCoA reductase inhibitors. J. Dairy Sci. 2001, 84, 1445–1448. [Google Scholar] [CrossRef]
- Nkemka, V.N.; Beauchemin, K.A.; Hao, X. Treatment of feces from beef cattle fed the enteric methane inhibitor 3-nitrooxypropanol. Water. Sci. Technol. 2019, 80, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Rebelo, L.R.; Luna, I.C.; Messana, J.D.; Araujo, R.C.; Simioni, T.A.; Granja-Salcedo, Y.T.; Vitoa, E.S.; Lee, C.; Teixeira, I.A.M.A.; Rooke, J.A.; et al. Effect of replacing soybean meal with urea or encapsulated nitrate with or without elemental sulfur on nitrogen digestion and methane emissions in feedlot cattle. Anim. Feed Sci. Technol. 2019, 257, 114293. [Google Scholar] [CrossRef]
- Ramin, M.; Franco, M.; Roleda, M.Y.; Aasen, I.M.; Hetta, M.; Steinshamn, H. In vitro evaluation of utilisable crude protein and methane production for a diet in which grass silage was replaced by different levels and fractions of extracted seaweed proteins. Anim. Feed Sci. Technol. 2019, 255, 114225. [Google Scholar] [CrossRef]
- Alvarez-Hess, P.S.; Moate, P.J.; Williams, S.R.O.; Jacobs, J.L.; Beauchemin, K.A.; Hannah, M.C.; Durmic, Z.; Eckard, R.J. Effect of combining wheat grain with nitrate, fat or 3-nitrooxypropanol on in vitro methane production. Anim. Feed Sci. Technol. 2019, 256, 114237. [Google Scholar] [CrossRef]
- Natel, A.S.; Abdalla, A.L.; Araujo, R.C.; McManus, C.; Paim, T.P.; Filho, A.L.A.; Louvandini, P.; Nazato, C. Encapsulated nitrate replacing soybean meal changes in vitro ruminal fermentation and methane production in diets differing in concentrate to forage ratio. Anim. Sci. J. 2019, 90, 1350–1361. [Google Scholar] [CrossRef]
- Ugbogu, E.A.; Elghandour, M.M.; Ikpeazu, V.O.; Buendía, G.R.; Molina, O.M.; Arunsi, U.O.; Salem, A.Z. The potential impacts of dietary plant natural products on the sustainable mitigation of methane emission from livestock farming. J. Clean. Prod. 2019, 213, 915–925. [Google Scholar] [CrossRef]
- Zhou, R.; Wu, J.; Lang, X.; Liu, L.; Casper, D.P.; Wang, C.; Zhang, L.; Wei, S. Effects of oregano essential oil on in vitro ruminal fermentation, methane production, and ruminal microbial community. J. Dairy Sci. 2020, 103, 2303–2314. [Google Scholar] [CrossRef] [PubMed]
- Adegbeye, M.J.; Elghandour, M.M.; Monroy, J.C.; Abegunde, T.O.; Salem, A.Z.; Barbabosa-Pliego, A.; Faniy, T.O. Potential influence of Yucca extract as feed additive on greenhouse gases emission for a cleaner livestock and aquaculture farming—A review. J. Clean. Prod. 2019, 239, 118074. [Google Scholar] [CrossRef]
- Patra, A.K.; Min, B.R.; Saxena, J. Dietary tannins on microbial ecology of the gastrointestinal tract in ruminants. In Dietary Phytochemicals and Microbes; Springer: Dordrecht, The Netherlands, 2012; pp. 237–262. [Google Scholar]
- Pérez-Barbería, F.J.; Mayes, R.W.; Giráldez, J.; Sánchez-Pérez, D. Ericaceous species reduce methane emissions in sheep and red deer: Respiration chamber measurements and predictions at the scale of European heathlands. Sci. Total Environ. 2020, 714, 136738. [Google Scholar] [CrossRef] [PubMed]
- Fandiño, I.; Ferret, A.; Calsamiglia, S. Dose and combinations of anise oil and capsicum oleoresin as rumen fermentation modifiers in vitro and in vivo with high concentrate diets fed to Holstein beef heifers. Anim. Feed Sci. Technol. 2020, 260, 114363. [Google Scholar] [CrossRef]
- Hart, K.J.; Jones, H.G.; Waddams, K.E.; Worgan, H.J.; Zweifel, B.; Newbold, C.J. An essential oil blend decreases methane emissions and increases milk yield in dairy cows. Open J. Anim. Sci. 2019, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Canbolat, Ö.; Kalkan, H.; Karaman, Ș.; Filya, I. The effect of essential oils on the digestibility, rumen fermentation and microbial protein production. Kafkas Üniv. Vet. Fak. Der. 2011, 17, 557–565. [Google Scholar] [CrossRef]
- Jahani-Azizabadi, H.; Durmic, Z.; Vadhanabhuti, J.; Vercoe, P.E. Effect of some australian native shrubs essential oils on in vitro rumen microbial fermentation of a high-concentrate diet. J. Anim. Plant Sci. 2019, 29, 8–15. [Google Scholar]
- Pedraza-Hernandez, J.; Elghandour, M.M.M.Y.; Khusro, A.; Camacho-Diaz, L.M.; Vallejo, L.H.; Barbabosa-Pliego, A.; Salem, A.Z.M. Mitigation of ruminal gases production from goats using Moringa oleifera extract and live yeast culture for a cleaner agriculture environment. J. Clean. Prod. 2019, 234, 779–786. [Google Scholar] [CrossRef]
- Sinz, S.; Marquardt, S.; Soliva, C.R.; Braun, U.; Liesegang, A.; Kreuzer, M. Phenolic plant extracts are additive in their effects against in vitro ruminal methane and ammonia formation. Asian-Australas. J. Anim. Sci. 2019, 32, 966. [Google Scholar] [CrossRef] [Green Version]
- Wann, C.; Wanapat, M.; Mapato, C.; Ampapon, T.; Huang, B. Effect of bamboo grass (Tiliacora triandra, Diels) pellet supplementation on rumen fermentation characteristics and methane production in Thai native beef cattle. Asian-Australas. J. Anim. Sci. 2019, 32, 1153–1160. [Google Scholar] [CrossRef]
- Abdelrahman, S.M.; Li, R.H.; Elnahr, M.; Farouk, M.H.; Lou, Y. Effects of different levels of eucalyptus oil on methane production under in vitro conditions. Pol. J. Environ. Stud. 2019, 28, 1031–1042. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, N.; Shekhar, C.; Kumar, R.; Chaudhary, L.C.; Kamra, D.N. Effect of peppermint (Mentha piperita) oil on in vitro methanogenesis and fermentation of feed with buffalo rumen liquor. Anim. Feed Sci. Technol. 2009, 148, 321–327. [Google Scholar] [CrossRef]
- Roca-Fernández, A.I.; Dillard, S.L.; Soder, K.J. Ruminal fermentation and enteric methane production of legumes containing condensed tannins fed in continuous culture. J. Dairy Sci. 2020, 103, 7028–7038. [Google Scholar] [CrossRef] [PubMed]
- Santos, N.W.; Zeoula, L.M.; Yoshimura, E.H.; Machado, E.; Macheboeuf, D.; Cornu, A. Brazilian propolis extract used as an additive to decrease methane emissions from the rumen microbial population in vitro. Trop. Anim. Health Prod. 2016, 48, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Morsy, A.S.; Soltan, Y.A.; Sallam, S.M.A.; Kreuzer, M.; Alencar, S.M.; Abdalla, A.L. Comparison of the in vitro efficiency of supplementary bee propolis extracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation of methane. Anim. Feed Sci. Technol. 2015, 199, 51–60. [Google Scholar] [CrossRef]
- Ehtesham, S.; Vakili, A.R.; Danesh Mesgaran, M.; Bankova, V. The effects of phenolic compounds in Iranian propolis extracts on in vitro rumen fermentation, methane production and microbial population. Iranian J. Appl. Anim. Sci. 2018, 8, 33–41. [Google Scholar]
- Morsy, A.S.; Soltan, Y.A.; El-Zaiat, H.M.; Alencar, S.M.; Abdalla, A.L. Bee propolis extract as a phytogenic feed additive to enhance diet digestibility, rumen microbial biosynthesis, mitigating methane formation and health status of late pregnant ewes. Anim. Feed Sci. Technol. 2021, 273, 114834. [Google Scholar] [CrossRef]
- Kara, K.; Güçlü, B.K.; Oğuz, F.K. Use of propolis and phenolic acids in ruminant nutrition. Erciyes Üniv. Vet. Fak. Der. 2014, 11, 43–53. [Google Scholar]
- Jayanegara, A.; Wina, E.; Takahashi, J. Meta-analysis on methane mitigating properties of saponin-rich sources in the rumen: Influence of addition levels and plant sources. Asian-Australas. J. Anim. Sci. 2014, 27, 1426. [Google Scholar] [CrossRef]
- Hess, H.D.; Beuret, R.A.; Lotscher, M.; Hindrichsen, I.K.; Machmuller, A.; Carulla, J.E.; Kreuzer, M. Ruminal fermentation, methanogenesis and nitrogen utilization of sheep receiving tropical grass hay-concentrate diets offered with Sapindus saponaria fruits and Cratylia argentea foliage. Anim. Sci. 2004, 79, 177–189. [Google Scholar] [CrossRef]
- Wang, C.J.; Wang, S.P.; Zhou, H. Influences of flavomycin, ropadiar, and saponin on nutrient digestibility, rumen fermentation, and methane emission from sheep. Anim. Feed Sci. Technol. 2009, 148, 157–166. [Google Scholar] [CrossRef]
- Klita, P.T.; Mathison, G.W.; Fenton, T.W.; Hardin, R.T. Effects of alfalfa root saponins on digestive function in sheep. J. Anim. Sci. 1996, 74, 1144–1156. [Google Scholar] [CrossRef] [PubMed]
- Pen, B.; Takaura, K.; Yamaguchi, S.; Asa, R.; Takahashi, J. Effects of Yucca schidigera and Quillaja saponaria with or without β 1–4 galacto-oligosaccharides on ruminal fermentation, methane production and nitrogen utilization in sheep. Anim. Feed Sci. Technol. 2007, 138, 75–88. [Google Scholar] [CrossRef]
- Śliwiński, B.J.; Kreuzer, M.; Wettstein, H.R.; Machmüller, A. Rumen fermentation and nitrogen balance of lambs fed diets containing plant extracts rich in tannins and saponins, and associated emissions of nitrogen and methane. Arch. Anim. Nutr. 2002, 56, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, T.; Chen, D.; Zhang, N.; Si, B.; Deng, K.; Diao, Q. Effects of tea saponin supplementation on nutrient digestibility, methanogenesis, and ruminal microbial flora in Dorper crossbred ewe. Animals 2019, 9, 29. [Google Scholar] [CrossRef] [Green Version]
- Van Zijderveld, S.M.; Dijkstra, J.; Perdok, H.B.; Newbold, J.R.; Gerrits, W.J.J. Dietary inclusion of diallyl disulfide, yucca powder, calcium fumarate, an extruded linseed product, or medium-chain fatty acids does not affect methane production in lactating dairy cows. J. Dairy Sci. 2011, 94, 3094–3104. [Google Scholar] [CrossRef] [Green Version]
- Holtshausen, L.; Chaves, A.V.; Beauchemin, K.A.; McGinn, S.M.; McAllister, T.A.; Odongo, N.E.; Benchaar, C. Feeding saponin-containing Yucca schidigera and Quillaja saponaria to decrease enteric methane production in dairy cows. J. Dairy Sci. 2009, 92, 2809–2821. [Google Scholar] [CrossRef]
- Yuan, Z.P.; Zhang, C.M.; Zhou, L.; Zou, C.X.; Guo, Y.Q.; Li, W.T.; Wu, Y.M. Inhibition of methanogenesis by tea saponin and tea saponin plus disodium fumarate in sheep. J. Anim. Feed Sci. 2007, 16, 560–565. [Google Scholar] [CrossRef] [Green Version]
- Santoso, B.; Mwenya, B.; Sar, C.; Gamo, Y.; Kobayashi, T.; Morikawa, R.; Takahashi, J. Effects of supplementing galacto-oligosaccharides, Yucca schidigera or nisin on rumen methanogenesis, nitrogen and energy metabolism in sheep. Livest. Prod. Sci. 2004, 91, 209–217. [Google Scholar] [CrossRef]
- Mao, H.L.; Wang, J.K.; Zhou, Y.Y.; Liu, J.X. Effects of addition of tea saponins and soybean oil on methane production, fermentation and microbial population in the rumen of growing lambs. Livest. Sci. 2010, 129, 56–62. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. The effect and mode of action of saponins on the microbial populations and fermentation in the rumen and ruminant production. Nut. Res. Rev. 2009, 22, 204–219. [Google Scholar] [CrossRef] [PubMed]
- Morgavi, D.P.; Eugène, M.; Martin, C.; Doreau, M. Reducing methane emissions in ruminants: Is it an achievable goal. Challenging Strategies to Promote the Sheep and Goat Sector in the Current Global Context. In Options Méditerranéennes: Série A. Séminaires Méditerranéens; CIHEAM: Paris, France, 2011; pp. 65–73. [Google Scholar]
- Wina, E.; Muetzel, S.; Hoffmann, E.; Makkar, H.P.S.; Becker, K. Saponins containing methanol extract of Sapindus rarak affect microbial fermentation, microbial activity and microbial community structure in vitro. Anim. Feed Sci. Technol. 2005, 121, 159–174. [Google Scholar] [CrossRef]
- Tan, C.; Ramírez-Restrepo, C.A.; Shah, A.M.; Hu, R.; Bell, M.; Wang, Z.; McSweeney, C. The community structure and microbial linkage of rumen protozoa and methanogens in response to the addition of tea seed saponins in the diet of beef cattle. Anim. Feed Sci. Technol. 2020, 11, 80. [Google Scholar] [CrossRef] [PubMed]
- Newbold, C.J.; De La Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The role of ciliate protozoa in the rumen. Front. Microbiol. 2015, 6, 1313. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.J.; McEwan, N.R.; McIntosh, F.M.; Teferedegne, B.; Newbold, C.J. Natural products as manipulators of rumen fermentation. Asian-Australas. J. Anim. Sci. 2002, 15, 1458–1468. [Google Scholar] [CrossRef]
- Desnoyers, M.; Giger-Reverdin, S.; Bertin, G.; Duvaux-Ponter, C.; Sauvant, D. Meta-analysis of the influence of Saccharomyces cerevisiae supplementation on ruminal parameters and milk production of ruminants. J. Dairy Sci. 2009, 92, 1620–1632. [Google Scholar] [CrossRef] [Green Version]
- O’brien, M.; Navarro-Villa, A.; Purcell, P.J.; Boland, T.M.; O’Kiely, P. Reducing in vitro rumen methanogenesis for two contrasting diets using a series of inclusion rates of different additives. Anim. Prod. Sci. 2014, 54, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Chaucheyras, F.; Fonty, G.; Bertin, G.; Gouet, P. In vitro H2 utilization by a ruminal acetogenic bacterium cultivated alone or in association with an Archaea methanogen is stimulated by a probiotic strain of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1995, 61, 3466–3467. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, C.; Wills, D.A.; Yan, T. Effects of dietary active dried yeast (Saccharomyces cerevisiae) supply at two levels of concentrate on energy and nitrogen utilisation and methane emissions of lactating dairy cows. Anim. Prod. Sci. 2017, 57, 656–664. [Google Scholar] [CrossRef]
- Bayat, A.R.; Kairenius, P.; Stefański, T.; Leskinen, H.; Comtet-Marre, S.; Forano, E.; Shingfield, K.J. Effect of camelina oil or live yeasts (Saccharomyces cerevisiae) on ruminal methane production, rumen fermentation, and milk fatty acid composition in lactating cows fed grass silage diets. J. Dairy Sci. 2015, 98, 3166–3181. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.; Harper, M.; Melgar, A.; Compart, D.P.; Hristov, A.N. Effects of Saccharomyces cerevisiae-based direct-fed microbial and exogenous enzyme products on enteric methane emission and productivity in lactating dairy cows. J. Dairy Sci. 2019, 102, 6065–6075. [Google Scholar] [CrossRef] [PubMed]
- Darabighane, B.; Salem, A.Z.M.; Aghjehgheshlagh, F.M.; Mahdavi, A.; Zarei, A.; Elghandour, M.M.M.Y.; López, S. Environmental efficiency of Saccharomyces cerevisiae on methane production in dairy and beef cattle via a meta-analysis. Environ. Sci. Pollut. 2019, 26, 3651–3658. [Google Scholar] [CrossRef] [PubMed]
- Mwenya, B.; Santoso, B.; Sar, C.; Gamo, Y.; Kobayashi, T.; Arai, I.; Takahashi, J. Effects of including β1–4 galacto-oligosaccharides, lactic acid bacteria or yeast culture on methanogenesis as well as energy and nitrogen metabolism in sheep. Anim. Feed Sci. Technol. 2004, 115, 313–326. [Google Scholar] [CrossRef]
- Lu, Q.; Wu, J.; Wang, M.; Zhou, C.; Han, X.; Odongo, E.N.; Tang, S. Effects of dietary addition of cellulase and a Saccharomyces cerevisiae fermentation product on nutrient digestibility, rumen fermentation and enteric methane emissions in growing goats. Arch. Anim. Nut. 2016, 70, 224–238. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Kalscheur, K.F.; Huhtanen, P.; Faciola, A.P. Effects of ruminal protozoa on methane emissions in ruminants—A meta-analysis. J. Dairy Sci. 2022, 105, 7482–7491. [Google Scholar] [CrossRef]
- Mutsvangwa, T.; Edwards, I.E.; Topps, J.H.; Paterson, G.F.M. The effect of dietary inclusion of yeast culture (Yea-Sacc) on patterns of rumen fermentation, food intake and growth of intensively fed bulls. Anim. Sci. 1992, 55, 35–40. [Google Scholar] [CrossRef]
- Shibata, M.; Terada, F. Factors affecting methane production and mitigation in ruminants. Anim. Sci. J. 2010, 81, 2–10. [Google Scholar] [CrossRef]
- McGuffey, R.K.; Richardson, L.F.; Wilkinson, J.I.D. Ionophores for dairy cattle: Current status and future outlook. J. Dairy Sci. 2001, 84, E194–E203. [Google Scholar] [CrossRef]
- Guan, H.; Wittenberg, K.M.; Ominski, K.H.; Krause, D.O. Efficacy of ionophores in cattle diets for mitigation of enteric methane. J. Anim. Sci. 2006, 84, 1896–1906. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Mohini, M.; Malla, B.A.; Mondal, G.; Pandita, S. Effects of monensin feeding on performance, nutrient utilization and enteric methane production in growing buffalo heifers. Trop. Anim. Health Pro. 2019, 51, 859–866. [Google Scholar] [CrossRef]
- Palangi, V. Effects of Processing Legume Forages with Organic Acids on In Vitro Gas Production, Rumen Fermantation and Methan Production. Ph.D. Thesis, Ataturk University, Erzurum, Turkey, 2019. [Google Scholar]
- Kara, K.; Aktuğ, E.; Çağrı, A.; Güçlü, B.K.; Baytok, E. Effect of formic acid on in vitro ruminal fermentation and methane emission. Turkish J. Agric. Food Sci. Technol. 2015, 3, 856–860. [Google Scholar] [CrossRef] [Green Version]
- Partanen, K.; Jalava, T. Effects of some organic acids and salts on microbial fermentation in the digestive tract of piglets estimated using an in vitro gas production technique. Agric. Food Sci. 2005, 14, 311–324. [Google Scholar] [CrossRef] [Green Version]
- Meale, S.J.; Beauchemin, K.A.; Hristov, A.N.; Chaves, A.V.; McAllister, T.A. Board-invited review: Opportunities and challenges in using exogenous enzymes to improve ruminant production. J. Anim. Sci. 2014, 92, 427–442. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.X.; Zou, Y.; Wang, M.; Salem, A.Z.M.; Odongo, N.E.; Zhou, C.S.; Kang, J.H. Effects of Exogenous Cellulase Source on In Vitro Fermentation Characteristics and Methane Production of Crop Straws and Grasses. Anim. Nutr. Feed Technol. 2013, 13, 489–505. [Google Scholar]
- He, Z.X.; Yang, L.Y.; Yang, W.Z.; Beauchemin, K.A.; Tang, S.X.; Huang, J.Y.; Tan, Z.L. Efficacy of exogenous xylanases for improving in vitro fermentation of forages. J. Agric. Sci. 2015, 153, 538–553. [Google Scholar] [CrossRef]
- Mohamed, M.A.E.; Yangchun, C.; Bodinga, B.M.; Lixin, Z.; Zekun, Y.; Lihui, L.; Wen, L. Research article effect of exogenous fibrolytic enzymes on ruminal fermentation and gas production by RUSITEC, in vitro Abomasum and Ileum digestibility. Int. J. Pharmacol. 2017, 13, 1020–1028. [Google Scholar] [CrossRef] [Green Version]
- McGinn, S.M.; Beauchemin, K.A.; Coates, T.; Colombatto, D. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. J. Anim. Sci. 2004, 82, 3346–3356. [Google Scholar] [CrossRef]
- Chung, Y.H.; Zhou, M.; Holtshausen, L.; Alexander, T.W.; McAllister, T.A.; Guan, L.L.; Beauchemin, K.A. A fibrolytic enzyme additive for lactating Holstein cow diets: Ruminal fermentation, rumen microbial populations, and enteric methane emissions. J. Dairy Sci. 2012, 95, 1419–1427. [Google Scholar] [CrossRef]
- Arriola, K.G.; Kim, S.C.; Staples, C.R.; Adesogan, A.T. Effect of fibrolytic enzyme application to low-and high-concentrate diets on the performance of lactating dairy cattle. J. Dairy Sci. 2011, 94, 832–841. [Google Scholar] [CrossRef] [Green Version]
- Abdelsalam, E.; Samer, M.; Attia, Y.A.; Abdel-Hadi, M.A.; Hassan, H.E.; Badr, Y. Comparison of nanoparticles effects on gas and methane production from anaerobic digestion of cattle dung slurry. Renew. Energy 2016, 87, 592–598. [Google Scholar] [CrossRef]
- Hernández-Sierra, J.F.; Ruiz, F.; Pena, D.C.C.; Martínez-Gutiérrez, F.; Martínez, A.E.; Guillén, A.D.J.P.; Castañón, G.M. The antimicrobial sensitivity of Streptococcus mutans to nanoparticles of silver, zinc oxide, and gold. Nanomed. Nanotechnol. Biol. Med. 2008, 4, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Fujinawa, K.; Nagoya, M.; Kouzuma, A.; Watanabe, K. Conductive carbon nanoparticles inhibit methanogens and stabilize hydrogen production in microbial electrolysis cells. Appl. Microbiol. Biotechnol. 2019, 103, 6385–6392. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Liu, H.; Zhang, Y.; Cui, M.H.; Fu, B.; Liu, H.B. Insight into sludge anaerobic digestion with granular activated carbon addition: Methanogenic acceleration and methane reduction relief. Bioresour. Technol. 2021, 319, 124131. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Si, H.B.; Wang, M.; Lin, B.; Deng, J.P.; Tan, L.W.; Tan, Z.L. Effects of elemental magnesium and magnesium oxide on hydrogen, methane and volatile fatty acids production in in vitro rumen batch cultures. Anim. Feed Sci. Technol. 2019, 252, 74–82. [Google Scholar] [CrossRef]
- Kazemi, M.; Vatandoost, M. The effect of different levels of magnesium oxide with high purity on digestion-fermentation characteristics and methane emissions of a high-concentrate diet in the in vitro batch culture. J. Anim. Environ. 2019, 11, 51–62. [Google Scholar]
- Salem, A.Z.M.; Ammar, H.; Lopez, S.; Gohar, Y.M.; González, J.S. Sensitivity of ruminal bacteria isolates of sheep, cattle and buffalo to some heavy metals. Anim. Feed Sci. Technol. 2011, 163, 143–149. [Google Scholar] [CrossRef]
- Chen, J.; Wang, W.; Wang, Z. Effect of nano-zinc oxide supplementation on rumen fermentation in vitro. Chin. J. Anim. Nutr. 2011, 23, 1415–1421. [Google Scholar]
- Chanzanagh, E.G.; Seifdavati, J.; Gheshlagh, F.M.A.; Benamar, H.A.; Sharifi, R.S. Effect of ZnO nanoparticles on in vitro gas production of some animal and plant protein sources. Kafkas Üniv. Vet. Fak. Der. 2018, 24, 25–32. [Google Scholar]
- Maorong, W.; Fang, M.; Wenbin, Y.; Yingxiang, H.; Chaohua, M.; Feng, W.; Yao, C. Influence of copper supplementation on nitrogen metabolism and volatile fatty acid production of mixed ruminal microbial growth in continuous culture flow-through fermentors. Chin. Agric. Sci. Bullet. 2008, 24, 19. [Google Scholar]
- Hernández-Sánchez, D.; Cervantes-Gómez, D.; Ramírez-Bribiesca, J.E.; Cobos-Peralta, M.; Pinto-Ruiz, R.; Astigarraga, L.; Gere, J.I. The influence of copper levels on in vitro ruminal fermentation, bacterial growth and methane production. J. Sci. Food Agric. 2019, 99, 1073–1077. [Google Scholar] [CrossRef]
- Anele, U.Y.; Yang, W.Z.; McGinn, P.J.; Tibbetts, S.M.; McAllister, T.A. Ruminal in vitro gas production, dry matter digestibility, methane abatement potential, and fatty acid biohydrogenation of six species of microalgae. Can. J. Anim. Sci. 2016, 96, 354–363. [Google Scholar] [CrossRef] [Green Version]
- Brooke Charles, G.; Roque Breanna, M.; Shaw, C.; Najafi, N.; Gonzalez, M.; Pfefferlen, A.; De Anda, V.; Ginsburg David, W.; Harden Maddelyn, C.; Nuzhdin Sergey, V.; et al. Methane reduction potential of two pacific coast macroalgae during in vitro ruminant fermentation. Front. Mar. Sci. 2020, 7, 561. [Google Scholar] [CrossRef]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Giger-Reverdin, S.; Lessire, M.; Lebas, F.; Ankers, P. Seaweeds for livestock diets: A review. Anim. Feed Sci. Technol. 2016, 212, 1–17. [Google Scholar] [CrossRef]
- Machado, L.; Magnusson, M.; Paul, N.A.; de Nys, R.; Tomkins, N. Effects of marine and freshwater macroalgae on in vitro total gas and methane production. PLoS ONE 2014, 9, e85289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glasson, C.R.; Kinley, R.D.; de Nys, R.; King, N.; Adams, S.L.; Packer, M.A.; Magnusson, M. Benefits and risks of including the bromoform containing seaweed Asparagopsis in feed for the reduction of methane production from ruminants. Algal Res. 2022, 64, 102673. [Google Scholar] [CrossRef]
- Min, B.R.; Parker, D.; Brauer, D.; Waldrip, H.; Lockard, C.; Hales, K.; Augyte, S. The role of seaweed as a potential dietary supplementation for enteric methane mitigation in ruminants: Challenges and opportunities. Anim. Nut. 2021, 7, 1371–1387. [Google Scholar] [CrossRef]
- DSM. Minimizing Methane from Cattle. 2022. Available online: https://www.dsm.com/corporate/sustainability/our-purpose/minimizing-methane-from-cattle.html (accessed on 22 August 2022).
- Pure. Available online: https://pure.au.dk/ws/files/197951334/Notat_3NOP_BOVAER_150920.pdf (accessed on 6 May 2022).
- mla (Meat & Livestock Australia). 2022. Available online: https://www.mla.com.au/news-and-events/industry-news/the-feed-additive-reducing-methane-emissions-by-up-to-90/ (accessed on 28 August 2022).
- Black, J.L.; Davison, T.M.; Box, I. Methane emissions from ruminants in australia: Mitigation potential and applicability of mitigation strategies. Animals 2021, 11, 951. [Google Scholar] [CrossRef]
- Gill, M.; Smith, P.; Wilkinson, J.M. Mitigating climate change: The role of domestic livestock. Animal 2010, 4, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Martínez, J.B.G.; Pearce, J.M.; Throup, J.; Cates, J.; Lackner, M.; Denkenberger, D.C. Methane single cell protein: Potential to secure a global protein supply against catastrophic food shocks. Front. Bioeng. Biotechnol. 2022, 10, 906704. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palangi, V.; Lackner, M. Management of Enteric Methane Emissions in Ruminants Using Feed Additives: A Review. Animals 2022, 12, 3452. https://doi.org/10.3390/ani12243452
Palangi V, Lackner M. Management of Enteric Methane Emissions in Ruminants Using Feed Additives: A Review. Animals. 2022; 12(24):3452. https://doi.org/10.3390/ani12243452
Chicago/Turabian StylePalangi, Valiollah, and Maximilian Lackner. 2022. "Management of Enteric Methane Emissions in Ruminants Using Feed Additives: A Review" Animals 12, no. 24: 3452. https://doi.org/10.3390/ani12243452