

Silage Mixtures of Alfalfa with Sweet Sorghum Alter Blood and Rumen Physiological Status and Rumen Microbiota of Karakul Lambs
 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Silage Mixtures and Experimental Diets
2.2. Animal Feeding and Sampling
2.3. Analysis of Blood Physiological Status
2.4. Analysis of Rumen Physiological Status
2.5. DNA Extraction, 16S rDNA Gene Amplification, Sequence Processing and Analysis
2.6. Statistical Analysis
3. Results
3.1. Physiological Status in Blood
3.2. DM Intake and Physiological Status in Rumen
3.3. Alpha Diversity and Analysis of OTUs
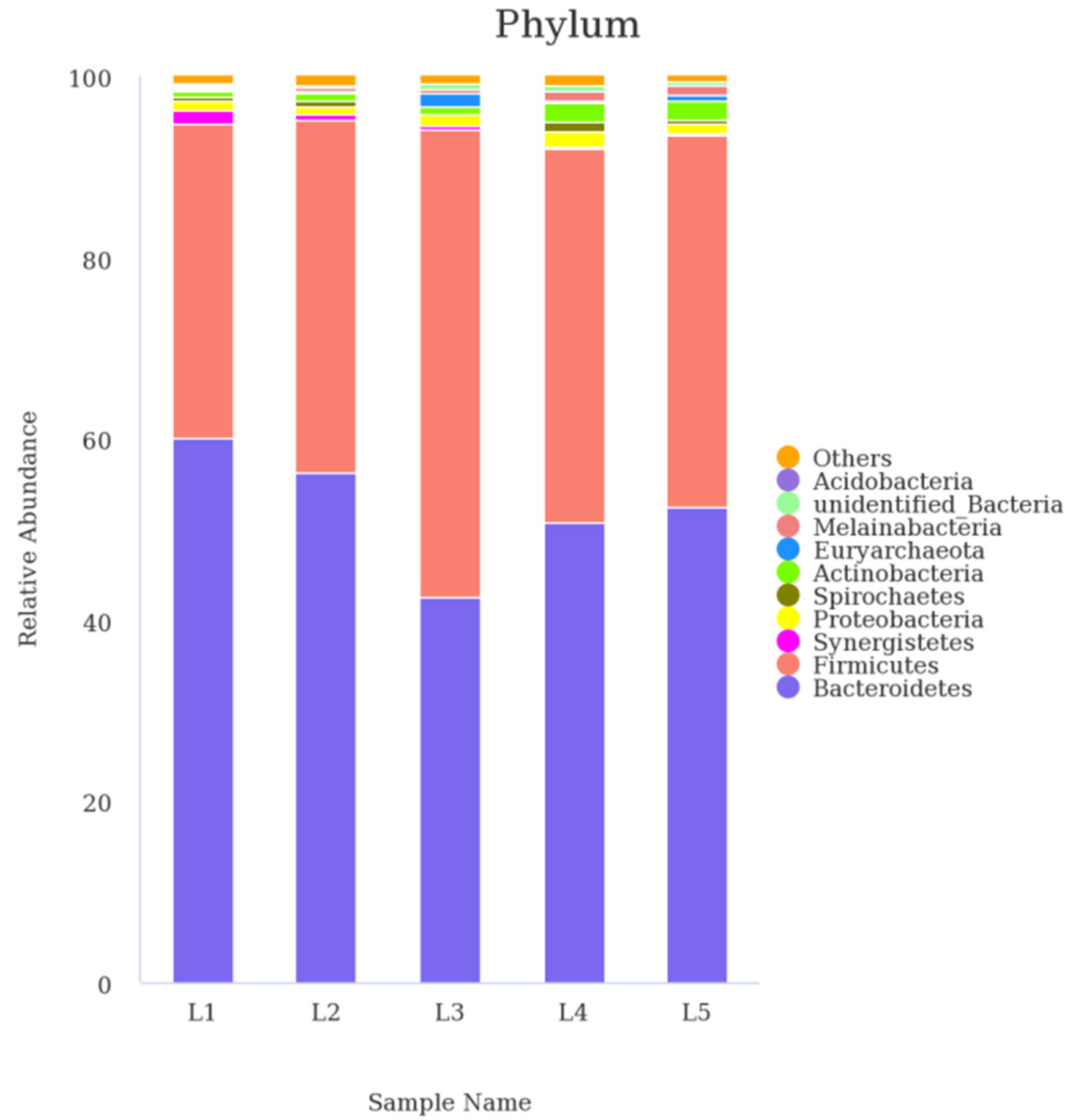
3.4. Ruminal Bacterial Communities at Phylum
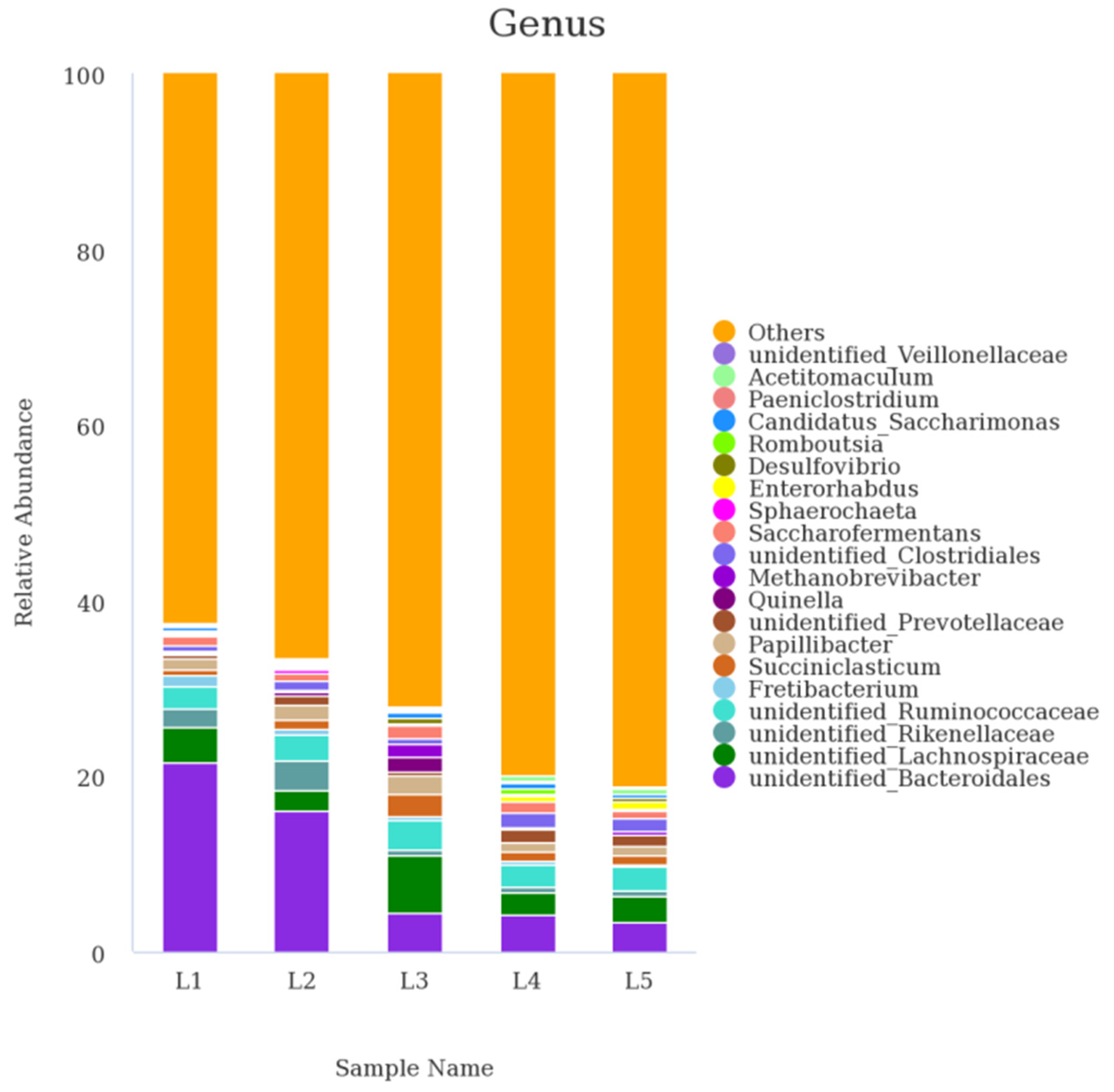
3.5. Ruminal Bacterial Communities at Genus and Species
4. Discussion
4.1. Physiological Status in Blood
4.2. DM Intake and Physiological Status in Rumen
4.3. Ruminal Bacterial Communities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Easdale, M.H.; Aguiar, M.R. Regional forage production assessment in arid and semi-arid rangelands-A step towards social-ecological analysis. J. Arid Environ. 2012, 83, 35–44. [Google Scholar] [CrossRef]
- Cremilleux, M.; Coppa, M.; Bouchon, M.; Delaby, L.; Beaure, G.; Constant, I.; Natalello, A.; Martin, B.; Michaud, A. Effects of forage quantity and access-time restriction on feeding behaviour, feed efficiency, nutritional status, and dairy performance of dairy cows fed indoors. Animal 2022, 9, 100608. [Google Scholar] [CrossRef] [PubMed]
- Pupo, M.R.; Wallau, M.O.; Ferraretto, L.F. Effects of season, variety type, and trait on dry matter yield, nutrient composition, and predicted intake and milk yield of whole-plant sorghum forage. J. Dairy Sci. 2022, 105, 5776–5785. [Google Scholar] [CrossRef]
- Kurle, J.E.; Sheaffer, C.C.; Crookston, R.K.; Peterson, R.H.; Chester-Jones, H.; Lueschen, W.E. Popcorn, sweet corn, and sorghum as alternative silage crops. J. Prod. Agric. 1991, 4, 432–436. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, X.; Tan, T. Ethanol production by solid state fermentation of sweet sorghum using thermotolerant yeast strain. Fuel Process. Technol. 2008, 89, 1056–1059. [Google Scholar] [CrossRef]
- Zhang, S.J.; Chaudhry, A.S.; Ramdani, D.; Osman, A.; Guo, X.F.; Edwards, G.R.; Cheng, L. Chemical composition and in vitro fermentation characteristics of high sugar forage sorghum as an alternative to forage maize for silage making in Tarim Basin, China. J. Integr. Agric. 2016, 15, 175–182. [Google Scholar] [CrossRef]
- Caraschi, J.C.; Viana, R.S.; Moreira, B.R.A.; Prates, G.A. Chemical and energetic characteristics of African sweet sorghum as a source of bioenergy. Eng. Agrícola 2019, 39, 426–433. [Google Scholar] [CrossRef]
- Zhang, S.J.; Chaudhry, A.S.; Osman, A.; Shi, C.Q.; Edwards, G.R.; Dewhurst, R.J.; Cheng, L. Associative effects of ensiling mixtures of sweet sorghum and alfalfa on nutritive value, fermentation and methane characteristics. Anim. Feed Sci. Tech. 2015, 206, 29–38. [Google Scholar] [CrossRef]
- Wang, J.; Yang, B.Y.; Zhang, S.J.; Amar, A.; Chaudhry, A.S.; Cheng, L.; Abbasi, I.H.R.; Al-Mamun, M.; Guo, X.F.; Shan, A.S. Using mixed silages of sweet sorghum and alfalfa in total mixed rations to improve growth performance, nutrient digestibility, carcass traits and meat quality of sheep. Animal 2021, 15, 100246. [Google Scholar] [CrossRef]
- Fijałkowska, M.; Pysera, B.; Lipiński, K.; Strusińska, D. Changes of nitrogen compounds during ensiling of high protein herbages—A review. Ann. Anim. Sci. 2015, 15, 289–305. [Google Scholar] [CrossRef] [Green Version]
- Besharati, M.; Niazifar, M.; Nemati, Z.; Palangi, V. Comparison study of flaxseed, cinnamon and lemon seed essential oils additives on quality and fermentation characteristics of lucerne silage. Acta Agric. Slov. 2020, 115, 455–462. [Google Scholar] [CrossRef]
- Cui, K.; Qi, M.; Wang, S.; Diao, Q.; Zhang, N. Dietary energy and protein levels influenced the growth performance, ruminal morphology and fermentation and microbial diversity of lambs. Sci. Rep. 2019, 9, 16612. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Flores, M.D.; Molina-Botero, I.C.; Arango, J.; Romano-Muñoz, J.L.; Solorio-Sánchez, F.J.; Aguilar-Pérez, C.F.; Ku-Vera, J.C. Effect of dried leaves of leucaena leucocephala on rumen fermentation, rumen microbial population, and enteric methane production in crossbred heifers. Animals 2020, 10, 300. [Google Scholar] [CrossRef] [PubMed]
- Lefter, N.A.; Vasilachi, A.; Voicu, D.; Hăbeanu, M.; Gheorghe, A.; Grosu, A.I. Effect of sorghum grain inclusion in Montbeliarde dairy cows diet on health status. Slovak J. Anim. Sci. 2019, 52, 63–68. [Google Scholar]
- Wang, B.; Luo, H. Effects of mulberry leaf silage on antioxidant and immunomodulatory activity and rumen bacterial community of lambs. BMC Microbiol. 2021, 21, 250. [Google Scholar] [CrossRef]
- Weatherburn, M.W. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Julák, J.; Stránská, E.; Procházková-Francisci, E.; Rosová, V. Blood cultures evaluation by gas chromatography of volatile fatty acids. Med. Sci. Monit. 2000, 6, 605–610. [Google Scholar]
- Sun, Z.; Yu, Z.; Wang, B. Perilla frutescens leaf alters the rumen microbial community of lactating dairy cows. Microorganisms 2019, 7, 562. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Rogens, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Memon, M.A.; Wang, Y.; Xu, T.; Ma, N.; Zhang, H.; Roy, A.C.; ul Aabdin, Z.; Shen, X. Lipopolysaccharide induces oxidative stress by triggering MAPK and Nrf2 signalling pathways in mammary glands of dairy cows fed a high-concentrate diet. Microb. Pathog. 2019, 128, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Kurata, M.; Suzuki, M.; Agar, N.S. Antioxidant systems and erythrocyte life-span in mammals. Comp. Biochem. Physiol. B 1993, 106, 477–487. [Google Scholar] [CrossRef]
- Yonathan, A.; Koji, W.; Yuriko, I.; Yasuhiro, K.; Masami, M.; Mizuki, M.; Makoto, T. Compositions, taste characteristics, volatile profiles, and antioxidant activities of sweet sorghum (Sorghum bicolor L.) and sugarcane (Saccharum officinarum L.) syrups. J. Food Meas. Charact. 2018, 12, 884–891. [Google Scholar]
- Chen, S.; Li, X.; Liu, X.; Wang, N.; An, Q.; Ye, X.M.; Zhao, Z.T.; Zhao, M.; Han, Y.; Ouyang, K.H.; et al. Investigation of chemical composition, antioxidant activity, and the effects of alfalfa flavonoids on growth performance. Oxid. Med. Cell. Longev. 2020, 10, 8569237. [Google Scholar] [CrossRef]
- Parra, M.C.; Costa, D.F.; Meale, S.J.; Silva, L.F.P. Rumen bacteria and feed efficiency of beef cattle fed diets with different protein content. Anim. Prod. Sci. 2022, 62, 1029–1039. [Google Scholar] [CrossRef]
- Tao, Y.; Sun, Q.; Li, F.; Xu, C.; Cai, Y. Comparative analysis of ensiling characteristics and protein degradation of alfalfa silage prepared with corn or sweet sorghum in semiarid region of Inner Mongolia. Anim. Sci. J. 2020, 91, e13321. [Google Scholar] [CrossRef]
- Mohsen, M.K.; Ali, M.F.; Gaafar, H.M.; Al-Sakka, T.S.; Aboelenin, S.M.; Soliman, M.M.; Dawood, M.A.O. Impact of dry sugar beet pulp on milk production, digestibility traits, and blood constituents of dairy holstein cows. Animals 2021, 11, 3496. [Google Scholar] [CrossRef]
- Golder, H.M.; Celi, P.; Rabiee, A.R.; Heuer, C.; Bramley, E.; Miller, D.W.; Lean, I.J. Effects of grain, fructose, and histidine on ruminal pH and fermentation products during an induced subacute acidosis protocol. J. Dairy Sci. 2012, 95, 1971–1982. [Google Scholar] [CrossRef] [PubMed]
- Polyorach, S.; Wanapat, M.; Cherdthong, A. Influence of yeast fermented cassava chip protein (YEFECAP) and roughage to concentrate ratio on ruminal fermentation and microorganisms using in vitro gas production technique. Asian-Austral. J. Anim. Sci. 2014, 27, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Gunun, N.; Ouppamong, T.; Khejornsart, P.; Cherdthong, A.; Wanapat, M.; Polyorach, S.; Kaewpila, C.; Kang, S.; Gunun, P. Effects of rubber seed kernel fermented with yeast on feed utilization, rumen fermentation and microbial protein synthesis in dairy heifers. Fermentation 2022, 8, 288. [Google Scholar] [CrossRef]
- Vinh, N.T.; Wanapat, M.; Khejornsart, P.; Kongmun, P. Studies of diversity of rumen microorganisms and fermentation in swamp buffalo fed different diets. J. Anim. Vet. Adv. 2011, 10, 406–414. [Google Scholar] [CrossRef]
- Kang, S.; Wanapat, M.; Cherdthorng, A. Effect of banana flower powder supplementation as a rumen buffer on rumen fermentation efficiency and nutrient digestibility in dairy steers fed a high-concentrate diet. Anim. Feed Sci. Tech. 2014, 196, 32–41. [Google Scholar] [CrossRef]
- Benchaar, C.; Petit, H.V.; Berthiaume, R.; Ouellet, D.R.; Chiquette, J.; Chouinard, P.Y. Effects of essential oils on digestion, ruminal fermentation, rumen microbial populations, milk production, and milk composition in dairy cows fed alfalfa silage or corn silage. J. Dairy Sci. 2007, 90, 886–897. [Google Scholar] [CrossRef]
- Saro, C.; Ranilla, M.J.; Tejido, M.L.; Carroc, M.D. Influence of forage type in the diet of sheep on rumen microbiota and fermentation characteristics. Livest. Sci. 2014, 160, 52–59. [Google Scholar] [CrossRef]
- Niyigena, V.; Coffey, K.P.; Coblentz, W.K.; Philipp, D.; Althaber, C.; Diaz Gomez, J.; Rhein, R.T.; Pruden, M.C. Intake, digestibility rumen fermentation and nitrogen balance in lambs offered alfalfa and tall fescue-mixtures harvested and ensiled after a frost. Anim. Feed Sci. Tech. 2022, 286, 115286. [Google Scholar] [CrossRef]
- He, Y.; Niu, W.; Qiu, Q.; Xia, C.; Shao, T.; Wang, H.; Li, Q.; Yu, Z.; Gao, Z.; Rahman, M.A.U.; et al. Effect of calcium salt of long-chain fatty acids and alfalfa supplementation on performance of Holstein bulls. Oncotarget 2018, 9, 3029–3042. [Google Scholar] [CrossRef]
- Njokweni, S.G.; Weimer, P.J.; Warburg, L.; Botes, M.; van Zyl, W.H. Valorisation of the invasive species, Prosopis julifora, using the carboxylate platform to produce volatile fatty acids. Bioresour. Technol. 2019, 288, 121602. [Google Scholar] [CrossRef]
- Du, S.; You, S.; Sun, L.; Wang, X.; Jia, Y.; Zhou, Y. Effects of replacing alfalfa hay with native grass hay in pelleted total mixed ration on physicochemical parameters, fatty acid profile, and rumen microbiota in lamb. Front. Microbiol. 2022, 13, 861025. [Google Scholar] [CrossRef] [PubMed]
- Pu, X.; Guo, X.; Shahzad, K.; Wang, M.; Jiang, C.; Liu, J.; Zhang, X.; Zhang, S.; Cheng, L. Effects of dietary non-fibrous carbohydrate (NFC) to neutral detergent fiber (NDF) ratio change on rumen bacteria in sheep based on three generations of full-length amplifiers sequencing. Animals 2020, 10, 192. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Mu, C.; Xu, Y.; Shen, J.; Zhu, W. Changes in the solid-, liquid-, and epithelium-associated bacterial communities in the rumen of Hu lambs in response to dietary urea supplementation. Front. Microbiol. 2020, 11, 244. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Zeng, D.; Ni, X.; Zhu, H.; Jian, P.; Zhou, Y.; Xu, S.; Lin, Y.; Li, Y.; Yin, Z.; et al. Microbial community compositions in the gastrointestinal tract of Chinese Mongolian sheep using Illumina MiSeq sequencing revealed high microbial diversity. AMB Express 2017, 7, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Thoetkiattikul, H.; Mhuantong, W.; Laothanachareon, T.; Tangphatsornruang, S.; Pattarajinda, V.; Eurwilaichitr, L.; Champreda, V. Comparative analysis of microbial profiles in cow rumen fed with different dietary fiber by tagged 16s rRNA gene pyrosequencing. Curr. Microbiol. 2013, 67, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Belanche, A.; Doreau, M.; Edwards, J.E.; Moorby, J.M.; Pinloche, E.; Newbold, C.J. Shifts in the rumen microbiota due to the type of carbohydrate and level of protein ingested by dairy cattle are associated with changes in rumen fermentation. J. Nutr. 2012, 142, 1684–1692. [Google Scholar] [CrossRef] [PubMed]
- Da Silva-Marques, R.P.; Zervoudakis, J.T.; Nakazato, L.; Hatamoto-Zervoudakis, L.K.; da Silva Cabral, L.; do Nascimento Matos, N.B.; da Silva, M.I.L.; Feliciano, A.L. Ruminal microbial populations and fermentation characteristics in beef cattle grazing tropical forage in dry season and supplemented with different protein levels. Curr. Microbiol. 2019, 76, 270–278. [Google Scholar] [CrossRef]
- Lu, Z.; Xu, Z.; Shen, Z.; Tian, Y.; Shen, H. Dietary energy level promotes rumen microbial protein synthesis by improving the energy productivity of the ruminal microbiome. Front. Microbiol. 2019, 10, 847–861. [Google Scholar] [CrossRef]
- Huws, S.A.; Edwards, J.E.; Creevey, C.J.; Rees, S.P.; Lin, W.; Girdwood, S.E.; Pachebat, J.A.; Kingston-Smith, A.H. Temporal dynamics of the metabolically active rumen bacteria colonizing fresh perennial ryegrass. FEMS Microbiol. Ecol. 2016, 92, fiv137. [Google Scholar] [CrossRef]
- Sun, B.; Wang, X.; Bernstein, S.; Huffman, M.A.; Xia, D.P.; Gu, Z.; Chen, R.; Sheeran, L.K.; Wagner, R.S.; Li, J. Marked variation between winter and spring gut microbiota in free-ranging Tibetan Macaques (Macaca thibetana). Sci. Rep. 2016, 6, 26035. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, G.; Li, Y.; Zhang, Y. Effects of high forage/concentrate diet on volatile fatty acid production and the microorganisms involved in VFA production in cow rumen. Animals 2020, 10, 223. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Alves, S.P.; Cappucci, A.; Cook, S.R.; Duarte, A.; Caldeira, R.M.; McAllister, T.A.; Bessa, R.J.B. Efects of condensed and hydrolyzable tannins on rumen metabolism with emphasis on the biohydrogenation of unsaturated fatty acids. J. Agric. Food Chem. 2018, 66, 3367–3377. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Items | AF Percentage in the Silage Mixtures | ||||
|---|---|---|---|---|---|
| 0%AF | 20%AF | 40%AF | 60%AF | 80%AF | |
| Ingredients | |||||
| Silage mixtures | 40 | 40 | 40 | 40 | 40 |
| Rice straw | 10 | 10 | 10 | 10 | 10 |
| Cottonseed hull | 10 | 10 | 10 | 10 | 10 |
| Corn | 13 | 13 | 13 | 13 | 13 |
| Soybean meal | 12 | 12 | 12 | 12 | 12 |
| Wheat bran | 10 | 10 | 10 | 10 | 10 |
| Premix 1 | 5 | 5 | 5 | 5 | 5 |
| Nutrients | |||||
| DM, g/kg | 661.3 | 659.1 | 660.3 | 668.6 | 661.5 |
| CP, g/kg | 105.1 | 108.1 | 111.8 | 124.0 | 127.5 |
| EE, g/kg | 22.0 | 22.5 | 23.8 | 25.1 | 25.9 |
| NDF, g/kg | 555.2 | 541.9 | 528.2 | 507.0 | 503.8 |
| ADF, g/kg | 279.0 | 257.3 | 247.4 | 240.4 | 240.2 |
| Ca, g/kg | 12.2 | 11.4 | 13.1 | 11.1 | 12.5 |
| P, g/kg | 8.5 | 8.2 | 8.4 | 8.6 | 8.1 |
| ME, MJ/kg | 9.1 | 9.2 | 9.5 | 9.7 | 9.9 |
| Items | 0%AF | 20%AF | 40%AF | 60%AF | 80%AF | SEM 1 | Linear | Quadratic |
|---|---|---|---|---|---|---|---|---|
| Creatinine, umol/L | 86.92 | 93.76 | 95.94 | 96.33 | 90.67 | 1.18 | 0.53 | 0.14 |
| BUN, mmol/L | 4.56 | 4.79 | 5.10 | 5.25 | 5.96 | 0.59 | 0.24 | 0.62 |
| T-AOC, U/mL | 4.86 b | 5.59 b | 6.21 ab | 7.32 a | 7.60 a | 0.74 | 0.02 | 0.84 |
| SOD, U/mL | 97.81 b | 99.56 b | 110.33 a | 108.46 a | 101.23 b | 1.48 | <0.01 | <0.01 |
| T-CHO, mmol/L | 2.61 | 2.92 | 2.98 | 3.08 | 3.00 | 0.30 | 0.59 | 0.36 |
| Items | 0%AF | 20%AF | 40%AF | 60%AF | 80%AF | SEM 1 | Linear | Quadratic |
|---|---|---|---|---|---|---|---|---|
| DM intake, kg/d | 0.78 b | 0.85 b | 0.86 b | 0.92 a | 0.89 a | 0.02 | <0.01 | <0.01 |
| pH | 6.79 | 6.74 | 6.69 | 6.54 | 6.46 | 0.11 | 0.09 | 0.60 |
| NH3-N, mg/dL | 13.16 b | 13.76 b | 14.87 a | 15.53 a | 14.66 a | 0.43 | 0.02 | 0.06 |
| tVFA, mmol/L | 104.82 c | 112.82 bc | 123.90 ab | 128.77 a | 117.73 ab | 5.04 | <0.01 | <0.01 |
| Acetate, mmol/L | 59.34 c | 63.68 bc | 71.17 ab | 74.28 a | 64.43 bc | 3.74 | 0.03 | <0.01 |
| Propionate, mmol/L | 28.58 | 29.63 | 31.94 | 32.13 | 32.06 | 1.81 | 0.23 | 0.39 |
| Butyrate, mmol/L | 12.71 | 15.12 | 15.90 | 17.03 | 16.00 | 1.35 | 0.08 | 0.10 |
| Valerate, mmol/L | 1.79 | 1.90 | 2.12 | 2.28 | 2.36 | 0.37 | 0.10 | 0.91 |
| Isobutyrate, mmol/L | 0.92 | 0.97 | 1.10 | 1.22 | 1.16 | 0.10 | 0.07 | 0.43 |
| Isovalerate, mmol/L | 1.46 | 1.51 | 1.67 | 1.84 | 1.73 | 0.13 | 0.07 | 0.36 |
| Acetate:Propionate | 2.08 | 2.15 | 2.23 | 2.32 | 2.01 | 0.13 | 0.88 | 0.06 |
| Indices | 0%AF | 20%AF | 40%AF | 60%AF | 80%AF | SEM 1 | Linear | Quadratic |
|---|---|---|---|---|---|---|---|---|
| Observed species | 1649 | 1624 | 1420 | 1686 | 1617 | 188.75 | 0.67 | 0.47 |
| Shannon | 6.78 | 7.75 | 7.68 | 7.81 | 8.19 | 0.46 | 0.10 | 0.44 |
| Simpson | 0.90 | 0.98 | 0.99 | 0.98 | 0.99 | 0.04 | 0.21 | 0.17 |
| Chao1 | 1869.43 | 1891.07 | 1591.29 | 1990.49 | 1748.07 | 206.18 | 0.41 | 0.76 |
| ace | 1940.82 | 1946.88 | 1651.33 | 1985.36 | 1795.63 | 218.76 | 0.55 | 0.69 |
| Phylum | 0%AF | 20%AF | 40%AF | 60%AF | 80%AF | SEM 1 | Linear | Quadratic |
|---|---|---|---|---|---|---|---|---|
| Bacteroidetes | 59.95 | 56.35 | 42.44 | 50.73 | 52.41 | 5.70 | 0.09 | 0.06 |
| Firmicutes | 34.82 b | 38.86 b | 51.66 a | 41.21 ab | 40.93 ab | 4.63 | 0.04 | 0.03 |
| Proteobacteria | 1.20 | 0.85 | 1.19 | 1.65 | 1.20 | 0.72 | 0.86 | 0.97 |
| Synergistetes | 1.37 | 0.49 | 0.46 | 0.27 | 0.24 | 0.68 | 0.49 | 0.42 |
| Actinobacteria | 0.73 b | 0.81 b | 0.75 b | 2.12 a | 2.20 a | 0.45 | 0.01 | 0.26 |
| Spirochaetes | 0.30 | 0.61 | 0.05 | 1.08 | 0.32 | 0.66 | 0.60 | 0.76 |
| unidentified_Bacteria | 0.29 b | 0.31 b | 0.77 a | 0.54 ab | 0.36 b | 0.11 | <0.01 | <0.01 |
| Melainabacteria | 0.19 b | 0.33 b | 0.35 b | 1.12 a | 1.13 a | 0.32 | 0.03 | 0.56 |
| Tenericutes | 0.19 | 0.27 | 0.20 | 0.16 | 0.18 | 0.04 | 0.19 | 0.43 |
| Euryarchaeota | 0.12 b | 0.39 b | 1.54 a | 0.16 b | 0.47 b | 0.42 | 0.04 | 0.05 |
| Others | 0.22 | 0.34 | 0.31 | 0.25 | 0.15 | 0.08 | 0.31 | 0.07 |
| Total | 99.37 | 99.62 | 99.73 | 99.28 | 99.60 | 0.28 | 0.87 | 0.58 |
| Genus | 0%AF | 20%AF | 40%AF | 60%AF | 80%AF | SEM 1 | Linear | Quadratic |
|---|---|---|---|---|---|---|---|---|
| unidentified_Bacteroidales | 21.51 a | 16.16 b | 4.39 c | 4.32 c | 3.30 c | 9.07 | 0.02 | 0.42 |
| unidentified_Lachnospiraceae | 4.13 | 2.26 | 6.49 | 2.50 | 2.95 | 1.34 | 0.50 | 0.34 |
| unidentified_Ruminococcaceae | 2.56 | 2.97 | 3.33 | 2.61 | 2.87 | 0.44 | 0.80 | 0.26 |
| unidentified_Rikenellaceae | 2.20 | 3.38 | 0.77 | 0.59 | 0.68 | 1.44 | 0.30 | 0.97 |
| Fretibacterium | 1.37 | 0.49 | 0.46 | 0.27 | 0.22 | 0.68 | 0.48 | 0.43 |
| Papillibacter | 1.19 | 1.73 | 2.26 | 1.05 | 1.21 | 0.41 | 0.07 | 0.05 |
| Saccharofermentans | 0.98 | 0.77 | 1.45 | 1.28 | 0.93 | 0.31 | 0.26 | 0.20 |
| unidentified_Clostridiales | 0.74 | 0.98 | 0.63 | 1.59 | 1.43 | 0.41 | 0.06 | 0.65 |
| Succiniclasticum | 0.61 c | 1.11 b | 2.43 a | 1.12 b | 0.91 c | 0.56 | 0.63 | 0.02 |
| unidentified_Prevotellaceae | 0.55 | 1.08 | 0.47 | 1.45 | 1.09 | 0.44 | 0.18 | 0.88 |
| Romboutsia | 0.34 | 0.13 | 0.07 | 0.59 | 0.15 | 0.26 | 0.32 | 0.88 |
| Candidatus_Saccharimonas | 0.25 | 0.29 | 0.72 | 0.52 | 0.34 | 0.13 | 0.18 | 0.21 |
| Bacteroides | 0.23 | 0.16 | 0.26 | 0.20 | 0.20 | 0.10 | 0.90 | 0.96 |
| Quinella | 0.22 | 0.35 | 1.59 | 0.13 | 0.11 | 0.36 | 0.59 | 0.11 |
| Acetitomaculum | 0.21 b | 0.18 b | 0.20 b | 0.53 a | 0.65 a | 0.14 | <0.01 | 0.13 |
| Enterorhabdus | 0.16 c | 0.28 bc | 0.17 c | 0.62 a | 0.80 a | 0.17 | <0.01 | 0.17 |
| Methanobrevibacter | 0.11 | 0.37 | 1.51 | 0.15 | 0.45 | 0.42 | 0.64 | 0.06 |
| unidentified_Christensenellaceae | 0.11 | 0.08 | 0.08 | 0.13 | 0.12 | 0.03 | 0.43 | 0.41 |
| Fibrobacter | 0.10 | 0.08 | 0.04 | 0.09 | 0.06 | 0.04 | 0.39 | 0.59 |
| Bacillus | 0.06 | 0.27 | 0.01 | 0.09 | 0.16 | 0.11 | 0.22 | 0.84 |
| Others | 58.39 | 62.80 | 68.85 | 74.66 | 76.68 | 8.58 | 0.24 | 0.83 |
| Total | 95.82 | 95.93 | 96.19 | 94.48 | 95.31 | 1.34 | 0.73 | 0.89 |
| Species | 0%AF | 20%AF | 40%AF | 60%AF | 80%AF | SEM 1 | Linear | Quadratic |
|---|---|---|---|---|---|---|---|---|
| Ruminococcus_flavefaciens | 0.33 | 0.31 | 0.47 | 0.22 | 0.27 | 0.10 | 0.40 | 0.30 |
| Ruminococcus_albus | 0.06 b | 0.09 ab | 0.09 ab | 0.10 ab | 0.19 a | 0.05 | 0.04 | 0.35 |
| Butyrivibrio_fibrisolvens | 0.45 | 0.39 | 0.49 | 0.44 | 0.46 | 0.36 | 0.93 | 0.12 |
| Clostridium_disporicum | 0.05 | 0.06 | 0.02 | 0.06 | 0.08 | 0.03 | 0.31 | 0.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Cheng, L.; Chaudhry, A.S.; Khanaki, H.; Abbasi, I.H.R.; Ma, Y.; Abbasi, F.; Guo, X.; Zhang, S. Silage Mixtures of Alfalfa with Sweet Sorghum Alter Blood and Rumen Physiological Status and Rumen Microbiota of Karakul Lambs. Animals 2022, 12, 2591. https://doi.org/10.3390/ani12192591
Wang J, Cheng L, Chaudhry AS, Khanaki H, Abbasi IHR, Ma Y, Abbasi F, Guo X, Zhang S. Silage Mixtures of Alfalfa with Sweet Sorghum Alter Blood and Rumen Physiological Status and Rumen Microbiota of Karakul Lambs. Animals. 2022; 12(19):2591. https://doi.org/10.3390/ani12192591
Chicago/Turabian StyleWang, Jiao, Long Cheng, Abdul Shakoor Chaudhry, Hassan Khanaki, Imtiaz H. R. Abbasi, Yi Ma, Farzana Abbasi, Xuefeng Guo, and Sujiang Zhang. 2022. "Silage Mixtures of Alfalfa with Sweet Sorghum Alter Blood and Rumen Physiological Status and Rumen Microbiota of Karakul Lambs" Animals 12, no. 19: 2591. https://doi.org/10.3390/ani12192591