
Effects of Paratuberculosis Vaccination at Different Ages in a Dairy Goat Herd: A 2-Year Follow-Up
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Information
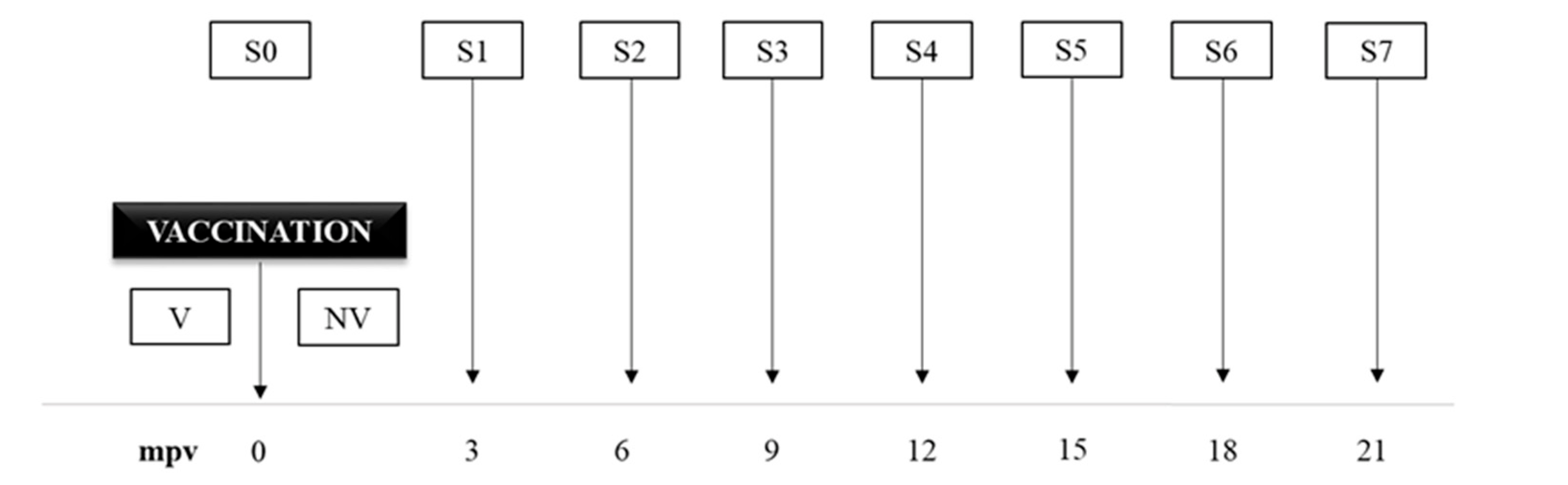
2.2. Animals and Experimental Design
2.3. Clinical Follow-Up and Necropsies
2.4. Peripheral Immune Response Evaluation
2.5. Statistical Analysis
3. Results
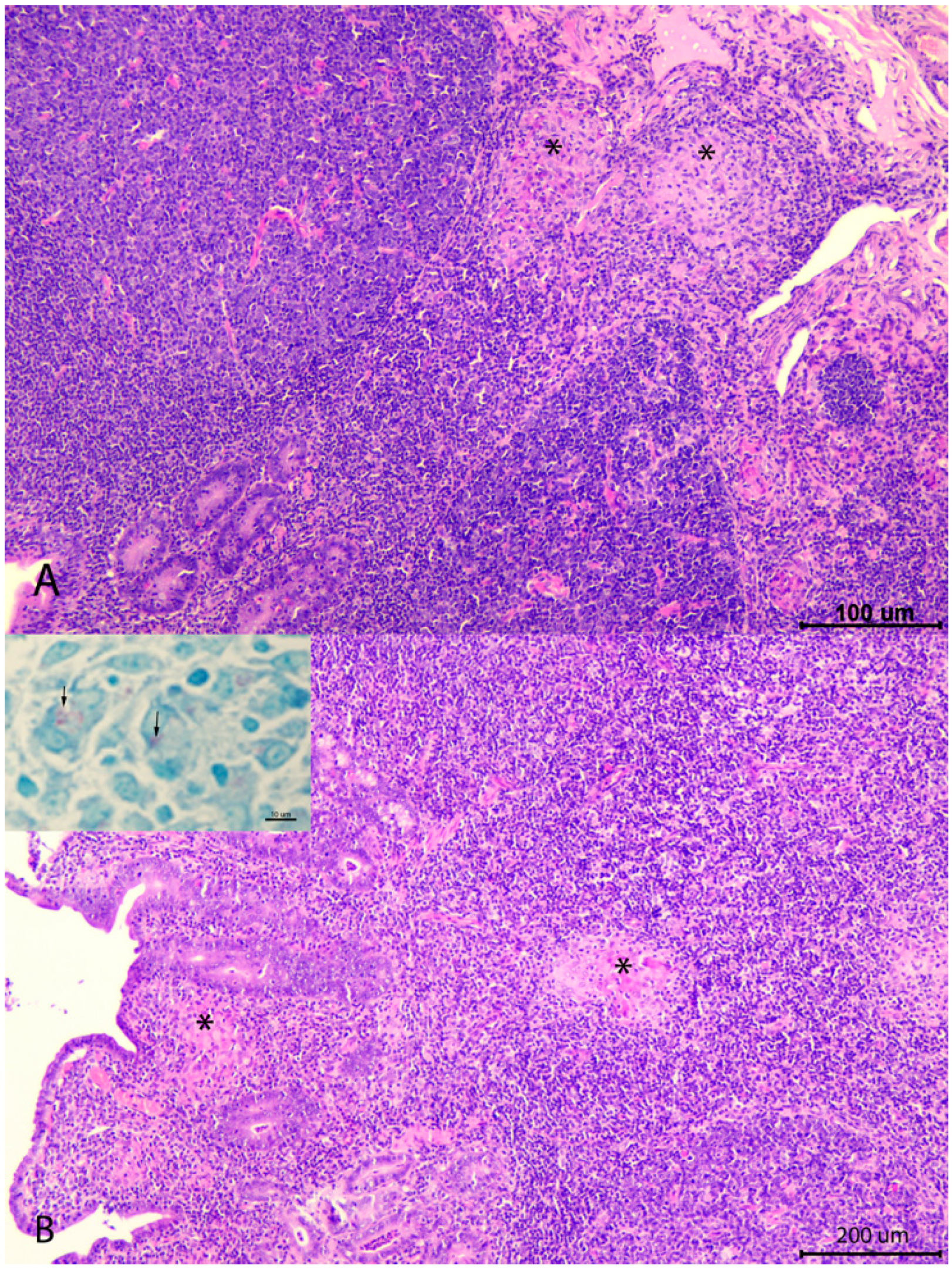
3.1. Evaluation of the Number of Losses
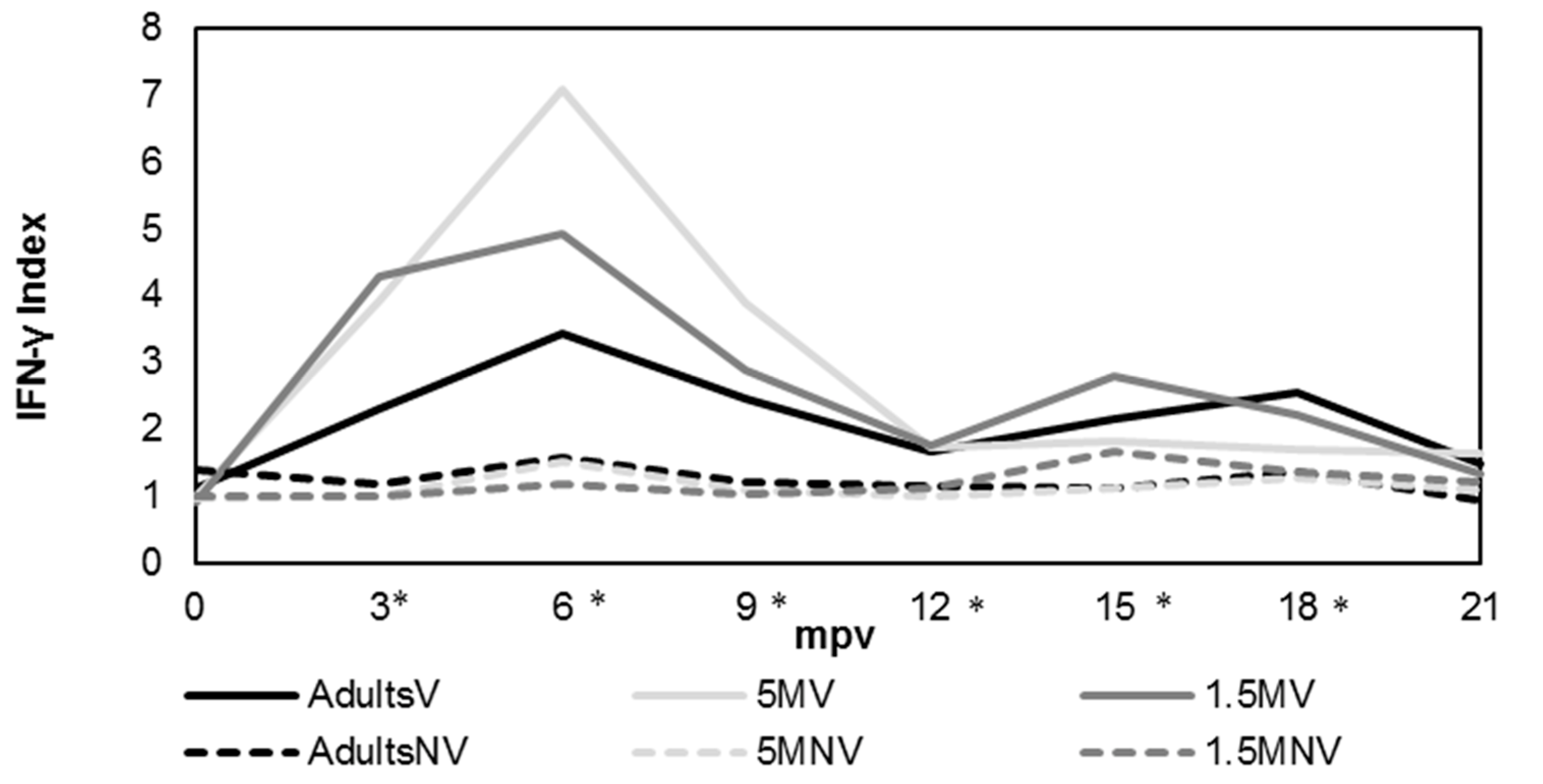
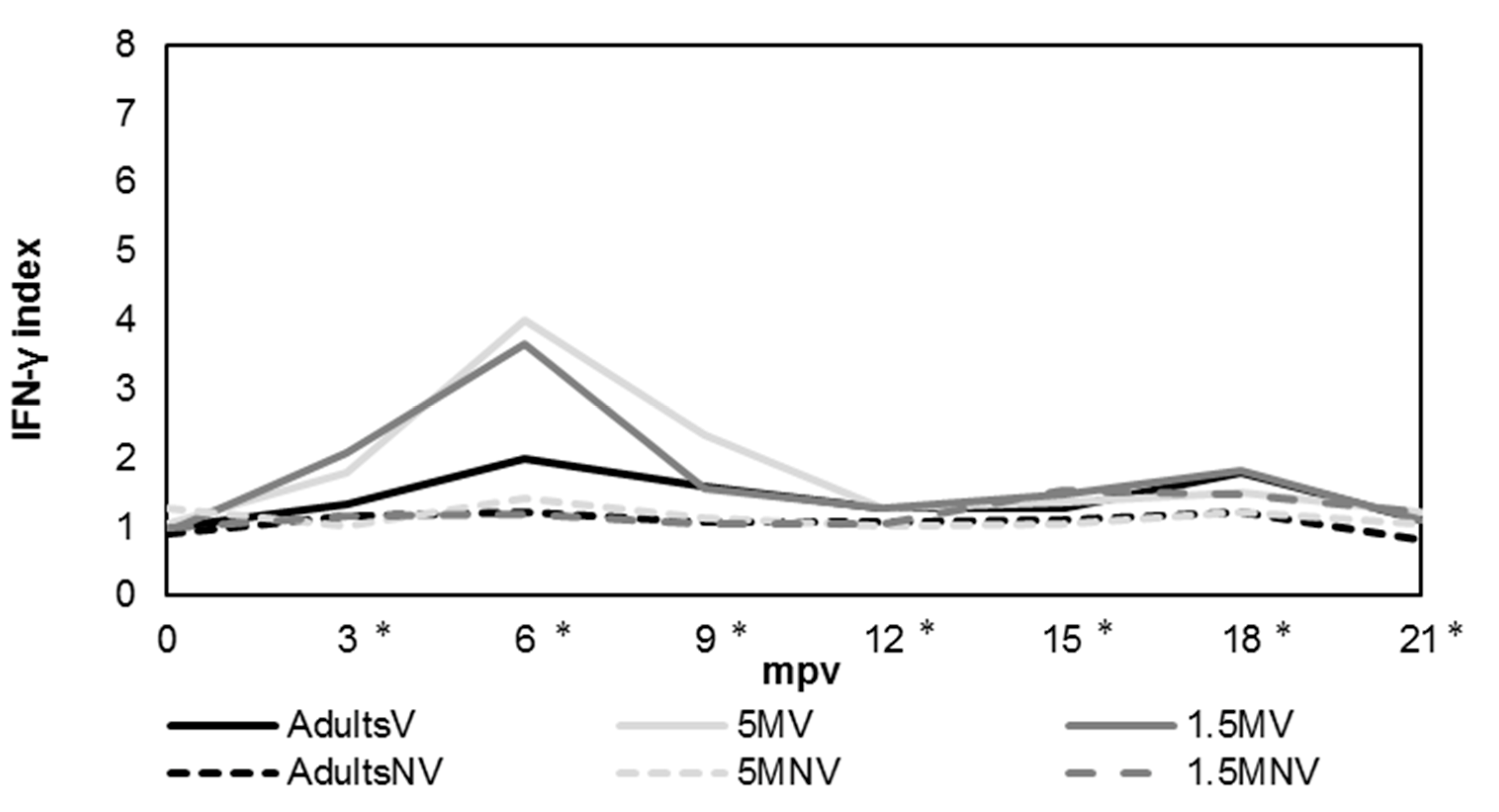
3.2. Cellular Immune Response Evaluation
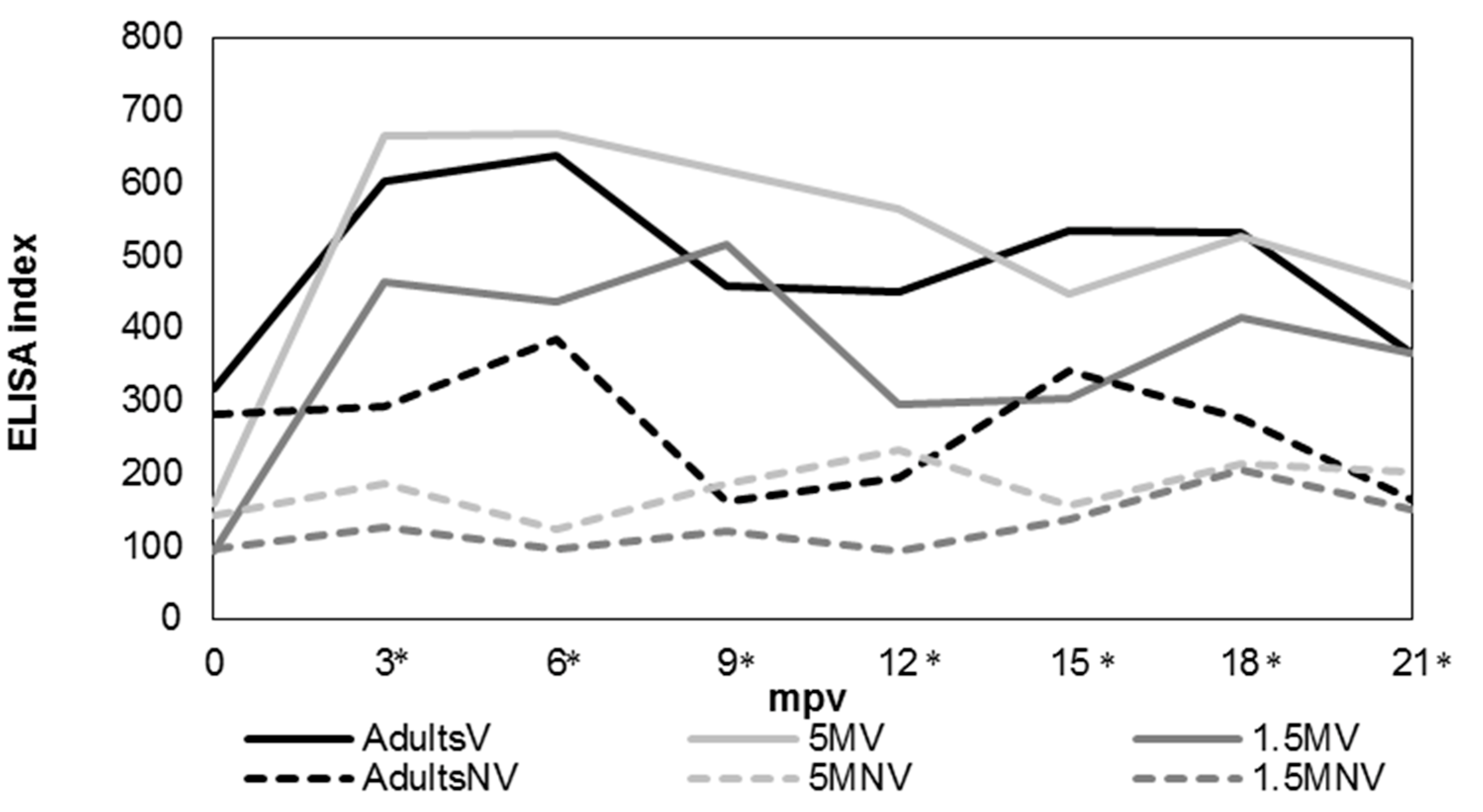
3.3. Humoral Immune Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wentink, G.; Bongers, J.; Zeeuwen, A.; Jaartsveld, F. Incidence of paratuberculosis after vaccination against M. paratuberculosis in two infected dairy herds. Zent. Vet. B 1994, 41, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Pérez, V.; García-Marín, J.; Bru, R.; Moreno, B.; Badiola, J. Resultados obtenidos en la vacunación de ovinos adultos frente a paratuberculosis. Med. Vet. 1995, 12, 196–201. [Google Scholar]
- Hosseiniporgham, S.; Rebechesu, L.; Pintore, P.; Lollai, S.; Dattena, M.; Russo, S.; Ruiu, A.; Sechi, L.A. A rapid phage assay for detection of viable Mycobacterium avium subsp. paratuberculosis in milk. Sci. Rep. 2022, 12, 475. [Google Scholar] [CrossRef] [PubMed]
- Foddai, A.; Strain, S.; Whitlock, R.H.; Elliott, C.T.; Grant, I.R. Application of a peptide-mediated magnetic separation-phage assay for detection of viable Mycobacterium avium subsp. paratuberculosis to bovine bulk tank milk and feces samples. J. Clin. Microbiol. 2011, 49, 2017–2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Hearn, M.; Molina, E.; Geijo, M.; Vazquez, P.; Sevilla, I.; Garrido, J.; Juste, R. Immunization of adult dairy cattle with a new heat-killed vaccine is associated with longer productive life prior to cows being sent to slaughter with suspected paratuberculosis. J. Dairy Sci. 2012, 95, 618–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Windsor, P. Managing control programs for ovine caseous lymphadenitis and paratuberculosis in Australia, and the need for persistent vaccination. Vet. Med. Res. Rep. 2014, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Juste, R.A.; Geijo, M.V.; Elguezabal, N.; Sevilla, I.A.; Alonso-Hearn, M.; Garrido, J.M. Paratuberculosis vaccination specific and non-specific effects on cattle lifespan. Vaccine 2021, 39, 1631–1641. [Google Scholar] [CrossRef]
- Mercier, P.; Brémaud, I.; Gautier, M.P. Vaccination of kids under one month of age with a killed vaccine and reduction in the frequency of faecal shedding of Mycobacterium avium subspecies paratuberculosis. Small Rumin. Res. 2014, 121, 425–433. [Google Scholar] [CrossRef]
- World Organization for Animal Health. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2022 of the World Organization for Animal Health Chapter 3.1.16. Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/3.01.16_PARATB.pdf (accessed on 23 August 2022).
- Corpa, J.; Pérez, V.; Sánchez, M.; García-Marín, J. Control of paratuberculosis (Johne’s disease) in goats by vaccination of adult animals. Vet. Rec. 2000, 146, 195–196. [Google Scholar] [CrossRef]
- Chiodini, R.; van Kruiningen, H.; Merkal, R. Ruminant paratuberculosis (Johne’s disease): The current status and future prospects. Cornell Vet. 1984, 74, 218–262. [Google Scholar]
- Köhler, H.; Gyra, H.; Zimmer, K.; Dräger, K.; Burkert, B.; Lemser, B.; Hausleithner, D.; Cubler, K.; Klawonn, W.; Hess, R. Immune reactions in cattle after immunization with a Mycobacterium paratuberculosis vaccine and implications for the diagnosis of M. paratuberculosis and M. bovis infections. J. Vet. Med. B Infect. Dis. Vet. Public Health 2001, 48, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Koets, A.; Ravesloot, L.; Ruuls, R.; Dinkla, A.; Eisenberg, S.; Lievaart-Peterson, K. Effects of age and environment on adaptive immune responses to Mycobacterium avium subsp. paratuberculosis (MAP) vaccination in dairy goats in relation to paratuberculosis control strategies. Vet. Sci. 2019, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Santema, W.; Rutten, V.; Segers, R.; Poot, J.; Hensen, S.; Heesterbeek, H.; Koets, A. Postexposure subunit vaccination against chronic enteric mycobacterial infection in a natural host. Infect. Immun. 2013, 81, 1990–1995. [Google Scholar] [CrossRef] [Green Version]
- Corpa, J.; Pérez, V.; García-Marín, J. Differences in the immune responses in lambs and kids vaccinated against paratuberculosis, according to the age of vaccination. Vet. Microbiol. 2000, 77, 475–485. [Google Scholar] [CrossRef]
- Benavides, J.; González, L.; Dagleish, M.; Pérez, V. Diagnostic pathology in microbial diseases of sheep and goats. Vet. Microbiol. 2015, 181, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Corpa, J.; Garrido, J.; García-Marín, J.; Pérez, V. Classification of lesions observed in natural cases of paratuberculosis in goats. J. Comp. Pathol. 2000, 122, 255–265. [Google Scholar] [CrossRef]
- Delgado, L.; Juste, R.; Muñoz, M.; Morales, S.; Benavides, J.; Ferreras, M.; García-Marín, J.; Pérez, V. Differences in the peripheral immune response between lambs and adult ewes experimentally infected with Mycobacterium avium subspecies paratuberculosis. Vet. Immunol. Immunopathol. 2012, 145, 23–31. [Google Scholar] [CrossRef]
- Dhand, N.; Eppleston, J.; Whittington, R.; Windsor, P. Changes in prevalence of ovine paratuberculosis following vaccination with Gudair®: Results of a longitudinal study conducted over a decade. Vaccine 2016, 34, 5107–5113. [Google Scholar] [CrossRef]
- Arteche-Villasol, N.; Gutiérrez-Expósito, D.; Elguezabal, N.; Sevilla, I.A.; Vallejo, R.; Espinosa, J.; Ferreras, M.; Benavides, J.; Pérez, V. Influence of heterologous and homologous vaccines and their components, on the host immune response and protection against experimental caprine paratuberuclosis. Front. Vet. Sci. 2022, 8, 744568. [Google Scholar] [CrossRef]
- Delgado, L.; García-Marín, J.; Muñoz, M.; Benavides, J.; Juste, R.; García-Pariente, C.; Fuertes, M.; González, J.; Ferreras, M.; Pérez, V. Pathological findings in young and adult sheep following experimental infection with 2 different doses of Mycobacterium avium subspecies paratuberculosis. Vet. Pathol. 2013, 50, 857–866. [Google Scholar] [CrossRef]
- Fernández, M.; Benavides, J.; Sevilla, I.; Fuertes, M.; Castaño, P.; Delgado, L.; García-Marín, J.; Garrido, J.; Ferreras, M.; Pérez, V. Experimental infection of lambs with C and S-type strains of Mycobacterium avium subspecies paratuberculosis: Immunological and pathological findings. Vet. Res. 2014, 45, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinnijenhuis, J.; van Crevel, R.; Netea, M. Trained immunity: Consequences for the heterologous effects of BCG vaccination. Trans. R Soc. Trop. Med. Hyg. 2015, 109, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Benn, C.; Joosten, L.; Jacobs, C.; van Loenhout, J.; Xavier, R.; Aaby, P.; van der Meer, J.; et al. Long-lasting effects of BCG vaccination on both heterologous Th1/Th17 responses and innate trained immunity. J. Innate Immun. 2014, 6, 152–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garly, M.L.; Martins, C.; Balé, C.; Baldé, M.; Hedegaard, K.; Gustafson, P.; Lisse, I.; Whittle, H.; Aaby, P. BCG scar and positive tuberculin reaction associated with reduced child mortality in West Africa. A non-specific beneficial effect of BCG? Vaccine 2003, 21, 2782–2790. [Google Scholar] [CrossRef]
- Vaugelade, J.; Pinchinat, S.; Guiella, G.; Elguero, E.; Simondon, F. Non-specific effects of vaccination on child survival: Prospective cohort study in Burkina Faso. BMJ 2004, 329, 1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niobey, F.; Duchiade, M.; Vasconcelos, A.; de Carvalho, M.; Leal, M.; Valente, J. Risk factors for death caused by pneumonia in children younger than 1 year old in a metropolitan region of southeastern Brazil. A case-Control study. Rev. Saude Publica 1992, 26, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.; Quintin, J.; van der Meer, J. Trained immunity: A memory for innate host defense. Cell Host Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.; Kleinnijenhuis, J.; Netea, M.; Dockrell, H. Whole blood profiling of Bacillus Calmette–Guérin-induced trained innate immunity in infants identifies Epidermal Growth Factor, IL-6, Platelet-Derived Growth Factor-AB/BB, and Natural Killer cell activation. Front. Immunol. 2017, 8, 644. [Google Scholar] [CrossRef] [Green Version]
- Gwozdz, J.; Thompson, K.; Manktelow, B.; Murray, A.; West, D. Vaccination against paratuberculosis of lambs already infected experimentally with Mycobacterium avium subspecies paratuberculosis. Aust. Vet. J. 2000, 78, 560–566. [Google Scholar] [CrossRef]
- Hines II, M.; Turnquist, S.; Ilha, M.; Rajeev, S.; Jones, A.; Whittington, L.; Bannantine, J.; Barletta, R.; Grohn, Y.; Katani, R.; et al. Evaluation of novel oral vaccine candidates and validation of a caprine model of Johne’s disease. Front. Cell Infect. Microbiol. 2014, 4, 26. [Google Scholar] [CrossRef]
- Chartier, C.; Mercier, P.; Pellet, M.; Vialard, J. Effect of an inactivated paratuberculosis vaccine on the intradermal testing of goats for tuberculosis. Vet. J. 2012, 191, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Chiodini, R. Immunology: Resistance to paratuberculosis. Vet. Clin. N. Am. Food Anim Pract 1996, 12, 313–343. [Google Scholar] [CrossRef]
- Begg, D.; Griffin, J. Vaccination of sheep against M. paratuberculosis: Immune parameters and protective efficacy. Vaccine 2005, 23, 4999–5008. [Google Scholar] [CrossRef]
- Koets, A.; Eda, S.; Sreevatsan, S. The within host dynamics of Mycobacterium avium ssp. paratuberculosis infection in cattle: Where time and place matter. Vet. Res. 2015, 46, 61. [Google Scholar] [CrossRef] [Green Version]
- Eppleston, J.; Reddacliff, L.; Windsor, P.; Links, I.; Whittington, R. Preliminary observations on the prevalence of sheep shedding Mycobacterium avium subsp paratuberculosis after 3 years of a vaccination program for ovine Johne’s disease. Aust. Vet. J. 2005, 83, 637–638. [Google Scholar] [CrossRef] [PubMed]
- Reddacliff, L.; Eppleston, J.; Windsor, P.; Whittington, R.; Jones, S. Efficacy of a killed vaccine for the control of paratuberculosis in Australian sheep flocks. Vet. Microbiol. 2006, 115, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Windsor, P.; Eppleston, J.; Dhand, N.; Whittington, R. Effectiveness of GudairTM vaccine for the control of ovine Johne’s disease in flocks vaccinating for at least 5 years. Aust. Vet. J. 2014, 92, 263–268. [Google Scholar] [CrossRef]
- Vázquez, P.; Garrido, J.; Juste, R. Specific antibody and interferon-gamma responses associated with immunopathological forms of bovine paratuberculosis in slaughtered Friesian cattle. PLoS ONE 2013, 8, e64568. [Google Scholar] [CrossRef]
- Fernández, M.; Fuertes, M.; Elguezabal, N.; Castaño, P.; Royo, M.; Ferreras, M.C.; Benavides, J.; Pérez, V. Immunohistochemical expression of interferon-γ in different types of granulomatous lesions associated with bovine paratuberculosis. Comp. Immunol. Microbiol. Infect. Dis. 2017, 51, 1–8. [Google Scholar] [CrossRef]
- Muskens, J.; van Zijderveld, F.; Eger, A.; Bakker, D. Evaluation of the long-term immune response in cattle after vaccination against paratuberculosis in two Dutch dairy herds. Vet. Microbiol. 2002, 86, 269–278. [Google Scholar] [CrossRef]
- Burrells, C.; Clarke, C.; Colston, A.; Kay, J.; Porter, J.; Little, D.; Sharp, J. A study of immunological responses of sheep clinically-affected with paratuberculosis (Johne’s disease). The relationship of blood, mesenteric lymph node and intestinal lymphocyte responses to gross and microscopic pathology. Vet. Immunol. Immunopathol. 1998, 66, 343–358. [Google Scholar] [CrossRef]
- Tanaka, S.; Sato, M.; Onitsuka, T.; Kamata, H.; Yokomizo, Y. Inflammatory cytokine gene expression in different types of granulomatous lesions during asymptomatic stages of bovine paratuberculosis. Vet. Pathol. 2005, 42, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Brady, C.; O’Grady, D.; O’Meara, F.; Egan, J.; Bassett, H. Relationships between clinical signs, pathological changes and tissue distribution of Mycobacterium avium subspecies paratuberculosis in 21 cows from herds affected by Johne’s disease. Vet. Rec. 2008, 162, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Pooley, H.B.; Begg, D.J.; Plain, K.M.; Whittington, R.J.; Purdie, A.C.; de Silva, K. The humoral immune response is essential for successful vaccine protection against paratuberculosis in sheep. BMC Vet. Res. 2019, 15, 223. [Google Scholar] [CrossRef] [Green Version]
- Hasvold, H.; Valheim, M.; Berntsen, G.; Storset, A. In vitro responses to purified protein derivate of caprine T lymphocytes following vaccination with live strains of Mycobacterium avium subsp. paratuberculosis. Vet. Immunol. Immunopathol. 2002, 90, 79–89. [Google Scholar] [CrossRef]
- Windsor, P. Research into vaccination against ovine Johne’s disease in Australia. Small Rumin. Res. 2006, 62, 139–142. [Google Scholar] [CrossRef]
- Nedrow, A.; Gavalchin, J.; Smith, M.; Stehman, S.; Maul, J.; McDonough, S.; Thonney, M. Antibody and skin-test responses of sheep vaccinated against Johne ’s disease. Vet. Immunol. Immunopathol. 2007, 116, 109–112. [Google Scholar] [CrossRef]
- Pérez de Val, B.; Nofrarías, M.; López-Soria, S.; Garrido, J.; Vordermeier, H.; Villarreal-Ramos, B.; Martín, M.; Puentes, E.; Juste, R.; Domingo, M. Effects of vaccination against paratuberculosis on tuberculosis in goats: Diagnostic interferences and cross-protection. BMC Vet. Res. 2012, 8, 191. [Google Scholar] [CrossRef] [Green Version]
- Garrido, J.; Vazquez, P.; Molina, E.; Plazaola, J.; Sevilla, I.; Geijo, M.; Alonso-Hearn, M.; Juste, R. Paratuberculosis vaccination causes only limited cross-reactivity in the skin test for diagnosis of bovine tuberculosis. PLoS ONE 2013, 8, 2–8. [Google Scholar] [CrossRef]
- Espinosa, J.; Fernández, M.; Royo, M.; Grau, A.; Collazos, J.A.; Benavides, J.; Ferreras, M.C.; Mínguez, O.; Pérez, V. Influence of vaccination against paratuberculosis on the diagnosis of caprine tuberculosis during official eradication programs in Castilla y León (Spain). Transbound. Emerg. Dis. 2021, 68, 692–703. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Number of Goats at Beginning | Number of Losses at Year 1 (%) | Number of Animals at Year 2 | Number of Losses at Year 2 (%) | Total Number of Losses (%) |
|---|---|---|---|---|---|
| AdultsV | 39 | 2 (5.13) | 37 | 1 (2.70) | 3 (7.69) |
| AdultsNV | 38 | 3 (7.89) | 35 | 4 (11.43) | 7 (18.42) |
| 5 MV | 40 | 2 (5.00) | 38 | 1 (2.63) | 3 (7.50) |
| 5 MNV | 39 | 1 (2.56) | 38 | 2 (5.26) | 3 (7.69) |
| 1.5 MV | 19 | 0 (0.00) | 19 | 0 (0.00) | 0 (0.00) |
| 1.5 MNV | 15 | 3 (20.00) | 12 | 0 (0.00) | 3 (20.00) |
| Total | 190 | 11 (5.79) | 179 | 8 (4.47) | 19 (19.00) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández, M.; Royo, M.; Fuertes, M.; Arteche-Villasol, N.; Ferreras, M.C.; Benavides, J.; Pérez, V. Effects of Paratuberculosis Vaccination at Different Ages in a Dairy Goat Herd: A 2-Year Follow-Up. Animals 2022, 12, 3135. https://doi.org/10.3390/ani12223135
Fernández M, Royo M, Fuertes M, Arteche-Villasol N, Ferreras MC, Benavides J, Pérez V. Effects of Paratuberculosis Vaccination at Different Ages in a Dairy Goat Herd: A 2-Year Follow-Up. Animals. 2022; 12(22):3135. https://doi.org/10.3390/ani12223135
Chicago/Turabian StyleFernández, Miguel, Marcos Royo, Miguel Fuertes, Noive Arteche-Villasol, M. Carmen Ferreras, Julio Benavides, and Valentín Pérez. 2022. "Effects of Paratuberculosis Vaccination at Different Ages in a Dairy Goat Herd: A 2-Year Follow-Up" Animals 12, no. 22: 3135. https://doi.org/10.3390/ani12223135