1. Introduction
Aquatic products are rich in high-quality protein, EPA, and DHA, which have beneficial effects on human pathologies [
1]. The world population is expected to cross 9.7 billion by the end of 2050 [
2]. An unprecedented food crisis will emerge [
3]. A recent study has predicted that the aquaculture industry will play a critical role in fulfilling the fast-growing demand for animal protein [
4]. In the meantime, the growth of aquatic animals needs a high amount of dietary protein intake [
5]. Thus, the supply of protein to aquatic feeds is important to the sustainable development of the aquaculture industry. Currently, fish meal (FM) is the major source of protein in aquatic feeds. With balanced amino acid composition, good palatability, and abundant biologically active compounds, FMs have high nutritional value and digestibility for aquatic animals [
6,
7]. However, with the rapid development of the aquaculture industry, the increasing FM demand put pressure on marine resources [
8,
9]. In recent years, the shortage of FM is now a big obstacle to having a sustainable aquaculture industry worldwide. Hence, we pay much more attention to replacing FM with novel protein sources [
10].
Some animal and plant protein sources are used to replace FM [
11]. Animal protein sources are more widely accepted by fish for their more balanced amino acid profiles [
12,
13]. Especially, insect protein is a type of animal protein that is environmentally and economically sustainable [
14]. At the same time, insects have low greenhouse gas and nitrogen emissions during the rearing process and cause less pollution to the environment [
15]. However, the lack of relevant research has hindered the promotion and application of insect protein in aquafeeds. At present, the production scale of domestic insect protein is not large, and the price is slightly higher [
16]. Thus, many insect proteins have been applied to the study of aquatic feeds, such as black soldier fly (
Hermetia illucens) larvae meal [
17], housefly maggot (
Musca domestica) meal [
18], and field cricket (
Gryllus bimaculatus) meal [
19].
Currently, Yellow mealworm (
Tenebrio molitor) meal (TM) is one of the novel insect protein sources which is easily available and in which the culture conditions are simple [
20]. The defatted TM has a high crude protein content of up to 63.84%, and its amino acid profile is similar to FM (
Table 1). Furthermore, previous studies have indicated that TM contained biologically active substances with anti-tumor, anti-bacterial, antioxidant, and immunomodulatory functions [
21,
22]. Studies had shown that replacing 50% of FM with defatted TM improved the growth and immunity function of Pacific white shrimp (
Litopenaeus vannamei) [
23]. Diets supplemented with defatted TM improve the survival rate of red seabream (
Pargus major) [
24]. Additionally, defatted TM can replace up to 50% of FM in the diet of largemouth bass (
Micropterus salmoides) without an effect on growth performance [
25]. Thus, defatted TM is likely a high-quality protein source in aquatic feeds.
Large yellow croaker (
Larimichthys crocea) is the most important marine fish in China, with high nutritional value, delicious taste, and important commercial value [
26]. In total, 47% crude protein and 10.5% crude lipid can meet the nutritional requirements of large yellow croakers, which means that a large amount of protein is needed in feed [
27]. At present, a high proportion of FM is abundantly used in feed for large yellow croakers, which is highly uneconomical and unsustainable [
28]. Therefore, it is imperative to find a suitable replacement for FM and develop a sustainable, efficient, and environmentally friendly feed. Currently, studies related to the replacement of large yellow croaker FM by protein sources such as fermented soybean [
29] and soy protein concentrate meal [
30] have been reported. However, as a carnivorous fish, animal-source protein has more advantages for large yellow croakers. Furthermore, juvenile large yellow croakers have incompletely differentiated and underdeveloped intestines [
31], and the production of large yellow croakers is highly susceptible to high juvenile mortality [
32]. Some traditional animal sources of protein used to replace FM result in intestine injury and poor growth performance. Here, the present study selected TM to replace FM in large yellow croaker diets. The effect of TM replacement FM on the growth performance and intestinal health of juvenile large yellow croakers was evaluated.
2. Materials and Method
2.1. Experimental Diets
Four groups of iso-nitrogenous and iso-lipidic diets were formulated with 0%, 6.7%, 13.3% and 20.0% TM protein content to substitute 0% (control group, FM content: 40%), 15%, 30%, and 45% protein content in fish meal (
Table 2). All ingredients were crushed, passed through a 180 µm mesh sieve, then accurately weighed according to the feed formulation and produced as 2 mm puffed pellets using a TES65 twin-screw extruder (Modern Yanggong Machinery Technology Development Co., Ltd., Beijing, China). After drying at 45 °C for 12 h, the mixed fish oil and soybean oil were evenly sprayed onto the surface of the pellets, and all diets were sealed and stored at −20 °C.
2.2. Fish and Feeding Trial
This study was conducted in accordance with the Experimental Animal Ethics Committee of the Fisheries College, Jimei University, with the approval number 2011-58. Juvenile large yellow croakers were obtained from a local hatchery (Ningde, China). Before the feeding trial, juvenile fish were temporarily cultured in floating sea cages (4.0 m × 4.0 m × 4.0 m) to acclimate to the experimental condition for 14 days. After the adaptation period, 1800 healthy fish with similar size (11.80 ± 0.02 g) were randomly selected and assigned to 12 floating cages (2.0 m × 2.0 m × 2.0 m) with 150 fish per cage. All experimental fish were significantly satiated by hand feeding twice daily (6:00 a.m. and 6:00 p.m.) using the four experimental diets. Satiation is considered to be reached when most fish stop eating. After each meal, the residual feed in each floating sea cage was collected, respectively, dried, and weighed to exclude the feed intake amount. The feeding trial lasted 56 days. During the feeding trial, the water temperature, pH, dissolved oxygen, and salinity ranged as follows: 20.4–25.8 °C, 8.16–8.65, 6.11–7.97 mg/L, and 27.3–27.9‰.
2.3. Sample Collection
Before sampling, all fish were fasted for 24 h and anesthetized with eugenol (1:10,000). The total number and weight of fish in each cage were counted to calculate the survival rate (SR) and weight gain (WG). Twenty fish were randomly selected from each cage. Blood was drawn from the tail vein using a sterile injector, and serum was separated by centrifuging (850× g, 10 min at 4 °C) and then stored at −80 °C until analysis. The liver and gut were collected for histological and other analyses.
2.4. Analytical Methods
2.4.1. Proximate Composition
The proximate compositions of feed and fish were measured according to the AOAC methods [
33]. The moisture was analyzed by drying continuously at 105 °C. The crude protein was determined using the Kjeldahl method after acid digestion. The crude lipid was determined by Soxhlet ether extraction, and the ash content was determined by scorching in a muffle furnace at 550 °C for 8 h. The phosphorus (P) concentration in experimental diets was quantified by inductively coupled plasma atomic emission spectrometry (ICP-OES, Prodigy7, Leeman, Hudson, NH, USA). The samples of feed and protein materials were lyophilized to constant weight, and 30 mg of each sample was taken and placed in 15 mL 6N HCl solution and hydrolyzed at 110 °C for 24 h. Determining amino acid profiles was performed with an automated amino acid analyzer (L-8900, Hitachi, Tokyo, Japan).
2.4.2. Digestibility Enzyme
Intestine tissues were homogenized in the pre-cooled normal saline and the homogenate was collected by centrifuging at 3000×
g for 10 min. Intestinal lipase and trypsin activities were measured by competition method using commercial kits (Nanjing Jiancheng Biological Company, Nanjing, China) according to the manufacturer’s instructions. Quantification of proteins was performed with a total protein quantitative kit (Beijing Solarbio Science and Technology Company, Beijing, China) according to our previous study [
34].
2.4.3. Serum Biochemical Parameters
Serum complement 4 (C4) levels, complement 3 (C3) levels, and D-lactate concentration were determined by the competition method according to previous reports [
35] using ELISA kits for the fish (Nanjing Jiancheng Biological Company, Nanjing, China). The determination of serum diamine oxidase (DAO), alkaline phosphatase (AKP), and lysozyme (LZM) activities according to the colorimetric method was performed with commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). One LZM activity unit was to be defined as the amount of enzyme required to decrease absorptivity at 37 °C at a rate of 0.001 min/mL, and one AKP activity unit was defined as the production of 1 mg of phenol by the action of serum with the substrate for 15 min at 37 °C, and one DAO activity unit was defined as the formation of 1 mmol of ammonia per minute per milliliter of serum at 37 °C.
2.4.4. Intestinal Histology
Gut samples were stained with hematoxylin and eosin (H and E) according to the methods described in our previous studies [
36]. Observation of the sections was conducted under a light microscope (Leica DM5500B, Solms, Germany), and morphometric analysis was performed with Image J software (NIH, Bethesda, MD, USA).
2.4.5. Gut Microbiota Collection and Analyses
The extraction of bacterial DNA from a large yellow croaker gut was performed using the HiPure Soil DNA Kit (Magen, Beijing, China), then Nanodrop 2000 (Thermo Scientific, Waltham, MA, USA) was used to detect the DNA yield. Amplicons of the V3-V4 region of the 16SrRNA gene were extracted from a 2% agarose gel and purified using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, San Francisco, CA, USA). Quantification was then performed using the ABI StepOnePlus Real-Time PCR System (Life Technologies, New York, NY, USA). The purified amplicons were sent to Illumina Miseq PE 250 for sequence analysis, performed by Gene Denovo Biotechnology Co., Ltd., Guangzhou, China.
All chimeric tags were removed with QIIME software to obtain valid tags for further analysis. The valid tags were clustered into operational taxonomic units (OTUs) of ≥97% similarity using UPARSE software. The highest abundant tags sequence was selected as the representative sequence within each cluster. The biological classification of representative sequences was performed using the RDP classifier based on the SILVA database with a confidence threshold of 80%. An NMDS plot with an unweighted uniFrace distance matrix was used to represent the variability of the intestinal bacterial community structure. LEFSE analysis was performed to compare gut microbial community composition among all groups.
Table 2.
Formulation and proximate composition of experimental diets (% dry matter).
Table 2.
Formulation and proximate composition of experimental diets (% dry matter).
| Ingredients (%) | Experimental Diets |
|---|
| TM0 | TM15 | TM30 | TM45 |
|---|
| Fish meal | 40.00 | 34.00 | 28.00 | 22.00 |
| Chicken meal a | 13.00 | 13.00 | 13.00 | 13.00 |
| Soybean meal | 10.00 | 10.00 | 10.00 | 10.00 |
| TM b | 0.00 | 6.70 | 13.30 | 20.00 |
| High gluten flour | 23.00 | 21.39 | 19.87 | 18.25 |
| Soybean oil | 1.50 | 1.50 | 1.50 | 1.50 |
| Fish oil | 2.50 | 2.90 | 3.30 | 3.70 |
| Lecithin | 1.50 | 1.50 | 1.50 | 1.50 |
| Stickwater c | 6.00 | 6.00 | 6.00 | 6.00 |
| Vitamin C d | 0.10 | 0.10 | 0.10 | 0.10 |
| Vitamin premix e | 0.40 | 0.40 | 0.40 | 0.40 |
| Mineral premix f | 0.50 | 0.50 | 0.50 | 0.50 |
| Ca(H2PO4)2 | 1.00 | 1.50 | 2.00 | 2.50 |
| Choline chloride | 0.50 | 0.50 | 0.50 | 0.50 |
| Methionine | 0.00 | 0.01 | 0.03 | 0.05 |
| Proximate analysis |
| Dry matter | 98.02 | 98.49 | 98.59 | 98.85 |
| Crude protein | 47.98 | 48.30 | 48.00 | 48.10 |
| Crude lipid | 8.25 | 8.63 | 8.76 | 8.79 |
| Ash | 11.12 | 10.70 | 10.85 | 10.50 |
| Methionine | 2.60 | 2.64 | 2.67 | 2.71 |
| Lysine | 0.93 | 0.93 | 0.93 | 0.94 |
| Total Phosphorus | 1.38 | 1.38 | 1.38 | 1.37 |
2.5. Calculations and Statistical Analysis
Survival rate (SR, %) = final body number/initial body number × 100
Weight gain rate (WG, %) = (final wet body weight − initial wet body weight)/initial wet body weight × 100
Feed efficiency (FE) = (final wet body weight − initial wet body weight)/dry feed intake
Feed intake (FI, %/day) = total dry feed intake × 2/[(final wet body weight + initial wet body weight) × number of feeding days] × 100
Protein efficiency ratio (PER) = wet body weight gain/dry protein intake × 100
Condition factor (CF, g/cm3) = final wet body weight/(final body length)3
Hepatosomatic index (HSI, %) = liver wet weight/final wet body weight × 100
Viscerosomatic index (VSI, %) = visceral wet weight/final wet body weight × 100
Protein deposition rate (PDR, %) = (final wet body weight × crude protein in final body − initial wet body weight × crude protein in initial body)/crude protein in diet × 100
Lipid deposition rate (LDR, %) = (final wet body weight × crude lipid in final body − initial wet body weight × crude lipid in initial body)/crude lipid in diet × 100
All data were analyzed by one-way ANOVA using SPSS 22.0, and multiple comparisons were performed using Tukey’s test. p < 0.05 is set to indicate a significant level of difference. The data are presented as means ± S.E.M. (standard error of the mean).
4. Discussion
In this study, the replacement of 30% FM with TM resulted in no obvious difference in the growth performance of fish. The result indicated that TM could be used as a partial substitution for FM. However, TM significantly affected WG when the substitution level reached 45%. Similarly, a study on African catfish (
Clarias gariepinus) showed that the growth performance of the fish was significantly reduced when the TM substitution level exceeded 40% [
38], and a study on European perch (
Perca fluviatilis) found a decreased growth performance when TM substituted 50% FM [
39]. Furthermore, in the present study, FE, PER, LDR, and PDR were reduced in the TM30 and TM45 groups, indicating that high levels of substitution affected feed utilization. In fact, TM contains about 4.92% of chitin [
40]. Some studies have confirmed that a diet containing more than 1% chitin will reduce the growth performance of fish [
41], which may be one of the reasons for the reduced growth performance of large yellow croakers in the high TM group in this experiment. The above results indicate that replacing 15% FM with TM in the ration is a reasonable substitution level with no adverse effects on the growth performance of juvenile large yellow croakers.
The addition of TM to diets did not affect the crude protein and crude lipid content of the whole body but increased the moisture and ash content, which is consistent with the results of the largemouth bass (
Micropterus salmoides) study [
42]. In addition, the crude lipid content of the liver was reduced in the TM15 group. However, studies in largemouth bass (
Micropterus salmoides) [
41] showed a TM substitution of up to 50% FM in the diet did not affect liver crude lipid content. This needs further study in the future.
Fish humoral immunity is an important component of fish immunity, while serum immune parameters can reflect the immune function of fish [
43]. AKP had been proved to have immunomodulatory effects by inhibiting neutrophil production and inflammatory responses [
44]. The decreased AKP activity in serum leads to the decreased immunity of fish [
45]. LZM is produced by neutrophils and macrophages and secreted into the blood and mucus to exert bacteriolytic effects with antiviral, antibacterial, and anti-inflammatory properties [
46]. Furthermore, complement is an important component of fish resistance to microbial infection and is involved in target cell lysis and the regulatory responses of the organism after initiation, while complement 3 and complement 4 are the main components of the complement system [
47]. Increasing the serum levels of C3 and C4 improves immune regulation, neutralizes toxins, and blocks inflammatory responses [
48]. The results of the present study showed that replacing 45% of FM with TM in diets reduced AKP activity. Replacing 15% of FM with TM increased LZM activity, and replacing 30% of FM increased C3 contents while replacing 30% or more FM decreased C4 contents. These results indicate that the high-level addition of TM leads to a decrease in the immunity function of the large yellow croaker.
It is widely accepted that intestinal histology is a required indicator to assess tissue alterations caused by different diets [
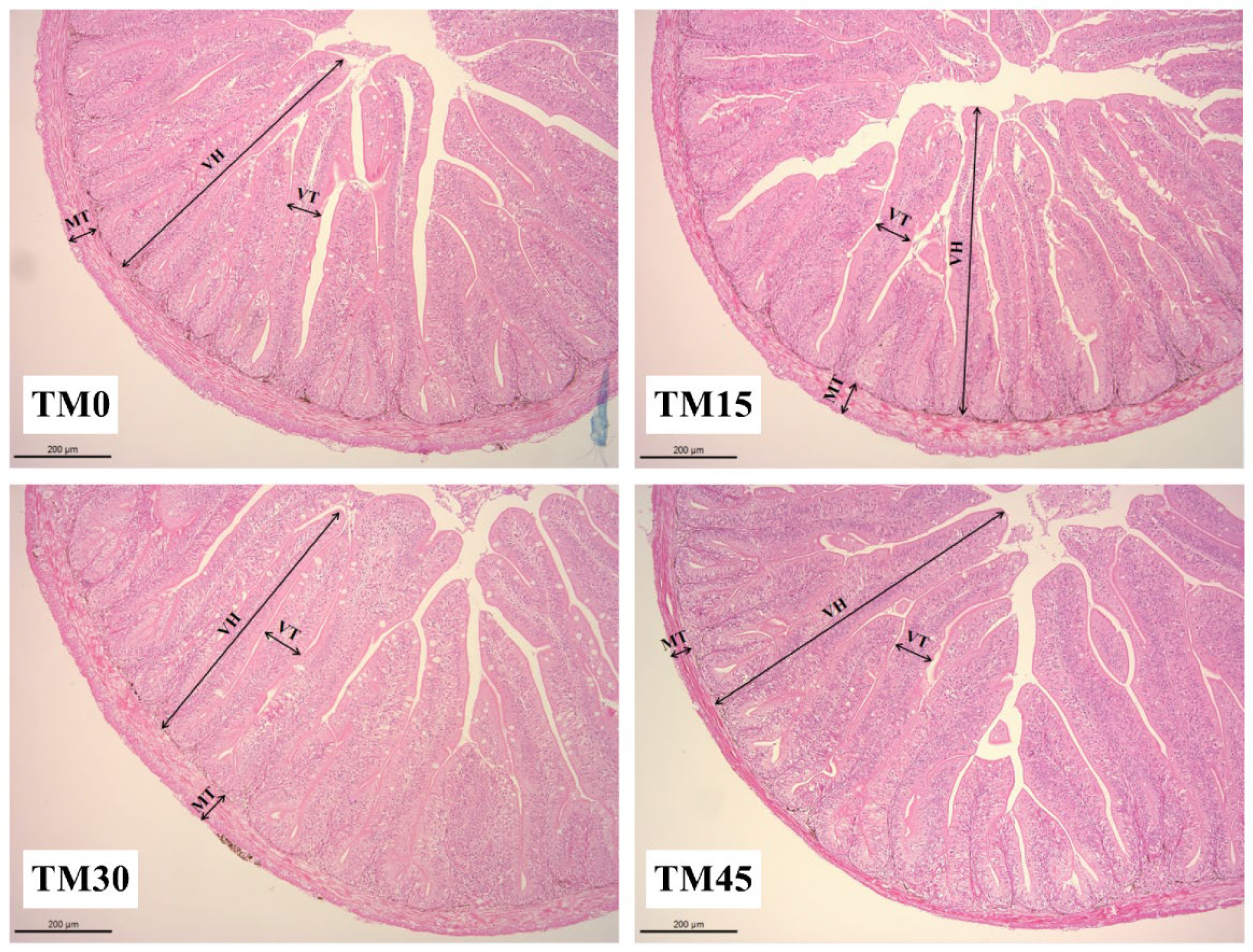
49]. Histological analysis through HE straining is a direct way to assess the health of the intestine. It is generally believed that the longer the height and thickness of the intestinal villi, the greater the digestion and absorption capacity of the intestine for nutrients by increasing the contact area with nutrients [
50], and muscle stenosis may cause intestinal inflammation to harm intestinal health [
51]. In the present study, TM diets increase the thickness of the intestinal villi in large yellow croakers; 15% and 45% substitution levels increase the height of the intestinal villi, and 15% and 30% substitution levels increase the thickness of the muscularis. These results suggest that TM may have a beneficial effect on intestinal health. Similar results were also observed in a previous study that used largemouth bass (
Micropterus salmoides) [
42]. Serum concentrations of D-lactate and DAO often increase when there is damage to intestinal epithelial structures [
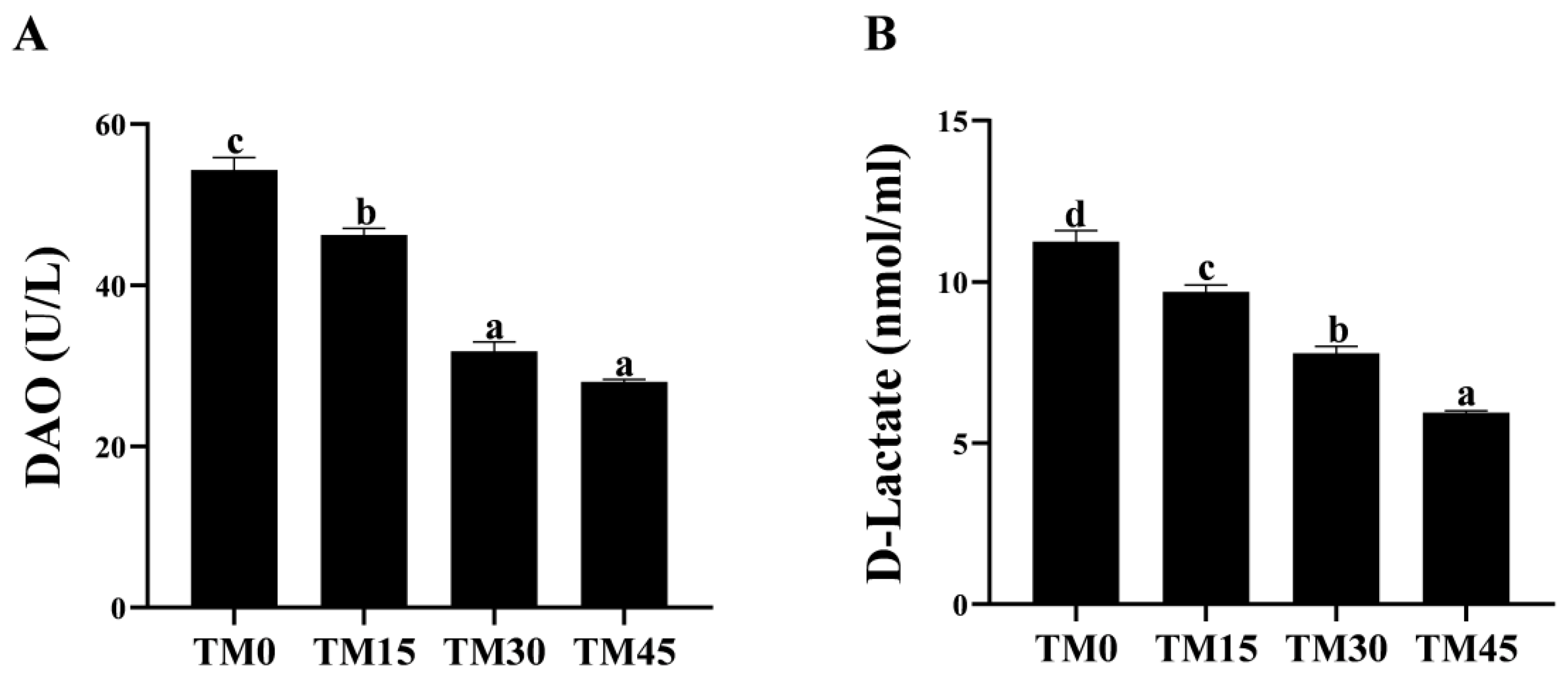
52]. In the present study, we evaluated intestinal epithelial permeability by measuring serum D-lactate and DAO levels/activity. As a result, TM reduced the serum concentrations of D-lactate and DAO, suggesting that TM has a repairing effect on the intestinal epithelial mucosa of large yellow croakers.
To a certain extent, digestive enzyme activity reflects the degree of the digestion and absorption of feed materials and nutrients by the organism [
53]. In the present study, TM did not affect the intestinal trypsin activity of large yellow croakers, but 15% and 45% substitution levels increase the intestinal lipase activity. Research has been conducted to show that fatty acids in insect proteins are incompatible with the digestion of
Pangasianodon hypophthalmus [
54] and that fatty acids act as signaling molecules to regulate lipid metabolic pathways such as lipase [
55], which may account for the elevated intestinal lipase activity in the TM high-substitution-level group. The above results indicate that TM has a promotive effect on the intestinal lipase enzymes of large yellow croakers.
Gut flora can influence the physiological, nutritional, immune, and metabolic functions of the host and plays a crucial role in the health status of marine fish [
56,
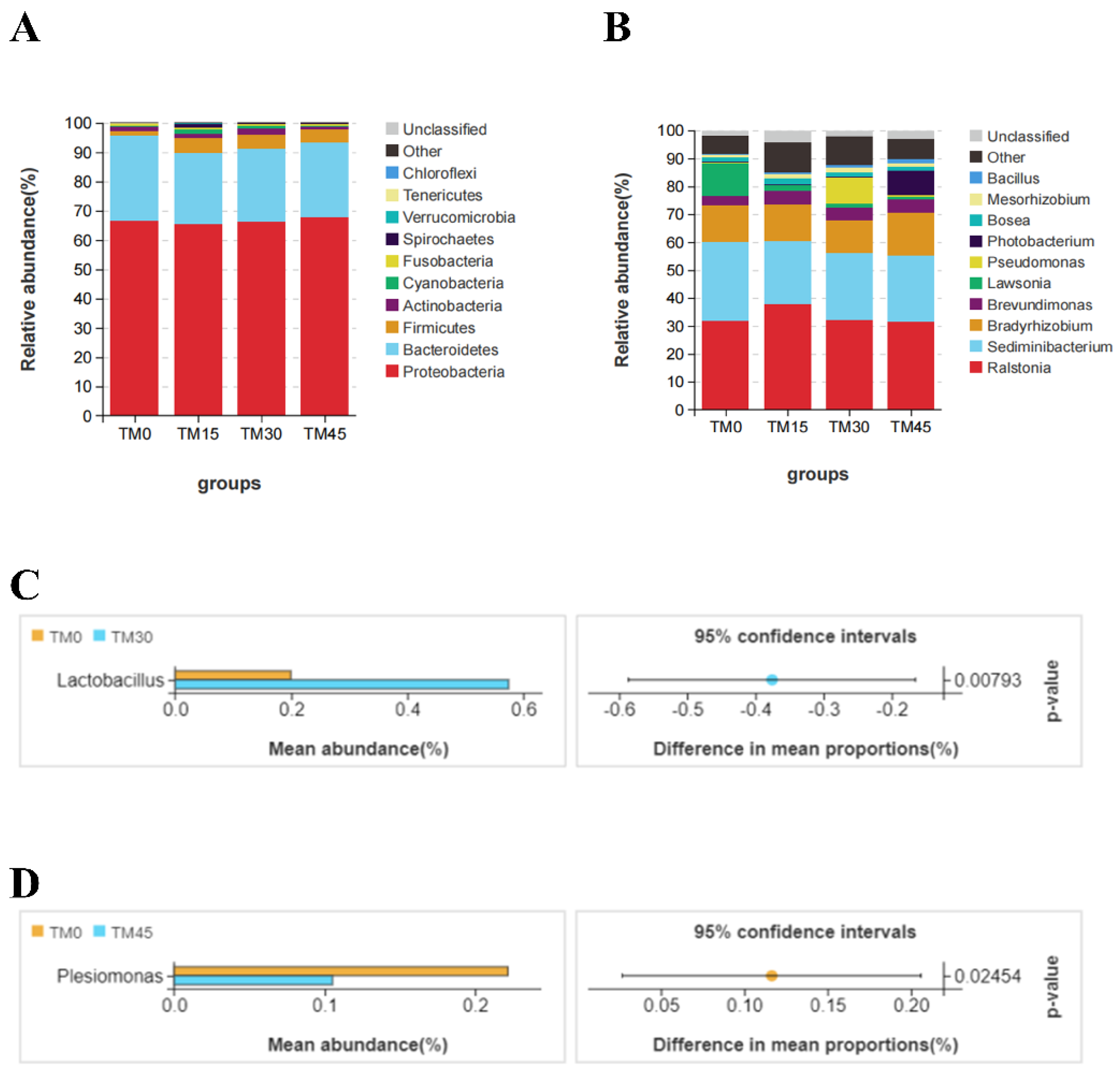
57]. In the present study, the three major phyla in the gut of large yellow croakers were
Proteobacteria,
Bacteroidetes, and
Firmicutes.
Ralstonia,
Sediminibacterium, and
Bradyrhizobium are the three major genera of bacteria. The relative abundance of
Pseudomonas was higher in TM30 and TM45 compared to TM0, and the former was shown to be a fish pathogen [
58]. This suggests that high levels of TM can adversely affect the composition of the intestinal flora of large yellow croakers. A previous study found that probiotic bacteria in
Lactobacillus are beneficial in aquaculture to protect fish from pathogens [
59,
60,
61]. In this study, TM30 increased the abundance of the genus
Lactobacillus. In contrast, bacteria in
Plesiomonas are a pathogen in fish, and human accidental ingestion of fish containing this bacterium is highly susceptible to gastrointestinal-related diseases [
62,
63]. Currently, the abundance of
Plesiomonas was significantly reduced with a 45% level of substitution. In addition, TM significantly improved the abundance of bacteria in the Bacilli in the intestinal tract of large yellow croakers, the former of which had been reported to be a probiotic bacteria with beneficial effects on the growth and intestinal health of fish [
64,
65,
66]. Moreover, the 30% and 45% substitution levels significantly reduced the abundance of Peptostreptococcaceae at the family level in the intestine, which was shown to be a potentially pathogenic bacterium that affects metabolic disorders [
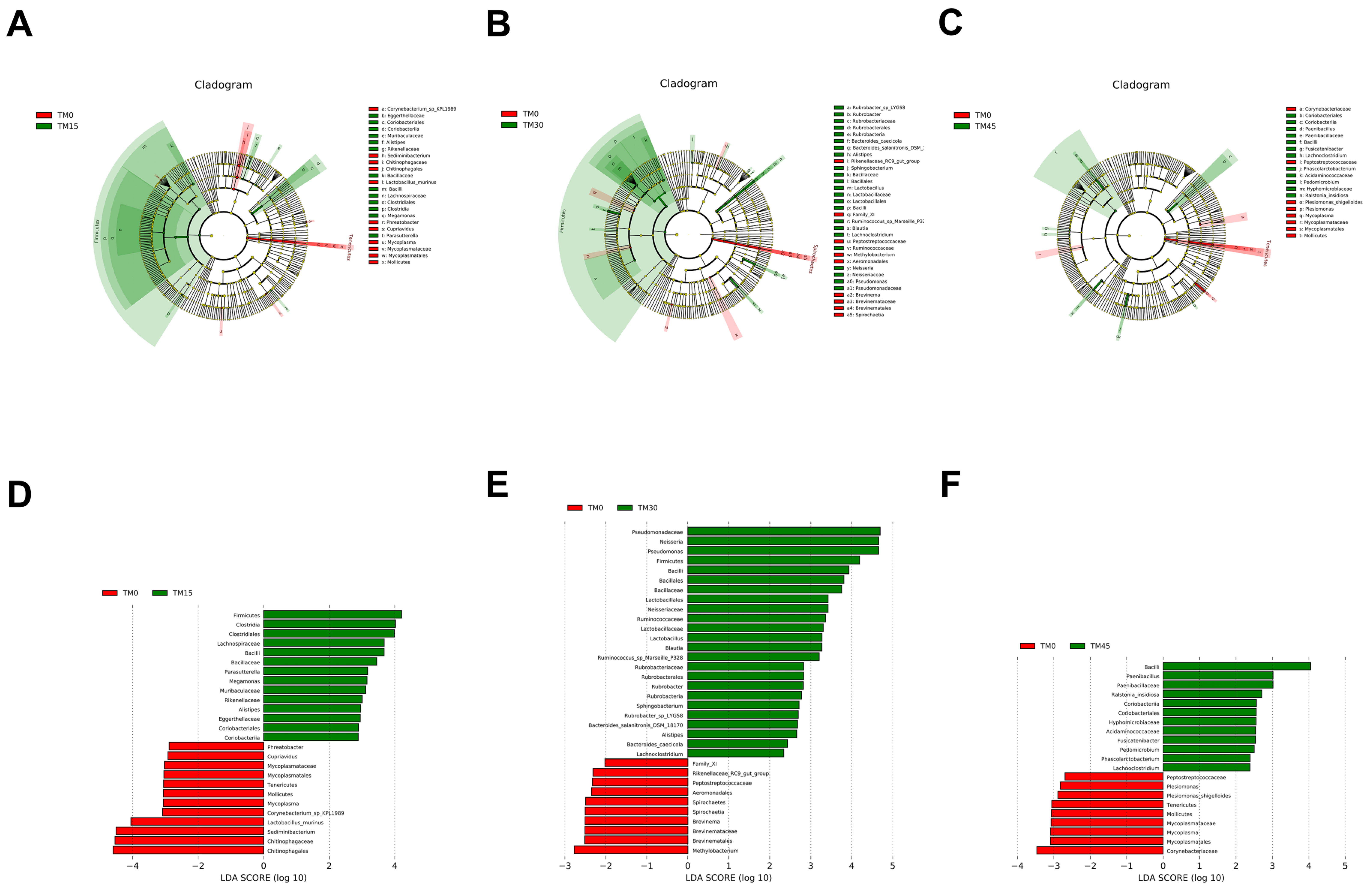
67]. These data suggest that TM selectively improved the proliferation of the intestinal flora and changed the flora composition of large yellow croakers.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





