
Prevalence of Parasitic Infections with Zoonotic Potential in Tilapia: A Systematic Review and Meta-Analysis
 , , , ,
, , , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
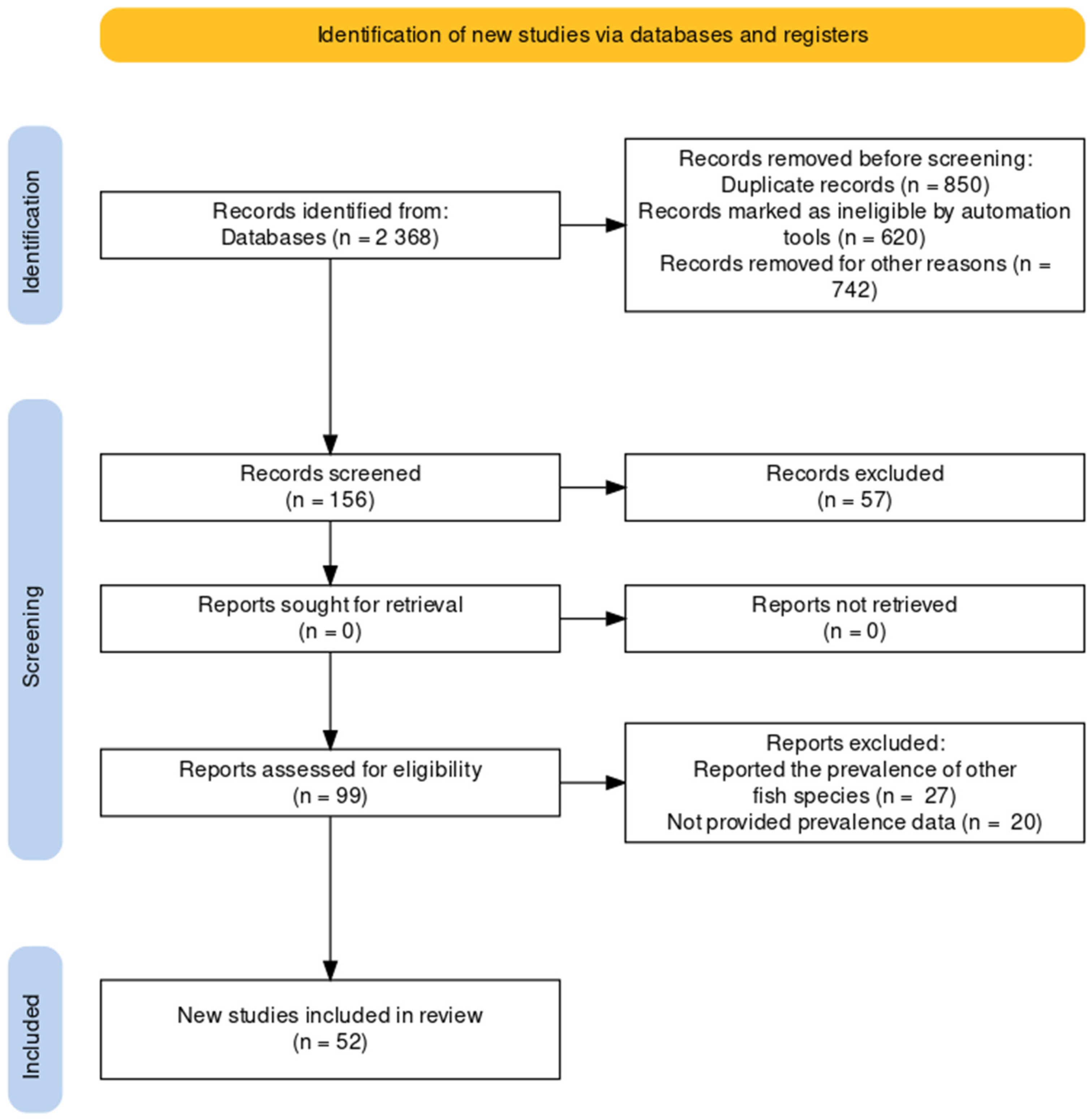
2.1. Data Sources and Searches
2.2. Study Selection and Eligibility Criteria
2.3. Data Extraction and Tabulation
2.4. Meta-Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Béné, C.; Arthur, R.; Norbury, H.; Allison, E.H.; Beveridge, M.; Bush, S.; Campling, L.; Leschen, W.; Little, D.; Squires, D.; et al. Contribution of Fisheries and Aquaculture to Food Security and Poverty Reduction: Assessing the Current Evidence. World Dev. 2016, 79, 177–196. [Google Scholar] [CrossRef]
- Food Agriculture Organization (FAO). El estado mundial de la Pesca y la Acuicultura. Mar. Pollut. Bull. 2020, 1, 165–171. [Google Scholar]
- Savaya, A.; Glassner, H.; Livne-Luzon, S.; Chishinski, R.; Molcho, J.; Aflalo, E.D.; Zilberg, D.; Sagi, A. Prawn monosex populations as biocontrol agents for snail vectors of fish parasites. Aquaculture 2020, 520, 1–36. [Google Scholar] [CrossRef]
- El-Sayed, A.-F.M. Taxonomy and Basic Biology. In Tilapia Culture, 2nd ed.; El-Sayed, A.-F.M., Ed.; Elsevier Inc. All.: Alexandria, Egypt, 2020; pp. 21–31. ISBN 978-0-12-816509-6. [Google Scholar] [CrossRef]
- Arguedas, D.; Ortega, C.; Martínez, S.; Astroza, A. Parasites of Nile Tilapia larvae Oreochromis niloticus (Pisces: Cichlidae) in concrete ponds in Guanacaste, Northern Costa Rica. UNED Res. J. 2017, 9, 313. [Google Scholar] [CrossRef]
- Fajer-Ávila, E.J.; Medina-Guerrero, R.M.; Morales-Serna, F.N. Strategies for prevention and control of parasite diseases in cultured tilapia. Acta Agrícola Pecu. 2017, 3, 25–31. [Google Scholar]
- Abdel-Latif, H.M.R.; Dawood, M.A.O.; Menanteau-Ledouble, S.; El-Matbouli, M. The nature and consequences of co-infections in tilapia: A review. J. Fish Dis. 2020, 43, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.; Cortés, M.; Zuñiga, L.; Jaime, B.; Galindo, J.; Basto, M. Small-scale culture of tilapia (Oreochromis niloticus), alimentary alternative for rural and peri-urban families in Mexico? Redvet 2010, 11, 177–185. [Google Scholar]
- Bao, M.; Pierce, G.J.; Strachan, N.J.C.; Pascual, S.; González-Muñoz, M.; Levsen, A. Human health, legislative and socioeconomic issues caused by the fish-borne zoonotic parasite Anisakis: Challenges in risk assessment. Trends Food Sci. Technol. 2019, 86, 298–310. [Google Scholar] [CrossRef]
- Carrique-Mas, J.J.; Bryant, J.E. A review of foodborne bacterial and parasitic zoonoses in Vietnam. Ecohealth 2013, 10, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Organización Mundial de la Salud (OMS). Zoonosis. 2022. p 1. Available online: https://www.who.int/es/news-room/fact-sheets/detail/zoonoses (accessed on 5 July 2022).
- Chibwana, F.D.; Mshana, J.G.; Katandukila, J.V. A Survey of Fish Parasites from Pangani Catchment and Lake Kitangiri in Singida, Tanzania. Tanzania J. Sci. 2020, 46, 42–52. [Google Scholar]
- Hung, N.M.; Dung, D.T.; Anh, N.T.L.; Van, P.T.; Thanh, B.N.; Van Ha, N.; Van Hien, H.; Canh, L.X. Current status of fish-borne zoonotic trematode infections in Gia Vien district, Ninh Binh province, Vietnam. Parasit Vectors 2015, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Young, N.; Korhonen, P.; Gasser, R. Clonorchis sinensis and Clonorchiasis: The Relevance of Exploring Genetic Variation. In Advances in Parasitology, 1st ed.; Elsevier Ltd.: London, UK, 2018; Volume 100, pp. 155–208. [Google Scholar] [CrossRef]
- Chi, T.T.; Dalsgaard, A.; Turnbull, J.; Tuan, P.; Murrell, D. Prevalence of zoonotic trematodes in fish from a vietnamese fish-farming community. J. Parasitol. 2008, 94, 423. [Google Scholar] [CrossRef]
- Ibrahim, M.M.; Soliman, M.F.M. Prevalence and site preferences of heterophyid metacercariae in Tilapia zilli from Ismalia fresh water canal. Egypt. Parasite 2010, 17, 233. [Google Scholar] [CrossRef] [Green Version]
- Wiriya, B.; Clausen, J.H.; Inpankaew, T.; Thaenkham, U.; Jittapalapong, S.; Satapornvanit, K.; Dalsgaard, A. Fish-borne trematodes in cultured Nile tilapia (Oreochromis niloticus) and wild-caught fish from Thailand. Vet. Parasitol. 2013, 198, 230–234. [Google Scholar] [CrossRef]
- Mardu, F.; Yohannes, M.; Tadesse, D. Prevalence of intestinal parasites and associated risk factors among inmates of Mekelle prison, Tigrai Region, Northern Ethiopia. BMC Infect. Dis. 2019, 19, 1. [Google Scholar] [CrossRef] [PubMed]
- Hop, N.T.; De, N.V.; Murrell, D.; Dalsgaard, A. Occurrence and species distribution of fishborne zoonotic trematodes in wastewater-fed aquaculture in northern Vietnam. Trop. Med. Int. Heal. 2007, 12 (Suppl. S2), 66. [Google Scholar] [CrossRef] [PubMed]
- Diaz Camacho, S.P.; Willms, K.; Ramos, M.; Del Carmen de la Cruz Otero, M.; Nawa, Y.; Akahane, H. Morphology of Gnathostoma spp. isolated from natural hosts in Sinaloa, Mexico. Parasitol. Res. 2002, 88, 639. [Google Scholar] [CrossRef] [PubMed]
- Motamedi, M.; Iranmanesh, A.; Teimori, A.; Sadjjadi, S.M.; Nasibi, S. Detection of Contracaecum multipapillatum (Nematoda: Anisakidae) in the indigenous killifish Aphanius hormuzensis (Teleostei; Aphaniidae) and its histopathological effects: A review of Iranian Aphanius species parasites. J. Appl. Ichthyol. 2019, 35, 558. [Google Scholar] [CrossRef]
- Salgado-Maldonado, G.; Aguilar-Aguilar, R.; Cabañas-Carranza, G.; Soto-Galera, E.; Mendoza-Palmero, C. Helminth parasites in freshwater fish from the Papaloapan river basin, Mexico. Parasitol. Res. 2005, 96, 69. [Google Scholar] [CrossRef]
- Ghoneim, N.H.; Abdel-Moein, K.A.; Saeed, H. Fish as a possible reservoir for zoonotic Giardia duodenalis assemblages. Parasitol. Res. 2012, 110, 2193. [Google Scholar] [CrossRef]
- Couso-Pérez, S.; Ares-Mazás, E.; Gómez-Couso, H. Identification of a novel piscine Cryptosporidium genotype and Cryptosporidium parvum in cultured rainbow trout (Oncorhynchus mykiss). Parasitol. Res. 2018, 117, 2987. [Google Scholar] [CrossRef]
- Koinari, M.; Karl, S.; Ng-Hublin, J.; Lymbery, A.J.; Ryan, U.M. Identification of novel and zoonotic Cryptosporidium species in fish from Papua New Guinea. Vet. Parasitol. 2013, 198, 1. [Google Scholar] [CrossRef]
- Milanez, G.D.; Masangkay, F.R.; Thomas, R.C.; Ordona, M.O.G.O.; Bernales, G.Q.; Corpuz, V.C.M.; Fortes, H.S.V.; Garcia, C.M.S.; Nicolas, L.C.; Nissapatorn, V. Molecular identification of Vermamoeba vermiformis from freshwater fish in lake Taal, Philippines. Exp. Parasitol. 2017, 183, 201. [Google Scholar] [CrossRef] [PubMed]
- Chelkha, N.; Hasni, I.; Louazani, A.C.; Levasseur, A.; La Scola, B.; Colson, P. Vermamoeba vermiformis CDC-19 draft genome sequence reveals considerable gene trafficking including with candidate phyla radiation and giant viruses. Sci. Rep. 2020, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Echi, P.; Okafor, F.; Eyo, J. Co-infection and morphometrics of three clinostomatids (Digenea: Clinostomatidae) in Tilapia guinensis Bleeker, 1862 from Opi lake, Nigeria. Bio Res. 2009, 7, 432. [Google Scholar] [CrossRef]
- Fleming, P.B.; Huffman, D.G.; Bonner, T.H.; Brandt, T.M. Metacercarial distribution of Centrocestus formosanus among fish hosts in the Guadalupe river drainage of Texas. J. Aquat. Anim. Health 2011, 23, 117. [Google Scholar] [CrossRef]
- Mahmoud, M.A.; Abdelsalam, M.; Mahdy, O.; El Miniawy, H.M.; Ahmed, Z.A.; Osman, A.H.; Mohamed, H.; Khattab, A.; Ewiss, M.Z. Infectious bacterial pathogens, parasites and pathological correlations of sewage pollution as an important threat to farmed fishes in Egypt. Environ. Pollut. 2016, 219, 939. [Google Scholar] [CrossRef]
- Morine, M.; Yang, R.; Ng, J.; Kueh, S.; Lymbery, A.J.; Ryan, U.M. Additional novel Cryptosporidium genotypes in ornamental fishes. Vet. Parasitol. 2012, 190, 578. [Google Scholar] [CrossRef] [Green Version]
- Dao, H.T.T.; Dermauw, V.; Gabriël, S.; Suwannatrai, A.; Tesana, S.; Nguyen, G.T.T.; Dorny, P. Opisthorchis viverrini infection in the snail and fish intermediate hosts in Central Vietnam. Acta Trop. 2017, 170, 120. [Google Scholar] [CrossRef] [PubMed]
- Elsheikha, H.M.; Elshazly, A.M. Host-dependent variations in the seasonal prevalence and intensity of heterophyid encysted metacercariae (Digenea: Heterophyidea) in brackish water fish in Egypt. Vet. Parasitol. 2008, 153, 65–72. [Google Scholar] [CrossRef]
- Elsheikha, H.M.; Elshazly, A.M. Preliminary observations on infection of brackish and fresh water fish by heterophyid encysted metacercariae in Egypt. Parasitol. Res. 2008, 103, 971. [Google Scholar] [CrossRef]
- Balduzzi, S.; Rücker, G.; Schwarzer, G. How to perform a meta-analysis with R: A practical tutorial. Evid. Based Ment. Health 2019, 22, 153. [Google Scholar] [CrossRef]
- Nyaga, V.N.; Arbyn, M.; Aerts, M. Metaprop: A Stata command to perform meta-analysis of binomial data. Arch. Public Health 2014, 72, 1. [Google Scholar] [CrossRef] [Green Version]
- Clopper, C.J.; Pearson, E.S. The Use of Confidence or Fiducial Limits Illustrated in the Case of the Binomial. Biometrika 1934, 26, 404–413. [Google Scholar] [CrossRef]
- Higgins, J.P.T.; Whitehead, A.; Simmonds, M. Sequential methods for random-effects meta-analysis. Stat. Med. 2011, 30, 903. [Google Scholar] [CrossRef] [Green Version]
- Sathish, S.; Chidambaram, P.; Uma, A. Prevalence of parasites in tilapia farms and their management practices in Tamil Nadu. India J. Entomol. Zool. Stud. 2021, 9, 678–689. [Google Scholar]
- Zhang, Y.; Gong, Q.; Lv, Q.; Qiu, Y.; Wang, Y.; Qiu, H.; Guo, X.; Gao, J.; Chang, Q.; Wang, C. Prevalence of Clonorchis sinensis infection in fish in South-East Asia: A systematic review and meta-analysis. J. Fish Dis. 2020, 43, 1409. [Google Scholar] [CrossRef]
- Gu, E.D.; Yu, D.F.; Yang, X.Y.; Xu, M.; Wei, H.; Luo, D.; Mu, D.X.; Hu, Y.C. Tilapia fisheries in Guangdong Province, China: Socio-economic benefits, and threats on native ecosystems and economics. Fish Manag. Ecol. 2019, 26, 97. [Google Scholar] [CrossRef]
- Rabie, G.; Ahlem, M.; Mehanna, S. Reproductive dynamics of the redbelly tilapia (Tilapia zillii gervais, 1848) in ayata lake as a ramsar site in south-eastern algeria. Egypt. J. Aquat. Biol. Fish. 2021, 25, 253. [Google Scholar] [CrossRef]
- Namaga, W.M.; Yahaya, B.; Salam, M.A. Proximate composition of male and female African catfish (Clarias gariepinus) and tilapia (Tilapia zilli) in Jega river, Kebbi state, Nigeria. J. Fish Lives Vet. Scien. 2020, 1, 28. [Google Scholar] [CrossRef]
- Guerrero, R.D.; Guerrero, L.A. Preliminary studies on the breeding and culture of the black-chin tilapia (Sarotherodon melanotheron) in freshwater ponds. Asia Life Sci. 2017, 26, 253–256. [Google Scholar]
- Sparks, J.S.; Stiassny, M.L.J. A new species of Ptychochromis from northeastern Madagascar (Teleostei: Cichlidae), with an update phylogeny and revised diagnosis for the genus. Zootaxa 2010, 2341, 33. [Google Scholar] [CrossRef]
- Pérez-Ponce De León, G.; Lagunas-Calvo, O.; García-Prieto, L.; Briosio-Aguilar, R.; Aguilar-Aguilar, R. Update on the distribution of the co-invasive Schyzocotyle acheilognathi (Bothriocephalus acheilognathi), the Asian fish tapeworm, in freshwater fishes of Mexico. J. Helminthol. 2018, 92, 279–290. [Google Scholar] [CrossRef]
- Ahmad, F.; Fazili, K.M.; Sofi, O.M.; Sheikh, B.A.; Sofi, T.A. Distribution and pathology caused by Bothriocephalus acheilognathi Yamaguti, 1934 (Cestoda: Bothriocephalidae): A review. Rev. Vet. 2018, 29, 142. [Google Scholar] [CrossRef]
- Molnár, K.; Buchmann, K.; Székely, C. Phylum Nematoda. In Fish Diseases and Disorders, 4th ed.; Elsevier Ltd.: London, UK, 2019; Volume 1, pp. 417–443. [Google Scholar] [CrossRef]
- León-Règagnon, V.; Osorio-Sarabia, D.; García-Prieto, L.; Lamothe-Argumedo, R.; Bertoni-Ruiz, F.; Oceguera-Figueroa, A. New host records of the nematode Gnathostoma sp. in Mexico. Parasitol. Int. 2005, 54, 51. [Google Scholar] [CrossRef]
- Leroy, J.; Cornu, M.; Deleplancque, A.S.; Loridant, S.; Dutoit, E.; Sendid, B. Sushi, ceviche and gnathostomiasis—A case report and review of imported infections. Travel Med. Infect Dis. 2017, 20, 26. [Google Scholar] [CrossRef]
- Macpherson, C.N.L. Human behaviour and the epidemiology of parasitic zoonoses. Int. J. Parasitol. 2005, 35, 1319. [Google Scholar] [CrossRef]
- Ogata, K.; Nawa, Y.; Akahane, H.; Camacho, S.D.; Lamothe-Argumedo, R.; Cruz-Reyes, A. Short Report: Gnathostomiasis In Mexico. Am. J. Trop. Med. Hyg. 1998, 58, 316. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.; Hernandez-Jover, M.; Shamsi, S. Fish substitutions which may increase human health risks from zoonotic seafood borne parasites: A review. Food Control. 2020, 118, 1. [Google Scholar] [CrossRef]
- Burton, B.; Clint, C.; Thomas, O. General Characteristics of the Trematoda. In Human Parasitology, 5th ed.; Elsevier Inc.: London, UK, 2019; Volume 1, pp. 149–174. [Google Scholar] [CrossRef]
- Kaleem, O.; Bio Singou Sabi, A.F. Overview of aquaculture systems in Egypt and Nigeria, prospects, potentials, and constraints. Aquac. Fish. 2020, 4, 1. [Google Scholar] [CrossRef]
- Squire, S.A.; Ryan, U. Cryptosporidium and Giardia in Africa: Current and future challenges. Parasites Vectors 2017, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- La Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO). World Food and Agriculture—Statistical Pocketbook World Food and Agriculture—Statistical Pocketbook 2018. Available online: https://www.fao.org/documents/card/es/c/ca6463en/ (accessed on 5 July 2022).
- Huang, S.; He, Y. Management of China’s capture fisheries: Review and prospect. Aquac. Fish. 2019, 4, 173. [Google Scholar] [CrossRef]
- Chai, J.Y.; Jung, B.K. Fishborne zoonotic heterophyid infections: An update. Food Waterborne Parasitol. 2017, 8–9, 33. [Google Scholar] [CrossRef]
- Webster, J.P.; Molyneux, D.H.; Hotez, P.J.; Fenwick, A. The contribution of mass drug administration to global health: Past, present and future. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 1. [Google Scholar] [CrossRef] [Green Version]
- Betson, M.; Alonte, A.J.; Ancog, R.C.; Aquino, A.M.; Belizario Jr, V.Y.; Bordado, A.M.; Clark, J.; Corales, M.C.; Dacuma, M.G.; Divina, B.P.; et al. Zoonotic transmission of intestinal helminths in southeast Asia: Implications for Control and Elimination. In Advances in Parasitology, 1st ed.; Elsevier Ltd.: London, UK, 2020; Volume 108, pp. 47–131. [Google Scholar] [CrossRef]
- Martínez Cruz, J.M.; Bravo Zamudio, R.; Aranda Patraca, A.; Martínez Marañón, R. La gnatostomiasis en méxico. Salud Publica Mex. 1989, 31, 541–549. [Google Scholar]
- McCarthy, J.; Moore, T.A. Emerging helminth zoonoses. Int. J. Parasitol. 2000, 30, 1351. [Google Scholar] [CrossRef]
- Diario Oficial de la Federación (DOF). NORMA Oficial Mexicana NOM-242-SSA1-2009, Productos y Servicios. Productos de la pesca frescos, refrigerados, congelados y procesados. Especificaciones sanitarias y métodos de prueba. Norma Of Mex. 2009. Available online: https://dof.gob.mx/normasOficiales/4295/salud2a/salud2a.htm (accessed on 5 July 2022).
- Tsiodras, S.; Kelesidis, T.; Kelesidis, I.; Bauchinger, U.; Falagas, M.E. Human infections associated with wild birds. J. Infect. 2008, 56, 83. [Google Scholar] [CrossRef]
- Adams, A.M.; Murrell, K.D.; Cross, J.H. Parasites of fish and risks to public health. Rev. Sci. Tech. 1997, 16, 652. [Google Scholar] [CrossRef]
- Pulido-Murillo, E.A.; Furtado, L.F.V.; Melo, A.L.; Rabelo, É.M.L.; Pinto, H.A. Fishborne zoonotic trematodes transmitted by Melanoides tuberculata snails, Peru. Emerg. Infect. Dis. 2018, 24, 606. [Google Scholar] [CrossRef] [Green Version]
- Vicente Pardo, J.M. Anisakis and Disease as an Occupational Disease. Med. Segur. Trab. 2016, 62, 223–240. [Google Scholar]
- Arafa, W.M.; Hassan, A.H.A.; Mahrous, L.N.; Abdel-Ghany, A.E.; Aboelhadid, S.M. Occurrence and molecular characterization of zoonotic Anisakis simplex sensu stricto and Anisakis pegreffii larvae in retail-marketed fish. J. Food Saf. 2019, 39, 1. [Google Scholar] [CrossRef]
- Herman, J.S. Gnathostomiasis Acquired by British Tourists in Botswana. Emerg. Infect. Dis. 2009, 15, 594. [Google Scholar] [CrossRef] [Green Version]
- Violante-González, J.; García-Varela, M.; Rojas-Herrera, A.; Guerrero, S.G. Diplostomiasis in cultured and wild tilapia Oreochromis niloticus in Guerrero State, Mexico. Parasitol. Res. 2009, 105, 803. [Google Scholar] [CrossRef]
- Behringer, D.C.; Karvonen, A.; Bojko, J. Parasite avoidance behaviours in aquatic environments. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 1. [Google Scholar] [CrossRef] [Green Version]
- Scholz, T.; Salgado-Maldonado, G. The introduction and dispersal of Centrocestus formosanus (Nishigori, 1924) (Digenea: Heterophyidae) in Mexico: A review. Am. Midl. Nat. 2000, 143, 185. [Google Scholar] [CrossRef]
- Umadevi, K.; Madhavi, R. The life cycle of Haplorchis pumilio (Trematoda: Heterophyidae) from the Indian region. J. Helminthol. 2006, 80, 327. [Google Scholar] [CrossRef]
- Amaechi, C.E. Prevalence, intensity and abundance of endoparasites in Oreochromis niloticus and Tilapia zilli (Pisces: Cichlidae) from Asa Dam, Ilorin, Nigeria. UNED Res. J. 2015, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Simon-Oke, I.A. Diversity, intensity and prevalence of parasites of cichlids in polluted and unpolluted sections of Eleyele Dam, Ibadan, Nigeria. UNED Res. J. 2017, 9, 45. [Google Scholar] [CrossRef]
- Yera, H.; Kuchta, R.; Brabec, J.; Peyron, F.; Dupouy-Camet, J. First identification of eggs of the Asian fish tapeworm Bothriocephalus acheilognathi (Cestoda: Bothriocephalidea) in human stool. Parasitol. Int. 2013, 62, 268–271. [Google Scholar] [CrossRef]
- Zhi, T.; Huang, C.; Sun, R.; Zheng, Y.; Chen, J.; Xu, X.; Brown, C.L.; Yang, T. Mucosal immune response of Nile tilapia Oreochromis niloticus during Gyrodactylus cichlidarum infection. Fish. Shellfish Immunol. 2020, 106, 21. [Google Scholar] [CrossRef]
- Papkou, A.; Gokhale, C.S.; Traulsen, A.; Schulenburg, H. Host-parasite coevolution: Why changing population size matters. Zoology 2016, 119, 330. [Google Scholar] [CrossRef]
- Mutengu, C.; Mhlanga, W.; Mupangwa, J.F. Occurrence of Clinostomum metacercariae in Oreochromis mossambicus from Mashoko Dam, Masvingo Province, Zimbabwe. Scientifica 2018, 2018, 1. [Google Scholar] [CrossRef] [Green Version]
- Cortés, D.A.; Dolz, G.; Zúñiga, J.J.R.; Rocha, A.E.J.; Alán, D.L. Centrocestus formosanus (Opisthorchiida: Heterophyidae) como causa de muerte de alevines de tilapia gris Oreochromis niloticus (Perciforme: Cichlidae) en el Pacífico seco de Costa Rica. Rev. Biol. Trop. 2010, 58, 1453. [Google Scholar] [CrossRef]
- Lun, Z.R.; Gasser, R.B.; Lai, D.H.; Li, A.X.; Zhu, X.Q.; Yu, X.B.; Fang, Y.Y. Clonorchiasis: A key foodborne zoonosis in China. Lancet Infect. Dis. 2005, 1, 31–41. [Google Scholar] [CrossRef]
- Dzikowski, R.; Diamant, A.; Paperna, I. Trematode metacercariae of fishes as sentinels for a changing limnological environment. Dis. Aquat. Organ. 2003, 55, 145. [Google Scholar] [CrossRef] [Green Version]
- Pinto, H.A.; Mati, V.L.T.; Melo, A.L. Metacercarial infection of wild nile tilapia (Oreochromis niloticus) from Brazil. Sci. World J. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Eissa, A.E.; Attia, M.M.; Elgendy, M.Y.; Ismail, G.A.; Sabry, N.M.; Prince, A. Streptococcus, Centrocestus formosanus and Myxobolus tilapiae concurrent infections in farmed Nile tilapia (Oreochromis niloticus). Microb. Pathog. 2021, 158, 105084. [Google Scholar] [CrossRef]
- Yimer, E. Preliminary survey of parasites and bacterial pathogens of fish at Lake Ziway. Ethiop. J. Sci. 2000, 23, 25. [Google Scholar] [CrossRef] [Green Version]
- Echi, P.C.; Eyo, J.E.; Okafor, F.C. Co-Parasitism And Morphometrics Of Three Clinostomatids (Digenea: Clinostomatidae) In Sarotherodon melanotheron From A Tropical Freshwater Lake. Anim. Res. Int. 2009, 6, 982. [Google Scholar] [CrossRef] [Green Version]
- Onyedineke, E.N.; Obi, U.; Ofoegbu, P.U.; Ukogo, I. Helminth Parasites of some Freshwater Fish from River Niger at Illushi, Edo State, Nigeria. J. Am. Sci. 2010, 6, 16–21. [Google Scholar]
- Roche, D.G.; Leung, B.; Mendoza Franco, E.F.; Torchin, M.E. Higher parasite richness, abundance and impact in native versus introduced cichlid fishes. Int. J. Parasitol. 2010, 40, 1525–1530. [Google Scholar] [CrossRef] [PubMed]
- Gulelat, Y.; Yimer, E.; Asmare, K.; Bekele, J. Study on parasitic helminths infecting three fish species from Koka reservoir, Ethiopia. SINET Ethiop. J. Sci. 2013, 36, 73–80. [Google Scholar]
- Amare, A.; Alemayehu, A.; Aylate, A. Prevalence of internal parasitic helminthes infected Oreochromis niloticus (Nile Tilapia), Clarias gariepinus (African Catfish) and Cyprinus carpio (Common Carp) in Lake Lugo (Hayke), Northeast Ethiopia. J. Aquac. Res. Dev. 2014, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Okoye, I.C.; Abu, S.J.; Obiezue, N.N.R.; Ofoezie, I.E. Prevalence and seasonality of parasites of fish in Agulu Lake, Southeast, Nigeria. Afr. J. Biotechnol. 2014, 13, 502. [Google Scholar] [CrossRef]
- Walakira, J.; Akoll, P.; Engole, M.; Sserwadda, M.; Nkambo, M.; Namulawa, V. Common fish diseases and parasites affecting wild and farmed tilapia and catfish in central and western Uganda. Uganda. J. Agric. Sci. 2014, 15, 113–125. [Google Scholar]
- Bekele, J.; Hussien, D. Prevalence of Internal Parasites of Oreochromis niloticus and Clarias gariepinus Fish Species in Lake Ziway, Ethiopia. J. Aquac. Res. Dev. 2015, 6, 10. [Google Scholar] [CrossRef]
- Reshid, M.; Adugna, M.; Tsegaye Redda, Y.; Awol, N.; Teklu, A. A Study of Clinostomum (Trematode) and Contracaecum (Nematode) Parasites Affecting Oreochromis Niloticus in Small Abaya Lake, Silite Zone, Ethiopia. J. Aquac. Res Dev. 2015, 6, 316. [Google Scholar] [CrossRef] [Green Version]
- Sèdogbo, M.H.; Zannou, B.T.; Siko, J.E.; Tossavi, N.D.; Togla, I.; Fiogbé, E.D. Faune des métazoaires parasites de Clarias gariepinus (Clariidae) et de Oreochromis niloticus (Cichlidae), deux poissons des whédos du delta supérieur du fleuve Ouémé au sud du Bénin. Int. J. Biol. Chem. Sci. 2019, 13, 984–997. [Google Scholar] [CrossRef] [Green Version]
- Adugna, M.; Fishery, N.; Life, A. The Prevalence of Fish Parasites of Nile Tilapia (Oreochromis niloticus) in Selected Fish farms, Amhara Regional State. Ethiop. J. Agric. Sci. 2020, 30, 119–128. [Google Scholar]
- Otachi, E.; Magana, A.; Jirsa, F.; Fellner-Frank, C. Parasites of commercially important fish from Lake Naivasha, Rift Valley, Kenya. Parasitol. Res. 2014, 113, 1057. [Google Scholar] [CrossRef] [PubMed]
- Otachi, E.; Szostakowska, B.; Jirsa, F.; Fellner-Frank, C. Parasite communities of the elongate tigerfish Hydrocynus forskahlii (Cuvier 1819) and redbelly tilapia Tilapia zillii (Gervais 1848) from Lake Turkana, Kenya: Influence of host sex and size. Acta Parasitol. 2015, 60, 9. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Olvera, L.; Benavides-González, F.; Rábago-Castro, J.L.; Pérez-Castañeda, R.; García-Prieto, L. Endohelminths of Fishes of Commercial Importance from Vicente Guerrero Reservoir, Tamaulipas, Mexico. Comp. Parasitol. 2017, 84, 194. [Google Scholar] [CrossRef]
- Davidovich, N.; Tedesco, P.; Caffara, M.; Yasur-Landau, D.; Gustinelli, A.; Drabkin, V. Morphological description and molecular characterization of Contracaecum larvae (Nematoda: Anisakidae) parasitizing market-size hybrid tilapia (Oreochromis aureus x Oreochromis niloticus) and red drum (Sciaenops ocellatus) farmed in Israel. Food Waterborne Parasitol. 2022, 26, e00147. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Hedegaard Clausen, J.; Murrell, D.; Liu, L.; Dalsgaard, A. Risks for fishborne zoonotic trematodes in Tilapia production systems in Guangdong province, China. Vet. Parasitol. 2013, 198, 223–229. [Google Scholar] [CrossRef]
- Wilson, J.R.; Saunders, R.J.; Hutson, K.S. Parasites of the invasive tilapia Oreochromis mossambicus: Evidence for co-introduction. Aquat. Invasions 2019, 14, 332. [Google Scholar] [CrossRef]
- Mosqueda Cabrera, M.Á.; Miranda, E.S.; Calderón, L.C.; Ortiz Nájera, H.E. Finding advanced third-stage larvae of Gnathostoma turgidum Stossich, 1902 in Mexico from natural and experimental host and contributions to the life cycle description. Parasitol. Res. 2009, 104, 1219. [Google Scholar] [CrossRef]
- Thien, P.; Dalsgaard, A.; Thanh, B.N.; Olsen, A.; Murrell, K.D. Prevalence of fishborne zoonotic parasites in important cultured fish species in the Mekong Delta, Vietnam. Parasitol. Res. 2007, 101, 1277. [Google Scholar] [CrossRef]
- Díaz, M.T.; Hernandez, L.E.; Bashirullah, A.K. Studies on the life cycle of Haplorchis pumilio (Looss, 1896) (Trematoda: Heterophyidae) in Venezuela. Rev. Cient. la Fac. Ciencias Vet la Univ. del Zulia 2008, 18, 35–42. [Google Scholar]
- Thien, P.; Dalsgaard, A.; Thanh Nhan, N.; Olsen, A.; Murrell, K.D. Prevalence of zoonotic trematode parasites in fish fry and juveniles in fish farms of the Mekong Delta, Vietnam. Aquaculture 2009, 295, 1–5. [Google Scholar] [CrossRef]
- Chi, T.T.; Murrell, K.D.; Mausen, H.; Khue, N.V.; Dalsgaard, A. Fishborne zoonotic trematodes in raw fish dishes served in restaurants in nam dinh province and hanoi, vietnam. J. Food Prot. 2009, 72, 2394. [Google Scholar] [CrossRef] [PubMed]
- Lobna, S.M.A.; Metawea, Y.F.; Elsheikha, H.M. Prevalence of heterophyiosis in Tilapia fish and humans in Northern Egypt. Parasitol Res. 2010, 107, 1029. [Google Scholar] [CrossRef]
- Thi Phan, V.; Kjær Ersbøll, A.; Quang Bui, T.; Thi Nguyen, H.; Murrell, D.; Dalsgaard, A. Fish-Borne Zoonotic Trematodes in Cultured and Wild-Caught Freshwater Fish from the Red River Delta, Vietnam. Vector-Borne Zoonotic Dis. 2010, 10, 861–866. [Google Scholar] [CrossRef]
- Thi Phan, V.; Kjær Ersbøll, A.; Nguyen, K.V.; Madsen, H.; Dalsgaard, A. Farm-level risk factors for Fish-borne zoonotic trematode infection in integrated Small-scale fish farms in Northern Vietnam. PLoS Negl. Trop. Dis. 2010, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Van De, N.; Le, T.H.; Murrell, K.D. Prevalence and intensity of fish-borne zoonotic trematodes in cultured freshwater fish from rural and Urban Areas of Northern Vietnam. J. Parasitol. 2012, 98, 1023. [Google Scholar] [CrossRef] [PubMed]
- Hegazi, M.A.; Abo-elkheir, O.I. Encysted Metacercariae of Family Heterophyidae in Infected Fish in Dakahlia Governorate, an Endemic Focus in Egypt. J. Egypt. Soc. Parasitol. 2014, 12, 547–558. [Google Scholar] [CrossRef]
- Kopolrat, K.; Sithithaworn, P. Susceptibility, metacercarial burden, and mortality of juvenile silver barb, common carp, mrigal, and tilapia following exposure to Haplorchis taichui. Parasitol. Res. 2015, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Madsen, H.; Dung, B.T.; The, D.T.; Viet, N.K.; Dalsgaard, A.; Van, P.T. The role of rice fields, fish ponds and water canals for transmission of fish-borne zoonotic trematodes in aquaculture ponds in Nam Dinh Province, Vietnam. Parasites Vectors 2015, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojwala, R.A.; Otachi, E.O.; Kitaka, N.K. Effect of water quality on the parasite assemblages infecting Nile tilapia in selected fish farms in Nakuru County, Kenya. Parasitol. Res. 2018, 117, 3459–3471. [Google Scholar] [CrossRef]
- Scholz, T.; Šimková, A.; Razanabolana, J.R.; Kuchta, R. The first record of the invasive Asian fish tapeworm (Schyzocotyle acheilognathi) from an endemic cichlid fish in Madagascar. Helminthologia 2018, 55, 84. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | (Oreochromis or Tilapia 1 or Cichlids) and (Parasite or Nematode or Cestode or Trematode) |
| 2 | zoonotic parasite/or foodborne parasites/or Oreochromis/or Tilapia 1/ |
| 3 | prevalence/or Diphyllobothrium/or Diphyllobothriasis/or Oreochromis/or Tilapia 1/ |
| 4 | prevalence/or Bothriocephalus or Schyzocotyle/or Bothriocephaliasis/or Oreochromis/or Tilapia 1/ |
| 5 | prevalence/or Centrocestus/or Centrocestiasis/or Oreochromis/or Tilapia 1/ |
| 6 | prevalence/or Clonorchis/or Clonorchiasis/or Oreochromis/or Tilapia 1/ |
| 7 | prevalence/or Opisthorchis/or Opisthorchiasis/or Oreochromis/or Tilapia 1/ |
| 8 | prevalence/or Heterophyes/or Heterofiasis/or Oreochromis/or Tilapia 1/ |
| 9 | prevalence/or Haplorchis/or Haplorchiasis/or Oreochromis/or Tilapia 1/ |
| 10 | prevalence/or Gnathostoma/or Gnathostomiasis/or Oreochromis/or Tilapia 1/ |
| Primary Outcomes |
|---|
| Number of events per study (positive cases) |
| Size of study population |
| Prevalence |
| Exploratory outcomes |
| Host fish (genera) |
| Parasite (genera and species) |
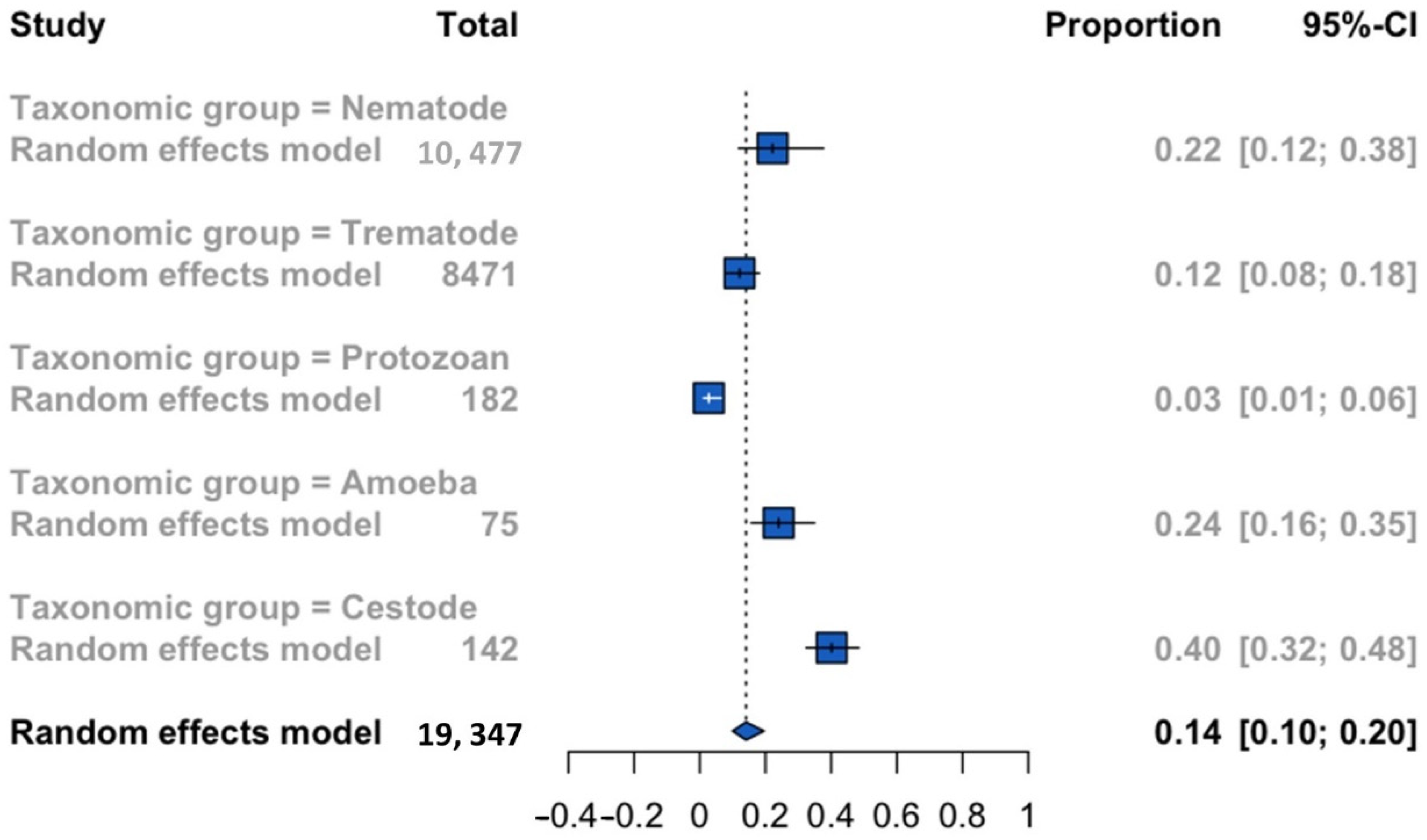
| Parasite taxonomic group (nematode, trematode, protozoan, amoeba independently tested for their free-living in ponds, and cestode) |
| Sample origin (aquaculture, fishing, restaurants, and aquaculture-fishing) |
| Diagnostic test (microscopy and PCR) |
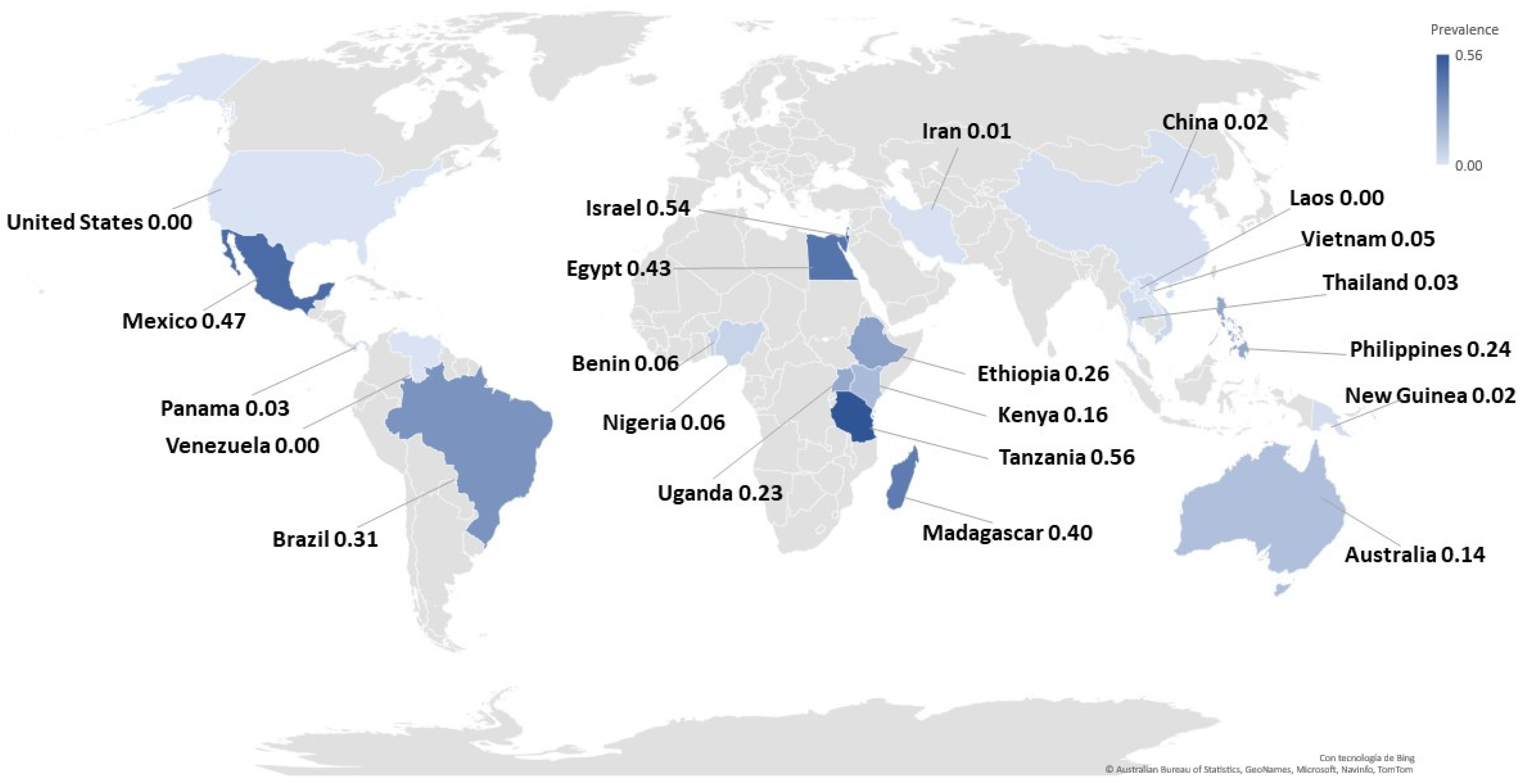
| Country of study |
| Continent of study |
| Item | Summary Proportion | Lower 95% CI | Upper 95% CI | I2 | Sample Size |
|---|---|---|---|---|---|
| Overall estimated | 0.14 | 0.10 | 0.20 | 98.4 | 19,347 |
| Host | |||||
| Oreochromis | 0.09 | 0.06 | 0.15 | 99.0 | 14,379 |
| Tilapia | 0.19 | 0.10 | 0.34 | 98.0 | 3923 |
| Sarotherodon | 0.42 | 0.22 | 0.65 | 98.0 | 863 |
| Ptychochromis | 0.40 | 0.32 | 0.48 | n.e. | 142 |
| Vieja | 0.02 | 0.003 | 0.15 | n.e. | 40 |
| p-value * | <0.0001 | ||||
| Taxonomic group | |||||
| Amoeba | 0.24 | 0.16 | 0.35 | n.e. | 75 |
| Cestode | 0.40 | 0.32 | 0.48 | n.e. | 142 |
| Nematode | 0.22 | 0.11 | 0.38 | 99.4 | 10,477 |
| Protozoan | 0.03 | 0.01 | 0.06 | 0.0 | 182 |
| Trematode | 0.12 | 0.08 | 0.18 | 96.3 | 8471 |
| p-value * | <0.0001 | ||||
| Continent | |||||
| Africa | 0.28 | 0.20 | 0.37 | 98.1 | 6420 |
| America | 0.13 | 0.00 | 0.41 | 99.8 | 8409 |
| Asia | 0.10 | 0.04 | 0.18 | 99.0 | 4356 |
| Oceania | 0.07 | 0.00 | 0.22 | 97.5 | 162 |
| p-value * | <0.0004 | ||||
| Sample origin | |||||
| Aquaculture | 0.05 | 0.02 | 0.11 | 92.6 | 2872 |
| Fishing | 0.24 | 0.17 | 0.33 | 98.8 | 15,615 |
| Restaurants | 0.15 | 0.08 | 0.26 | n.e. | 65 |
| Aquaculture-fishing | 0.05 | 0.03 | 0.11 | 86.0 | 795 |
| p-value * | <0.0001 | ||||
| Diagnostic test | |||||
| Microscopy | 0.14 | 0.09 | 0.20 | 98.2 | 14,476 |
| PCR | 0.13 | 0.07 | 0.24 | 96.6 | 4871 |
| p-value * | 0.97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acosta-Pérez, V.J.; Ángeles-Hernández, J.C.; Vega-Sánchez, V.; Zepeda-Velázquez, A.P.; Añorve-Morga, J.; Ponce-Noguez, J.B.; Reyes-Rodríguez, N.E.; De-La-Rosa-Arana, J.L.; Ramírez-Paredes, J.G.; Gómez-De-Anda, F.R. Prevalence of Parasitic Infections with Zoonotic Potential in Tilapia: A Systematic Review and Meta-Analysis. Animals 2022, 12, 2800. https://doi.org/10.3390/ani12202800
Acosta-Pérez VJ, Ángeles-Hernández JC, Vega-Sánchez V, Zepeda-Velázquez AP, Añorve-Morga J, Ponce-Noguez JB, Reyes-Rodríguez NE, De-La-Rosa-Arana JL, Ramírez-Paredes JG, Gómez-De-Anda FR. Prevalence of Parasitic Infections with Zoonotic Potential in Tilapia: A Systematic Review and Meta-Analysis. Animals. 2022; 12(20):2800. https://doi.org/10.3390/ani12202800
Chicago/Turabian StyleAcosta-Pérez, Víctor Johan, Juan Carlos Ángeles-Hernández, Vicente Vega-Sánchez, Andrea Paloma Zepeda-Velázquez, Javier Añorve-Morga, Jesús Benjamín Ponce-Noguez, Nydia Edith Reyes-Rodríguez, Jorge Luis De-La-Rosa-Arana, José Gustavo Ramírez-Paredes, and Fabián Ricardo Gómez-De-Anda. 2022. "Prevalence of Parasitic Infections with Zoonotic Potential in Tilapia: A Systematic Review and Meta-Analysis" Animals 12, no. 20: 2800. https://doi.org/10.3390/ani12202800