
Relative Bioavailability of Trace Minerals in Production Animal Nutrition: A Review


Abstract
:Simple Summary
Abstract
1. Introduction
2. Trace Minerals in Animal Nutrition
2.1. Inorganic Trace Minerals
2.2. Organic Trace Minerals
2.3. Mineral Uptake Mechanisms
2.3.1. Adequate Levels
Copper
Iron
Manganese
Zinc
2.3.2. Suboptimal Levels
Copper
Iron
Manganese
Zinc
3. Bioavailability
3.1. Evaluation of Bioavailability
3.1.1. Reference/Standard Source
3.1.2. Model Selection
3.1.3. Choice of Response Criteria
3.1.4. Comparison of Mineral Sources
3.2. Relative Bioavailability Tables
3.3. Key Species Observations from RBV Tables
3.3.1. Ruminants—Beef and Dairy
Copper Relative Bioavailability—Beef and Dairy
Iron Relative Bioavailability—Beef and Dairy
Manganese Relative Bioavailability—Beef and Dairy
Zinc Relative Bioavailability—Beef and Dairy
3.3.2. Ruminants—Sheep
Copper Relative Bioavailability—Sheep
Iron Relative Bioavailability—Sheep
Manganese Relative Bioavailability—Sheep
Zinc Relative Bioavailability—Sheep
3.3.3. Poultry
Copper Relative Bioavailability—Poultry
Iron Relative Bioavailability—Poultry
Manganese Relative Bioavailability—Poultry
Zinc Relative Bioavailability—Poultry
3.3.4. Swine
Copper Relative Bioavailability—Swine
Iron Relative Bioavailability—Swine
Manganese Relative Bioavailability—Swine
Zinc Relative Bioavailability—Swine
3.4. Mineral Stability and Associated Relationship with Bioavailability
4. Replacement of ITM with OTM in Feedstuffs
4.1. Ruminants
4.2. Poultry
4.3. Swine
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Amino acid |
| AAFCO | Association of American Feed Control Officials |
| Abs | Absorption |
| ACTH | Adrenocorticotropic hormone |
| BRD | Bovine respiratory disease |
| CAT | Cationic amino acid transporter |
| DM | Dry matter |
| DMT | Divalent metal transporter |
| EDTA | Ethylenediaminetetraacetic acid |
| EFSA | European Food Safety Authority |
| FP | Ferroportin |
| GLM | General linear model |
| HMTBa | 2-hydroxy-4-(methylthio)butanoate |
| ITM | Inorganic trace mineral(s) |
| MCT1 | Monocarboxylate transporter |
| MnSOD | Manganese superoxide dismutase activity |
| mRNA | Messenger ribonucleic acid |
| MT | Metallothionein |
| OTM | Organic trace mineral(s) |
| Qf | Formation quotient |
| RBV | Relative bioavailability value |
| SBM | Soybean meal |
| SOD | Superoxide dismutase |
| SPC | Soy protein concentrate |
| TBCC | dicopper chloride trihydroxide (or tribasic copper chloride) |
| TBZC | Zinc chloride hydroxide monohydrate (or tetrabasic zinc chloride) |
References
- PRISMA. Preferred Reporting Items for Systematic Reviews and Meta-Analyses. Available online: https://www.prisma-statement.org/ (accessed on 14 May 2022).
- PubMed. 2022. Available online: https://pubmed.ncbi.nlm.nih.gov/ (accessed on 14 May 2022).
- Scopus. 2022. Available online: https://www.scopus.com/ (accessed on 14 May 2022).
- WebofScience. 2022. Available online: https://clarivate.com/webofsciencegroup/solutions/web-of-science/ (accessed on 14 May 2022).
- ScienceDirect. 2022. Available online: https://www.sciencedirect.com/ (accessed on 14 May 2022).
- Ammerman, C.B.; Baker, D.H.; Lewis, A.J. (Eds.) Preface. In Bioavailability of Nutrients for Animals; Academic Press: San Diego, CA, USA, 1995; pp. xiii–xiv. [Google Scholar] [CrossRef]
- AAFCO. Feed terms and ingredient definitions. In American Association of Feed Control Officials; Eyck, R.T., Ed.; 2020 Official Publication; American Association of Feed Control Officials: Champaign, IL, USA, 2020. [Google Scholar]
- Hill, G.M.; Spears, J.W. Trace and Ultratrace elements in swine nutrition. In Swine Nutrition, 2nd ed.; Lewis, A.J., Southern, L.L., Eds.; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar] [CrossRef]
- Turnlund, J.R. Human whole-body copper metabolism. Am. J. Clin. Nutr. 1998, 67, 960S–964S. [Google Scholar] [CrossRef] [Green Version]
- Gaetke, L.; Chow, C. Copper toxicity, oxidative stress and antioxidant nutrients. Toxicology 2003, 189, 147–163. [Google Scholar] [CrossRef]
- Crapo, J.D.; Oury, T.; Rabouille, C.; Slot, J.W.; Chang, L.Y. Copper, zinc superoxide dismutase is primarily a cytosolic protein in human cells. Proc. Natl. Acad. Sci. USA 1992, 89, 10405. [Google Scholar] [CrossRef] [Green Version]
- Manto, M. Abnormal Copper Homeostasis: Mechanisms and Roles in Neurodegeneration. Toxics 2014, 2, 327–345. [Google Scholar] [CrossRef]
- Hill, G.M. Minerals and Mineral Utilization in Swine. In Sustainable Swine Nutrition; Chiba, L.I., Ed.; Wiley-Blackwell: Chichester, UK, 2012; pp. 173–195. [Google Scholar]
- EFSA. Revision of the currently authorised maximum copper content in complete feed; EFSA Panel on additives products or substances used in animal feed. EFSA J. 2016, 14, 4563. [Google Scholar] [CrossRef]
- Suttle, N.F. Mineral Nutrition of Livestock, 4th ed.; CABI Publishing: Wallingford, UK, 2010; pp. 1–547. [Google Scholar]
- Leeson, S. Copper metabolism and dietary needs. World’s Poult. Sci. J. 2009, 65, 353–366. [Google Scholar] [CrossRef]
- Europe Union. Commission Implementing Regulation (EU); 2018/1039 of 23 July 2018; Publications Office of the European Union: Luxembourg, 2018; Volume 186. [Google Scholar]
- Rincker, M.J.; Hill, G.M.; Link, J.E.; Rowntree, J.E. Effects of dietary iron supplementation on growth performance, hematological status, and whole-body mineral concentrations of nursery pigs. J. Anim. Sci. 2004, 82, 3189–3197. [Google Scholar] [CrossRef]
- Underwood, E.J. (Ed.) 2-Iron. In Trace Elements in Human and Animal Nutrition, 4th ed.; Academic Press: Cambridge, MA, USA, 1977; pp. 13–55. [Google Scholar] [CrossRef]
- Europe Union. Commission Implementing Regulation (EU); 2017/2330 of 14 December 2017; Publications Office of the European Union: Luxembourg, 2017; Volume 351. [Google Scholar]
- Cheeke, P.R. Applied Animal Nutrition: Feeds and Feeding; Pearson: Upper Saddle River, NJ, USA, 2005. [Google Scholar]
- Coomer, J. The Importance of Microminerals: Manganese. Available online: https://agriking.com/the-importance-of-micro-minerals-manganese/ (accessed on 12 November 2021).
- Henry, P.R. 11-Manganese bioavailability. In Bioavailability of Nutrients for Animals; Ammerman, C.B., Baker, D.H., Lewis, A.J., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 239–256. [Google Scholar] [CrossRef]
- Europe Union. Commission Implementing Regulation (EU); 2017/1490 of 21 August 2017; Publications office of the European Union: Luxembourg, 2017; Volume 216. [Google Scholar]
- Baker, D.H.; Ammerman, C.B. 17—Zinc bioavailability. In Bioavailability of Nutrients for Animals; Ammerman, C.B., Baker, D.H., Lewis, A.J., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 367–398. [Google Scholar] [CrossRef]
- Hara, T.; Takeda, T.A.; Takagishi, T.; Fukue, K.; Kambe, T.; Fukada, T. Physiological roles of zinc transporters: Molecular and genetic importance in zinc homeostasis. J. Physiol. Sci. JPS 2017, 67, 283–301. [Google Scholar] [CrossRef]
- Rink, L.; Gabriel, P. Zinc and the immune system. Proc. Nutr. Soc. 2000, 59, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Reese, D. Pharmacological Levels of Zinc in Nursery Diets—A Review; University of Nebraska-Lincoln: Lincoln, NE, USA, 1995. [Google Scholar]
- Europe Union. Commission Implementing Regulation (EU); 2016/1095 of 6 July 2016; Publications Office of the European Union: Luxembourg, 2016; Volume 182. [Google Scholar]
- Underwood, E.J.; Suttle, N.F. The Mineral Nutrition of Livestock; CABI Publishing: Wallingford, UK, 1999. [Google Scholar]
- Powell, J.J.; Whitehead, M.W.; Ainley, C.C.; Kendall, M.D.; Nicholson, J.K.; Thompson, R.P.H. Dietary minerals in the gastrointestinal tract: Hydroxypolymerisation of aluminium is regulated by luminal mucins. J. Inorg. Biochem. 1999, 75, 167–180. [Google Scholar] [CrossRef]
- Pesti, G.M.; Bakalli, R.I. Studies on the feeding of cupric sulfate pentahydrate and cupric citrate to broiler chickens. Poult. Sci. 1996, 75, 1086–1091. [Google Scholar] [CrossRef]
- Thompson, L.J.; Hall, J.O.; Meerdink, G.L. Toxic Effects of Trace Element Excess. Vet. Clin. N. Am. Food Anim. Pract. 1991, 7, 277–306. [Google Scholar] [CrossRef]
- Flohr, J.R.; DeRouchey, J.M.; Woodworth, J.C. A survey of current feeding regimens for vitamins and trace minerals in the US swine industry. J. Swine Health Prod. 2016, 24, 290–303. [Google Scholar]
- Broom, L.J.; Monteiro, A.; Piñon, A. Recent Advances in Understanding the Influence of Zinc, Copper, and Manganese on the Gastrointestinal Environment of Pigs and Poultry. Animals 2021, 11, 1276. [Google Scholar] [CrossRef]
- López-Alonso, M. Trace minerals and livestock: Not too much not too little. Int. Sch. Res. Not. Vet. Sci. 2012, 2012, 704825. [Google Scholar] [CrossRef] [Green Version]
- López-Alonso, M.; Miranda, M. Implications of excessive livestock mineral supplementation on environmental pollution and human health. In Trace Elements: Environmental Sources, Geochemistry and Human Health; Nova Science: New York, NY, USA, 2012; pp. 40–53. [Google Scholar]
- European Commission. Commission regulation (EC) No 1334/2003 of 25 July 2003 amending the conditions for authorisation of a number of additives in feedingstuffs belonging to the group of trace elements. In Official Journal of the European Union; European Union: Geneva, Switzerland, 2003; Volume 187, p. 11. [Google Scholar]
- Laven, R.A.; Livesey, C.T.; Offer, N.W.; Fountain, D. Apparent subclinical hepatopathy due to excess copper intake in lactating Holstein cattle. Vet. Rec. 2004, 155, 120–121. [Google Scholar] [CrossRef]
- Bidewell, C.A.; David, G.P.; Livesey, C.T. Copper toxicity in cattle. Vet. Rec. 2000, 147, 399–400. [Google Scholar]
- Henningson, J.N. Too Much of a Good Thing…Over Supplementation of Minerals in Cattle; Kansas State Veterinary Diagnostic Laboratory: Manhattan, KS, USA, 2016; Available online: https://www.ksvdl.org/resources/news/diagnostic_insights/january2016/over-supplementation.html (accessed on 5 December 2021).
- Bradley, C.H. Copper poisoning in a dairy herd fed a mineral supplement. Can. Vet. J. 1993, 34, 287–292. [Google Scholar]
- Minervino, A.H.H.; Barrêto Júnior, R.A.; Ferreira, R.N.F.; Rodrigues, F.A.M.L.; Headley, S.A.; Mori, C.S.; Ortolani, E.L. Clinical observations of cattle and buffalos with experimentally induced chronic copper poisoning. Res. Vet. Sci. 2009, 87, 473–478. [Google Scholar] [CrossRef]
- López-Alonso, M.; Crespo, A.; Miranda, M.; Castillo, C.; Hernández, J.; Benedito, J.L. Assessment of Some Blood Parameters as Potential Markers of Hepatic Copper Accumulation in Cattle. J. Vet. Diagn. Investig. 2006, 18, 71–75. [Google Scholar] [CrossRef] [Green Version]
- Standish, J.F.; Ammerman, C.B. Effect of Excess Dietary Iron as Ferrous Sulfate and Ferric Citrate on Tissue Mineral Composition of Sheep. J. Anim. Sci. 1971, 33, 481–484. [Google Scholar] [CrossRef]
- Campbell, A.G.; Coup, M.R.; Bishop, W.H.; Wright, D.E. Effect of elevated iron intake on the copper status of grazing cattle. N. Z. J. Agric. Res. 1974, 17, 393–399. [Google Scholar] [CrossRef]
- Standish, J.F.; Ammerman, C.B.; Palmer, A.Z.; Simpson, C.F. Influence of dietary iron and phosphorus on performance, tissue mineral composition and mineral absorption in steers. J. Anim. Sci. 1971, 33, 171–178. [Google Scholar] [CrossRef]
- Hall, E.D.; Symonds, H.W.; Mallinson, C.B. Maximum capacity of the bovine liver to remove manganese from portal plasma and the effect of the route of entry of manganese on its rate of removal. Res. Vet. Sci. 1982, 33, 89–94. [Google Scholar] [CrossRef]
- Symonds, H.W.; Hall, E.D. Acute manganese toxicity and the absorption and biliary excretion of manganese in cattle. Res. Vet. Sci. 1983, 35, 5–13. [Google Scholar] [CrossRef]
- Ho, S.Y.; Miller, W.J.; Gentry, R.P.; Neathery, M.W.; Blackmon, D.M. Effects of high but nontoxic dietary manganese and iron on their metabolism by calves. J. Dairy Sci. 1984, 67, 1489–1495. [Google Scholar] [CrossRef]
- Watson, L.T.; Ammerman, C.B.; Feaster, J.P.; Roessler, C.E. Influence of Manganese Intake on Metabolism of Manganese and Other Minerals in Sheep. J. Anim. Sci. 1973, 36, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.J.; Amos, H.E.; Gentry, R.P.; Blackmon, D.M.; Durrance, R.M.; Crowe, C.T.; Fielding, A.S.; Neathery, M.W. Long-term feeding of high zinc sulfate diets to lactating and gestating dairy cows. J. Dairy Sci. 1989, 72, 1499–1508. [Google Scholar] [CrossRef]
- Allen, J.G.; Masters, H.G.; Peet, R.L.; Mullins, K.R.; Lewis, R.D.; Skirrow, S.Z.; Fry, J. Zinc toxicity in ruminants. J. Comp. Pathol. 1983, 93, 363–377. [Google Scholar] [CrossRef]
- Coppenet, M.; Golven, J.; Simon, J.; Le Corre, L.; Le Roy, M. Chemical evolution of soils in intensive animal-rearing farms: The example of Finistere. Agronomie 1993, 13, 77–83. [Google Scholar] [CrossRef]
- Schlegel, P.; Durosoy, S.; Jongbloed, A.W. Trace Elements in Animal Production Systems; Wageningen Academic Publishers: Wageningen, The Netherlands, 2008. [Google Scholar]
- Bao, Y.M.; Choct, M.; Iji, P.A.; Bruerton, K. Effect of organically complexed copper, iron, manganese, and zinc on broiler performance, mineral excretion, and accumulation in tissues. J. Appl. Poult. Res. 2007, 16, 448–455. [Google Scholar] [CrossRef]
- Jarosz, Ł.; Marek, A.; Grądzki, Z.; Kwiecień, M.; Kalinowski, M. The effect of feed supplementation with zinc chelate and zinc sulphate on selected humoral and cell-mediated immune parameters and cytokine concentration in broiler chickens. Res. Vet. Sci. 2017, 112, 59–65. [Google Scholar] [CrossRef]
- European Commission. Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. In Official Journal of the European Union; European Union: Geneva, Switzerland, 2006; Volume 364, pp. 324–365. [Google Scholar]
- Byrne, L.; Hynes, M.J.; Connolly, C.D.; Murphy, R.A. Influence of the chelation process on the stability of organic trace mineral supplements used in animal nutrition. Animals 2021, 11, 1730. [Google Scholar] [CrossRef]
- Greene, L.W. The nutritional value of inorganic and organic mineral sources. Update of mineral nutrition of beef cattle. In Proceedings of the Plains Nutrition Council Symposium, San Antonio, TX, USA, 16–17 April 1995; pp. 23–32. [Google Scholar]
- Spears, J. Recent Developments in Trace Element Metabolism and Function. J. Nutr. 1989, 119, 1050. [Google Scholar] [CrossRef]
- Rompala, R.E.; Halley, J.T. Explaining the absorption of chelated trace minerals: The Trojan horse of nutrition. In Feed Management; WATT Publishing: Rockford, IL, USA, 1995; Volume 46, pp. 52–58. [Google Scholar]
- Magee, D.F.; Dalley, A.F., II. Digestion and the Structure and Function of the Gut; Karge Continuing Education Series; Karger: Basel, Switzerland, 1986; Volume 8. [Google Scholar]
- Radcliffe, J.S.; Aldridge, B.E.; Saddoris, K.L. Understanding Organic Mineral Uptake Mechanisms: Experiments with Bioplex® Cu. Engormix. 2007. Available online: https://en.engormix.com/pig-industry/articles/bioplex-cu-t33770.htm (accessed on 12 November 2021).
- McDowell, L.R. (Ed.) Chapter 1—General Introduction. In Minerals in Animal and Human Nutrition, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2003; pp. 1–32. [Google Scholar] [CrossRef]
- Goff, J.P. Invited review: Mineral absorption mechanisms, mineral interactions that affect acid-base and antioxidant status, and diet considerations to improve mineral status. J. Dairy Sci. 2018, 101, 2763–2813. [Google Scholar] [CrossRef]
- López-Alonso, M.; Miranda, M. Copper Supplementation, A Challenge in Cattle. Animals 2020, 10, 1890. [Google Scholar] [CrossRef]
- Muszyński, S.; Tomaszewska, E.; Kwiecień, M.; Dobrowolski, P.; Tomczyk, A. Effect of Dietary Phytase Supplementation on Bone and Hyaline Cartilage Development of Broilers Fed with Organically Complexed Copper in a Cu-Deficient Diet. Biol. Trace Elem. Res. 2018, 182, 339–353. [Google Scholar] [CrossRef] [Green Version]
- Ashmead, H.D. Comparative intestinal absorption and subsequent metabolism of metal amino acid chelates and inorganic metal salts. In Biological Trace Element Research; Subramanian, K.S., Iyengar, G.K., Okamoto, K., Eds.; American Chemical Society: Washington, DC, USA, 1991; pp. 306–319. [Google Scholar]
- Aldridge, B.E.; Saddoris, K.L.; Radcliffe, J.S. Copper can be absorbed as a Cu-peptide chelate through the PepT1 transporter in the jejunum of weanling pigs. J. Anim. Sci. 2007, 85, 154–155. [Google Scholar]
- Sauer, A.K.; Pfaender, S.; Hagmeyer, S.; Tarana, L.; Mattes, A.-K.; Briel, F.; Küry, S.; Boeckers, T.M.; Grabrucker, A.M. Characterization of zinc amino acid complexes for zinc delivery in vitro using Caco-2 cells and enterocytes from hiPSC. Biometals 2017, 30, 643–661. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Hemken, R.W.; Harmon, R.J. Copper Metabolism of Holstein and Jersey Cows and Heifers Fed Diets High in Cupric Sulfate or Copper Proteinate. J. Dairy Sci. 1996, 79, 1873–1880. [Google Scholar] [CrossRef]
- Glover, C.N.; Wood, C.M. Absorption of copper and copper-histidine complexes across the apical surface of freshwater rainbow trout intestine. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2008, 178, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Souba, W.W.; Pacitti, A.J. How amino acids get into cells: Mechanisms, models, menus, and mediators. JPEN J. Parenter. Enter. Nutr. 1992, 16, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Bröer, S. Amino Acid Transport Across Mammalian Intestinal and Renal Epithelia. Physiol. Rev. 2008, 88, 249–286. [Google Scholar] [CrossRef] [PubMed]
- Verrey, F.; Singer, D.; Ramadan, T.; Vuille-dit-Bille, R.N.; Mariotta, L.; Camargo, S.M.R. Kidney amino acid transport. PflÜGers Arch. Eur. J. Physiol. 2009, 458, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Wong, F.H.; Chen, J.S.; Reddy, V.; Day, J.L.; Shlykov, M.A.; Wakabayashi, S.T.; Saier, M.H., Jr. The amino acid-polyamine-organocation superfamily. J. Mol. Microbiol. Biotechnol. 2012, 22, 105–113. [Google Scholar] [CrossRef]
- Palacin, M.; Estevez, R.; Bertran, J.; Zorzano, A. Molecular Biology of Mammalian Plasma Membrane Amino Acid Transporters. Physiol. Rev. 1998, 78, 969–1054. [Google Scholar] [CrossRef]
- Saier, M.H.; Daniels, G.A.; Boerner, P.; Lin, J. Neutral amino acid transport systems in animal cells: Potential targets of oncogene action and regulators of cellular growth. J. Membr. Biol. 1988, 104, 1–20. [Google Scholar] [CrossRef]
- Webb, K.E., Jr. Intestinal absorption of protein hydrolysis products: A review. J. Anim. Sci. 1990, 68, 3011–3022. [Google Scholar] [CrossRef] [Green Version]
- Stevens, B.R.; Kaunitz, J.D.; Wright, E.M. Intestinal transport of amino acids and sugars: Advances using membrane vesicles. Annu. Rev. Physiol. 1984, 46, 417–433. [Google Scholar] [CrossRef]
- Frenhani, P.B.; Burini, R.C. Mechanisms of absorption of amino acids and oligopeptides. Control and implications in human diet therapy. Arq. Gastroenterol. 1999, 36, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Yin, T.; Xu, B.; Ma, Y.; Hu, M. Amino acid facilitates absorption of copper in the Caco-2 cell culture model. Life Sci. 2014, 109, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wong, E.A.; Gilbert, E.R. Bioavailability of different dietary supplemental methionine sources in animals. Front. Biosci. Elite 2015, 7, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Romanet, S.; Aschenbach, J.R.; Pieper, R.; Zentek, J.; Htoo, J.K.; Whelan, R.A.; Mastrototaro, L. Expression of proposed methionine transporters along the gastrointestinal tract of pigs and their regulation by dietary methionine sources. Genes Nutr. 2021, 16, 14. [Google Scholar] [CrossRef] [PubMed]
- To, V.P.T.H.; Masagounder, K.; Loewen, M.E. Critical transporters of methionine and methionine hydroxy analogue supplements across the intestine: What we know so far and what can be learned to advance animal nutrition. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2021, 255, 110908. [Google Scholar] [CrossRef]
- Brachet, P.; Puigserver, A. Transport of Methionine Hydroxy Analog across the Brush Border Membrane of Rat Jejunum. J. Nutr. 1987, 117, 1241–1246. [Google Scholar] [CrossRef] [Green Version]
- Stein, J.; Zores, M.; Schröder, O. Short-chain fatty acid (SCFA) uptake into Caco-2 cells by a pH-dependent and carrier mediated transport mechanism. Eur. J. Nutr. 2000, 39, 121–125. [Google Scholar] [CrossRef]
- Fafournoux, P.; Rémésy, C.; Demigné, C. Propionate transport in rat liver cells. Biochim. Biophys. Acta (BBA) Biomembr. 1985, 818, 73–80. [Google Scholar] [CrossRef]
- Stumpff, F. A look at the smelly side of physiology: Transport of short chain fatty acids. PflÜGers Arch. Eur. J. Physiol. 2018, 470, 571–598. [Google Scholar] [CrossRef]
- Adibi, S.A. Intestinal transport of dipeptides in man: Relative importance of hydrolysis and intact absorption. J. Clin. Investig. 1971, 50, 2266–2275. [Google Scholar] [CrossRef] [Green Version]
- Craft, I.L.; Geddes, D.; Hyde, C.W.; Wise, I.J.; Matthews, D.M. Absorption and malabsorption of glycine and glycine peptides in man. Gut 1968, 9, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Cheng, B.; Navab, F.; Lis, M.T.; Miller, T.N.; Matthews, D.M. Mechanisms of dipeptide uptake by rat small intestine in vitro. Clin. Sci. 1971, 40, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Burston, D.; Addison, J.M.; Matthews, D.M. Uptake of dipeptides containing basic and acidic amino acids by rat small intestine in vitro. Clin. Sci. 1972, 43, 823–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, H.; Funabiki, R.; Iwata, M.; Yamazaki, K. Portal absorption of small peptides in rats under unrestrained conditions. J. Nutr. 1984, 114, 1122–1129. [Google Scholar] [CrossRef]
- Silk, D.B.; Fairclough, P.D.; Clark, M.L.; Hegarty, J.E.; Marrs, T.C.; Addison, J.M.; Burston, D.; Clegg, K.M.; Matthews, D.M. Use of a peptide rather than free amino acid nitrogen source in chemically defined “elemental” diets. JPEN J. Parenter. Enter. Nutr. 1980, 4, 548–553. [Google Scholar] [CrossRef]
- Asatoor, A.M.; Cheng, B.; Edwards, K.D.; Lant, A.F.; Matthews, D.M.; Milne, M.D.; Navab, F.; Richards, A.J. Intestinal absorption of dipeptides and corresponding free amino acids in Hartnup disease. Clin. Sci. 1970, 39, 1P. [Google Scholar] [CrossRef]
- Hellier, M.D.; Holdsworth, C.D.; Perrett, D.; Thirumalai, C. Intestinal depeptide transport in normal and cystinuric subjects. Clin. Sci. 1972, 43, 659–668. [Google Scholar] [CrossRef]
- Sun, X.; Acquah, C.; Aluko, R.E.; Udenigwe, C.C. Considering food matrix and gastrointestinal effects in enhancing bioactive peptide absorption and bioavailability. J. Funct. Foods 2020, 64, 103680. [Google Scholar] [CrossRef]
- Wada, Y.; Lonnerdal, B. Bioactive peptides derived from human milk proteins—Mechanisms of action. J. Nutr. Biochem. 2013, 25, 503–514. [Google Scholar] [CrossRef]
- Doguer, C.; Ha, J.-H.; Collins, J.F. Intersection of Iron and Copper Metabolism in the Mammalian Intestine and Liver. Compr. Physiol. 2018, 8, 1433–1461. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Wu, X.; Lee, J. SLC31 (CTR) family of copper transporters in health and disease. Mol. Asp. Med. 2013, 34, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Puchkova, L.V.; Broggini, M.; Polishchuk, E.V.; Ilyechova, E.Y.; Polishchuk, R.S. Silver Ions as a Tool for Understanding Different Aspects of Copper Metabolism. Nutrients 2019, 11, 1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Berghe, P.; Klomp, L. New Developments in the regulation of intestinal copper absorption. Nutr. Rev. 2009, 67, 658–672. [Google Scholar] [CrossRef] [PubMed]
- Fry, R.S.; Spears, J.W.; Lloyd, K.E.; O’Nan, A.T.; Ashwell, M.S. Effect of dietary copper and breed on gene products involved in copper acquisition, distribution, and use in Angus and Simmental cows and fetuses. J. Anim. Sci. 2013, 91, 861–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hambidge, K.M.; Casey, C.E.; Krebs, N.F. 1-Zinc. In Trace Elements in Human and Animal Nutrition, 5th ed.; Mertz, W., Ed.; Academic Press: San Diego, CA, USA, 1986; pp. 1–137. [Google Scholar] [CrossRef]
- O’Dell, B.L.; Sunde, R.A. Handbook of Nutritionally Essential Mineral Elements; CRC Press: New York, NY, USA, 1997. [Google Scholar]
- Curnock, R.; Cullen, P.J. Mammalian copper homeostasis requires retromer-dependent recycling of the high-affinity copper transporter 1. J. Cell Sci. 2020, 133, jcs249201. [Google Scholar] [CrossRef]
- Spears, J.W. Advancements in Ruminant Trace Mineral Nutrition. In Proceedings of the Cornell Nutrition Conference for Feed Manufacturers, New York, NY, USA, 22–24 October 2013. [Google Scholar]
- Kim, B.E.; Nevitt, T.; Thiele, D.J. Mechanisms for copper acquisition, distribution and regulation. Nat. Chem. Biol. 2008, 4, 176–185. [Google Scholar] [CrossRef]
- Conrad, M.E.; Umbreit, J.N. Pathways of iron absorption. Blood Cells Mol. Dis. 2002, 29, 336–355. [Google Scholar] [CrossRef]
- Mackenzie, B.; Garrick, M.D. Iron Imports. II. Iron uptake at the apical membrane in the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G981–G986. [Google Scholar] [CrossRef] [Green Version]
- De Domenico, I.; McVey Ward, D.; Kaplan, J. Regulation of iron acquisition and storage: Consequences for iron-linked disorders. Nat. Rev. Mol. Cell Biol. 2008, 9, 72–81. [Google Scholar] [CrossRef]
- Duck, K.A.; Connor, J.R. Iron uptake and transport across physiological barriers. Biometals 2016, 29, 573–591. [Google Scholar] [CrossRef] [Green Version]
- Gozzelino, R.; Arosio, P. Iron Homeostasis in Health and Disease. Int. J. Mol. Sci. 2016, 17, 130. [Google Scholar] [CrossRef] [Green Version]
- Sangkhae, V.; Nemeth, E. Regulation of the Iron Homeostatic Hormone Hepcidin. Adv. Nutr. 2017, 8, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Zhang, K.; Ding, X.; Wang, J.; Zeng, Q.; Peng, H.; Bai, J.; Xuan, Y.; Su, Z.; Wu, B. Uptake of Manganese from the Manganese-Lysine Complex in Primary Chicken Intestinal Epithelial Cells. Animals 2019, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cousins, R.J.; Liuzzi, J.P. Chapter 61—Trace Metal Absorption and Transport. In Physiology of the Gastrointestinal Tract, 6th ed.; Said, H.M., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 1485–1498. [Google Scholar] [CrossRef]
- Jenkitkasemwong, S.; Wang, C.-Y.; Mackenzie, B.; Knutson, M.D. Physiologic implications of metal-ion transport by ZIP14 and ZIP8. Biometals 2012, 25, 643–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopalsamy, G.; Alpers, D.; Binder, H.; Tran, C.; Ramakrishna, B.; Brown, I.; Manary, M.; Mortimer, E.; Young, G. The Relevance of the Colon to Zinc Nutrition. Nutrients 2015, 7, 572. [Google Scholar] [CrossRef]
- Cousins, R.J.; Liuzzi, J.P.; Lichten, L.A. Mammalian Zinc Transport, Trafficking, and Signals. J. Biol. Chem. 2006, 281, 24085–24089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maares, M.; Haase, H. A Guide to Human Zinc Absorption: General Overview and Recent Advances of In Vitro Intestinal Models. Nutrients 2020, 12, 762. [Google Scholar] [CrossRef] [Green Version]
- Eide, D.J. Zinc transporters and the cellular trafficking of zinc. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2006, 1763, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Yuzbasiyan-Gurkan, V.; Bartlett, E. Identification of a unique splice site variant in SLC39A4 in bovine hereditary zinc deficiency, lethal trait A46: An animal model of acrodermatitis enteropathica. Genomics 2006, 88, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Condomina, J.; Zornoza-Sabina, T.; Granero, L.; Polache, A. Kinetics of zinc transport in vitro in rat small intestine and colon: Interaction with copper. Eur. J. Pharm. Sci. 2002, 16, 289–295. [Google Scholar] [CrossRef]
- Hashimoto, A.; Kambe, T. Mg, Zn and Cu Transport Proteins: A Brief Overview from Physiological and Molecular Perspectives. J. Nutr. Sci. Vitaminol. 2015, 61, S116–S118. [Google Scholar] [CrossRef] [Green Version]
- Lutsenko, S.; Barnes, N.L.; Bartee, M.Y.; Dmitriev, O.Y. Function and Regulation of Human Copper-Transporting ATPases. Physiol. Rev. 2007, 87, 1011–1046. [Google Scholar] [CrossRef] [PubMed]
- Zimnicka, A.M.; Ivy, K.; Kaplan, J.H. Acquisition of dietary copper: A role for anion transporters in intestinal apical copper uptake. Am. J. Physiol. Cell Physiol. 2011, 300, C588–C599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cousins, R.J. Gastrointestinal factors influencing zinc absorption and homeostasis. Int. J. Vitam. Nutr. Res. 2010, 80, 243–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Dell, B.L. Bioavailability of essential and toxic trace elements. Introduction. Fed. Proc. 1983, 42, 1714–1715. [Google Scholar]
- Littell, R.C.; Lewis, A.J.; Henry, P.R. 1—Statistical evaluation of bioavailability assays. In Bioavailability of Nutrients for Animals; Ammerman, C.B., Baker, D.H., Lewis, A.J., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 5–33. [Google Scholar] [CrossRef]
- Miller, E.R. Techniques for determining bioavailability of trace elements. In Proceedings of the Sixth Annual International Minerals Conferene, St. Petersburg Beach, FL, USA; 1983. [Google Scholar]
- Ledoux, D.R.; Shannon, M.C. Bioavailability and Antagonists of Trace Minerals in Ruminant Metabolism. In Proceedings of the Florida Ruminant Symposium, Gainesville, FL, USA, 1–2 February 2005. [Google Scholar]
- Miles, R.D.; Henry, P.R. Relative trace mineral bioavailability. Braz. Anim. Sci. 2006, 1, 73–93. [Google Scholar]
- De Groote, G.; Lippens, M.; Jongbloed, A.W.; Meschy, F. Study on the Bioavailability of Major and Trace Minerals; International Association of the European (EU) Manufacturers of Major, Trace and Specific Feed Mineral Materials (Emfema): Brussels, Belgium, 2002. [Google Scholar]
- Baker, D.H.; Odle, J.; Funk, M.A.; Wieland, T.M. Research note: Bioavailability of copper in cupric oxide, cuprous oxide, and in a copper-lysine complex. Poult. Sci. 1991, 70, 177–179. [Google Scholar] [CrossRef]
- Fairweather-Tait, S.J. The concept of bioavailability as it relates to iron nutrition. Nutr. Res. 1987, 7, 319–325. [Google Scholar] [CrossRef]
- Ledoux, D.R.; Pott, E.B.; Henry, P.R.; Ammerman, C.B.; Merritt, A.M.; Madison, J.B. Estimation of the relative bioavailability of inorganic copper sources for sheep. Nutr. Res. 1995, 15, 1803–1813. [Google Scholar] [CrossRef]
- Ledoux, D.R.; Henry, P.R.; Ammerman, C.B.; Rao, P.V.; Miles, R.D. Estimation of the relative bioavailability of inorganic copper sources for chicks using tissue uptake of copper. J. Anim. Sci. 1991, 69, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Kegley, E.B.; Spears, J.W. Bioavailability of feed-grade copper sources (oxide, sulfate, or lysine) in growing cattle. J. Anim. Sci. 1994, 72, 2728–2734. [Google Scholar] [CrossRef]
- Edwards, H.M.; Baker, D.H. Bioavailability of zinc in several sources of zinc oxide, zinc sulfate, and zinc metal. J. Anim. Sci. 1999, 77, 2730–2735. [Google Scholar] [CrossRef]
- Ammerman, C.B.; Baker, D.H.; Lewis, A.J. (Eds.) Introduction. In Bioavailability of Nutrients for Animals; Academic Press: San Diego, CA, USA, 1995; pp. 1–3. [Google Scholar] [CrossRef]
- L’Abbé, M.R.; Fischer, P.W. The effects of high dietary zinc and copper deficiency on the activity of copper-requiring metalloenzymes in the growing rat. J. Nutr. 1984, 114, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Suttle, N.F. A technique for measuring the biological availability of copper to sheep, using hypocupraemic ewes. Br. J. Nutr. 1974, 32, 395–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, R.L.; Southern, L.L. Changes in glutathione peroxidase and tissue selenium concentrations of broilers after consuming a diet adequate in selenium. Poult. Sci. 2005, 84, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.D.; Shirley, R.B.; Winkelbauer, P.D.A.; Atwell, C.A.; Wuelling, C.W.; Wehmeyer, M.; Buttin, P. Bioavailability of zinc sources in chickens determined via real time polymerase chain reaction (RT-PCR) assay for metallothionein. In Proceedings of the 16th European Symposium on Poultry Nutrition, World Poultry Science Association, Strasbourg, France, 26–30 August 2007. [Google Scholar]
- Huang, Y.L.; Lu, L.; Li, S.F.; Luo, X.G.; Liu, B. Relative bioavailabilities of organic zinc sources with different chelation strengths for broilers fed a conventional corn-soybean meal diet. J. Anim. Sci. 2009, 87, 2038–2046. [Google Scholar] [CrossRef] [Green Version]
- Wedekind, K.J.; Lewis, A.J.; Giesemann, M.A.; Miller, P.S. Bioavailability of zinc from inorganic and organic sources for pigs fed corn-soybean meal diets. J. Anim. Sci. 1994, 72, 2681–2689. [Google Scholar] [CrossRef]
- Schell, T.C.; Kornegay, E.T. Zinc concentration in tissues and performance of weanling pigs fed pharmacological levels of zinc from ZnO, Zn-methionine, Zn-lysine, or ZnSO4. J. Anim. Sci. 1996, 74, 1584–1593. [Google Scholar] [CrossRef]
- Wedekind, K.J.; Hortin, A.E.; Baker, D.H. Methodology for assessing zinc bioavailability: Efficacy estimates for zinc-methionine, zinc sulfate, and zinc oxide. J. Anim. Sci. 1992, 70, 178–187. [Google Scholar] [CrossRef]
- Sandoval, M.; Henry, P.R.; Ammerman, C.B.; Miles, R.D.; Littell, R.C. Relative bioavailability of supplemental inorganic zinc sources for chicks. J. Anim. Sci. 1997, 75, 3195–3205. [Google Scholar] [CrossRef] [Green Version]
- Cano-Sancho, G.; Rovira, J.; Perelló, G.; Martorell, I.; Tous, N.; Nadal, M.; Domingo, J.L. Extensive Literature Search on the bioavailability of selected trace elements in animal nutrition: Incompatibilities and interactions. EFSA Supporting Publ. 2014, 11, 565E. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.F.; Zeringue, L.K. Laboratory evaluations of solubility and structural integrity of complexed and chelated trace mineral supplements. J. Dairy Sci. 1994, 77, 181–189. [Google Scholar] [CrossRef]
- Cao, J.; Henry, P.R.; Guo, R.; Holwerda, R.A.; Toth, J.P.; Littell, R.C.; Miles, R.D.; Ammerman, C.B. Chemical characteristics and relative bioavailability of supplemental organic zinc sources for poultry and ruminants. J. Anim. Sci. 2000, 78, 2039–2054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, R.; Henry, P.R.; Holwerda, R.; Cao, J.; Littell, R.C.; Miles, R.D.; Ammerman, C.B. Chemical characteristics and relative bioavailability of supplemental organic copper sources for poultry. J. Anim. Sci. 2001, 79, 1132–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udechukwu, M.C.; Collins, S.A.; Udenigwe, C.C. Prospects of enhancing dietary zinc bioavailability with food-derived zinc-chelating peptides. Food Funct. 2016, 7, 4137–4144. [Google Scholar] [CrossRef] [PubMed]
- Caetano-Silva, M.; Cilla, A.; Pacheco, M.T.; Netto, F.; Alegría, A. Evaluation of in vitro iron bioavailability in free form and as whey peptide-iron complexes. J. Food Compos. Anal. 2018, 68, 95–100. [Google Scholar] [CrossRef]
- Scheers, N.M.; Sandberg, A.-S. Ascorbic acid uptake affects ferritin, Dcytb and Nramp2 expression in Caco-2 cells. Eur. J. Nutr. 2008, 47, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, P.; Sauvant, D.; Jondreville, C. Bioavailability of zinc sources and their interaction with phytates in broilers and piglets. Animal 2013, 7, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Sauvant, D.; Perez, J.M.; Tran, G. Tables of Composition and Nutritional Value of Feed Materials: Pig, Poultry, Sheep, Goats, Rabbits, Horses, Fish; Wagening Academic Publishers: Wageningen, The Netherlands, 2004. [Google Scholar] [CrossRef]
- Susaki, H.; Matsui, T.; Kinya, A.; Fujita, S.; Nakajima, T.; Yano, H. Availability of a Zinc Amino Acid Chelate for Growing Pigs. Nihon Chikusan Gakkaiho 1999, 70, 124–128. [Google Scholar] [CrossRef] [Green Version]
- Zacharias, B.; Pelletier, W.; Drochner, W. Availability of inorganic and organic bound copper and zinc fed at physiological levels to fattening pigs. Zemes Ukio Mokslai 2007, 14, 45–50. [Google Scholar]
- Richards, J.; Fisher, P.; Evans, J.; Wedekind, K. Greater bioavailability of chelated compared with inorganic zinc in broiler chicks in the presence or absence of elevated calcium and phosphorus. Open Access Anim. Physiol. 2015, 7, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Jondreville, C.; Lescoat, P.; Magnin, M.; Feuerstein, D.; Gruenberg, B.; Nys, Y. Sparing effect of microbial phytase on zinc supplementation in maize–soya-bean meal diets for chickens. Animal 2007, 1, 804–811. [Google Scholar] [CrossRef] [Green Version]
- Richards, J.; Zhao, J.; Harrell, R.; Atwell, C.; Dibner, J. Trace Mineral Nutrition in Poultry and Swine. Asian-Australas. J. Anim. Sci. 2010, 23, 1527–1534. [Google Scholar] [CrossRef]
- Nockels, C.F.; DeBonis, J.; Torrent, J. Stress induction affects copper and zinc balance in calves fed organic and inorganic copper and zinc sources. J. Anim. Sci. 1993, 71, 2539–2545. [Google Scholar] [CrossRef] [PubMed]
- Parks, F.P.a.K.J.H. An assay method from proteinates: Judging organic trace minerals. Feed Manag. 1994, 45, 35–38. [Google Scholar]
- Khatun, A.; Chowdhury, S.D.; Roy, B.C.; Dey, B.; Haque, A.; Chandran, B. Comparative effects of inorganic and three forms of organic trace minerals on growth performance, carcass traits, immunity, and profitability of broilers. J. Adv. Vet. Anim. Res. 2019, 6, 66–73. [Google Scholar] [CrossRef]
- Savaram Venkata, R.R.; Bhukya, P.; Raju, M.V.L.N.; Ullengala, R. Effect of Dietary Supplementation of Organic Trace Minerals at Reduced Concentrations on Performance, Bone Mineralization, and Antioxidant Variables in Broiler Chicken Reared in Two Different Seasons in a Tropical Region. Biol. Trace Elem. Res. 2021, 199, 3817–3824. [Google Scholar] [CrossRef]
- Wang, G.; Liu, L.; Wang, Z.; Pei, X.; Tao, W.; Xiao, Z.; Liu, B.; Wang, M.; Lin, G.; Ao, T. Comparison of Inorganic and Organically Bound Trace Minerals on Tissue Mineral Deposition and Fecal Excretion in Broiler Breeders. Biol. Trace Elem. Res. 2019, 189, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Liu, L.J.; Tao, W.J.; Xiao, Z.P.; Pei, X.; Liu, B.J.; Wang, M.Q.; Lin, G.; Ao, T.Y. Effects of replacing inorganic trace minerals with organic trace minerals on the production performance, blood profiles, and antioxidant status of broiler breeders. Poult. Sci. 2019, 98, 2888–2895. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Xiong, P.; Chen, N.; He, J.; Lin, G.; Xue, Y.; Li, W.; Yu, D. Effects of Replacing of Inorganic Trace Minerals by Organically Bound Trace Minerals on Growth Performance, Tissue Mineral Status, and Fecal Mineral Excretion in Commercial Grower-Finisher Pigs. Biol. Trace Elem. Res. 2016, 173, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Araújo, C.S.S.; Hermes, R.G.; Bittencourt, L.C.; Silva, C.C.; Araújo, L.F.; Granghelli, C.A.; Pelissari, P.H.; Roque, F.A.; Leite, B.G.S. Different dietary trace mineral sources for broiler breeders and their progenies. Poult. Sci. 2019, 98, 4716–4721. [Google Scholar] [CrossRef]
- Kincaid, R.L.; Blauwiekel, R.M.; Cronrath, J.D. Supplementation of Copper as Copper Sulfate or Copper Proteinate for Growing Calves Fed Forages Containing Molybdenum. J. Dairy Sci. 1986, 69, 160–163. [Google Scholar] [CrossRef]
- Ward, J.D.; Spears, J.W.; Kegley, E.B. Bioavailability of copper proteinate and copper carbonate relative to copper sulfate in cattle. J. Dairy Sci. 1996, 79, 127–132. [Google Scholar] [CrossRef]
- Nollet, L.; van der Klis, J.D.; Lensing, M.; Spring, P. The Effect of Replacing Inorganic With Organic Trace Minerals in Broiler Diets on Productive Performance and Mineral Excretion. J. Appl. Poult. Res. 2007, 16, 592–597. [Google Scholar] [CrossRef]
- Spears, J.W. Zinc Methionine for Ruminants: Relative Bioavailability of Zinc in Lambs and Effects of Growth and Performance of Growing Heifers. J. Anim. Sci. 1989, 67, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Luo, X.; Liu, B.; Crenshaw, T.D.; Kuang, X.; Shao, G.; Yu, S. Use of chemical characteristics to predict the relative bioavailability of supplemental organic manganese sources for broilers. J. Anim. Sci. 2004, 82, 2352–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elashry, G. Effect of Feeding a Combination of Zinc, Manganese and Copper Methionine Chelates of Early Lactation High Producing Dairy Cow. Food Nutr. Sci. 2012, 3, 1084–1091. [Google Scholar] [CrossRef] [Green Version]
- Nocek, J.E.; Socha, M.T.; Tomlinson, D.J. The Effect of Trace Mineral Fortification Level and Source on Performance of Dairy Cattle. J. Dairy Sci. 2006, 89, 2679–2693. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, L.M.; Loeffler, S.H.; Socha, M.T.; Tomlinson, D.J.; Johnson, A.B. Effects of supplementing complexed zinc, manganese, copper and cobalt on lactation and reproductive performance of intensively grazed lactating dairy cattle on the South Island of New Zealand. Anim. Feed Sci. Technol. 2007, 137, 69–83. [Google Scholar] [CrossRef]
- Siciliano-Jones, J.L.; Socha, M.; Tomlinson, D.; Defrain, J. Effect of Trace Mineral Source on Lactation Performance, Claw Integrity, and Fertility of Dairy Cattle. J. Dairy Sci. 2008, 91, 1985–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackbart, K.S.; Ferreira, R.M.; Dietsche, A.A.; Socha, M.T.; Shaver, R.D.; Wiltbank, M.C.; Fricke, P.M. Effect of dietary organic zinc, manganese, copper, and cobalt supplementation on milk production, follicular growth, embryo quality, and tissue mineral concentrations in dairy cows. J. Anim. Sci. 2010, 88, 3856–3870. [Google Scholar] [CrossRef] [Green Version]
- Rabiee, A.R.; Lean, I.J.; Stevenson, M.A.; Socha, M.T. Effects of feeding organic trace minerals on milk production and reproductive performance in lactating dairy cows: A meta-analysis. J. Dairy Sci. 2010, 93, 4239–4251. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, D.W.; Tomlinson, D.J.; Socha, M.T.; Johnson, A.B. Effects of Zinc Methionine Complex on Milk Production and Somatic Cell Count of Dairy Cows: Twelve-Trial Summary. Prof. Anim. Sci. 2004, 20, 295–301. [Google Scholar] [CrossRef]
- Rossi, C.; Grossi, S.; Compiani, R.; Baldi, G.; Agovino, M.; Rossi, L. Effects of different mineral supplementation programs on beef cattle serum Se, Zn, Cu, Mn concentration, health, growth performance and meat quality. Large Anim. Rev. 2020, 26, 57–64. [Google Scholar]
- Price, D.; Arellano, K.; Irsik, M.; Rae, D.O.; Yelich, J.; Mjoun, K.; Hersom, M. Effects of trace mineral supplement source during gestation and lactation in Angus and Brangus cows and subsequent calf immunoglobulin concentrations, growth, and development. Prof. Anim. Sci. 2017, 33, 194–204. [Google Scholar] [CrossRef]
- Cope, C.; Mackenzie, A.M.; Wilde, D.; Sinclair, L.A. Effects of level and form of dietary zinc on dairy cattle performance and keratin production in the teat canal. Proc. Br. Soc. Anim. Sci. 2009, 2009, 85. [Google Scholar] [CrossRef]
- Pino, F.; Urrutia, N.; Gelsinger, S.L.; Gehman, A.M.; Heinrichs, A. Long-term effect of organic trace minerals on growth, reproductive performance, and first lactation in dairy heifers. Prof. Anim. Sci. 2018, 34, 51–58. [Google Scholar] [CrossRef]
- Pino, F.; Heinrichs, A.J. Effect of trace minerals and starch on digestibility and rumen fermentation in diets for dairy heifers. J. Dairy Sci. 2016, 99, 2797–2810. [Google Scholar] [CrossRef] [Green Version]
- Scaletti, R.W.; Harmon, R.J. Effect of dietary copper source on response to coliform mastitis in dairy cows. J. Dairy Sci. 2012, 95, 654–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, D.H.; Ammerman, C.B. 7—Copper bioavailability. In Bioavailability of Nutrients for Animals; Ammerman, C.B., Baker, D.H., Lewis, A.J., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 127–156. [Google Scholar] [CrossRef]
- Xin, Z.; Waterman, D.F.; Hemken, R.W.; Harmon, R.J.; Jackson, J.A. Effects of Copper Sources and Dietary Cation-Anion Balance on Copper Availability and Acid-Base Status in Dairy Calves1. J. Dairy Sci. 1991, 74, 3167–3173. [Google Scholar] [CrossRef]
- Xin, Z.; Waterman, D.F.; Hemken, R.W.; Harmon, R.J. Effects of Copper Status on Neutrophil Function, Superoxide Dismutase, and Copper Distribution in Steers1. J. Dairy Sci. 1991, 74, 3078–3085. [Google Scholar] [CrossRef]
- Lee, D.-Y.; Schroeder, J., III; Gordon, D.T. Enhancement of Cu Bioavailability in the Rat by Phytic Acid. J. Nutr. 1988, 118, 712–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spears, J.W.; Kegley, E.B.; Mullis, L.A.; Wise, T.A. Bioavailability of copper from tri-basic copper chloride in cattle. J. Anim. Sci. 1997, 75, 265. [Google Scholar]
- Arthington, J.D.; Pate, F.M.; Spears, J.W. Effect of copper source and level on performance and copper status of cattle consuming molasses-based supplements. J. Anim. Sci. 2003, 81, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.D.; Spears, J.W.; Kegley, E.B. Effect of copper level and source (copper lysine vs copper sulfate) on copper status, performance, and immune response in growing steers fed diets with or without supplemental molybdenum and sulfur. J. Anim. Sci. 1993, 71, 2748–2755. [Google Scholar] [CrossRef]
- Hansen, S.L.; Schlegel, P.; Legleiter, L.R.; Lloyd, K.E.; Spears, J.W. Bioavailability of copper from copper glycinate in steers fed high dietary sulfur and molybdenum. J. Anim. Sci. 2008, 86, 173–179. [Google Scholar] [CrossRef]
- Genther, O.N.; Hansen, S.L. The effect of trace mineral source and concentration on ruminal digestion and mineral solubility. J. Dairy Sci. 2015, 98, 566–573. [Google Scholar] [CrossRef]
- Arthington, J.D.; Spears, J.W. Effects of tribasic copper chloride versus copper sulfate provided in corn-and molasses-based supplements on forage intake and copper status of beef heifers. J. Anim. Sci. 2007, 85, 871–876. [Google Scholar] [CrossRef] [Green Version]
- Spears, J.; Kegley, E.; Mullis, L. Bioavailability of copper from tribasic copper chloride and copper sulfate in growing cattle. Anim. Feed Sci. Technol. 2004, 116, 1–13. [Google Scholar] [CrossRef]
- Sinclair, L.A.; Hart, K.J.; Johnson, D.; Mackenzie, A.M. Effect of inorganic or organic copper fed without or with added sulfur and molybdenum on the performance, indicators of copper status, and hepatic mRNA in dairy cows. J. Dairy Sci. 2013, 96, 4355–4367. [Google Scholar] [CrossRef] [Green Version]
- van den Top, A.M. Reviews on the Mineral Provision in Ruminants (IX): Copper Metabolism and Requirements in Ruminants; Centraal Veevoeder Bureau: Lelystad, The Netherlands, 2005. [Google Scholar]
- Hemken, R.W. Use of Copper Proteinates and Copper Lysine in Animal Feeding Programs. 1997. Available online: https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.585.9306&rep=rep1&type=pdf (accessed on 31 October 2021).
- McGuire, S.O.; Miller, W.J.; Gentry, R.P.; Neathery, M.W.; Ho, S.Y.; Blackmon, D.M. Influence of high dietary iron as ferrous carbonate and ferrous sulfate on iron metabolism in young calves. J. Dairy Sci. 1985, 68, 2621–2628. [Google Scholar] [CrossRef]
- Bremner, I.; Dalgarno, A.C. Iron metabolism in the veal calf. The availability of different iron compounds. Br. J. Nutr. 1973, 29, 229–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neathery, M.W.; Rachmat, S.; Miller, W.J.; Gentry, R.P.; Blackmon, D.M. Effect of Chemical Form of Orally Administered 65Zn on Absorption and Metabolism in Cattle. Proc. Soc. Exp. Biol. Med. 1972, 139, 953–956. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.W.; Craig, W.M.; Southern, L.L. Ruminal distribution of zinc in steers fed a polysaccharide-zinc complex or zinc oxide. J. Anim. Sci. 1993, 71, 1281–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, D.W.; Craig, W.M.; Southern, L.L. Southern and M. Engstrom. Ruminal partitioning of zinc in steers fed a polysaccharide complex of zinc or zinc oxide. J. Anim. Sci. 1988, 66, 462. [Google Scholar]
- Kincaid, R.L.; Hodgson, A.S.; Riley, R.E., Jr.; Conrath, J.D. Supplementation of diets for lactating cows with zinc as zinc oxide and zinc methionine. J. Dairy Sci. 1984, 67, 103. [Google Scholar]
- Kincaid, R.L. Biological availability of zinc from inorganic sources with excess dietary calcium. J. Dairy Sci. 1979, 62, 1081–1085. [Google Scholar] [CrossRef]
- Wright, C.L.; Spears, J.W. Effect of Zinc Source and Dietary Level on Zinc Metabolism in Holstein Calves. J. Dairy Sci. 2004, 87, 1085–1091. [Google Scholar] [CrossRef] [Green Version]
- Spears, J.; Kegley, E. Effect of zinc source (zinc oxide vs zinc proteinate) and level on performance, carcass characteristics, and immune response of growing and finishing steers. J. Anim. Sci. 2002, 80, 2747–2752. [Google Scholar] [CrossRef]
- Kerley, M.S.; Ledoux, D.R. Practical applications for chelated minerals in cattle. In Proceedings of the National Feed Ingredient Association Nutrition Institute, Des Moines, IA, USA; 1992; pp. 1–12. [Google Scholar]
- Cope, C.M.; Mackenzie, A.M.; Wilde, D.; Sinclair, L.A. Effects of level and form of dietary zinc on dairy cow performance and health. J. Dairy Sci. 2009, 92, 2128–2135. [Google Scholar] [CrossRef] [Green Version]
- Stewart, W.C.; Scasta, J.D.; Taylor, J.B.; Murphy, T.W.; Julian, A.A.M. Invited Review: Mineral nutrition considerations for extensive sheep production systems. Appl. Anim. Sci. 2021, 37, 256–272. [Google Scholar] [CrossRef]
- Spears, J. Trace Mineral Bioavailability in Ruminants. J. Nutr. 2003, 133, 1506S–1509S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Fan, C.; Zhang, W.; Zhu, X.; Yan, X.; Wang, R.; Jia, Z. Effects of dietary copper source and level on performance, carcass characteristics and lipid metabolism in lambs. Asian-Australas. J. Anim. Sci. 2008, 21, 685–691. [Google Scholar] [CrossRef]
- Pott, E.B.; Henry, P.R.; Ammerman, C.B.; Merritt, A.M.; Madison, J.B.; Miles, R.D. Relative bioavailability of copper in a copperlysine complex for chicks and lambs. Anim. Feed Sci. Technol. 1994, 45, 193–203. [Google Scholar] [CrossRef]
- Pal, D.T.; Gowda, N.K.; Prasad, C.S.; Amarnath, R.; Bharadwaj, U.; Suresh Babu, G.; Sampath, K.T. Effect of copper- and zinc-methionine supplementation on bioavailability, mineral status and tissue concentrations of copper and zinc in ewes. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. (GMS) 2010, 24, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Eckert, G.; Greene, L.; Carstens, G.; Ramsey, W. Copper status of ewes fed increasing amounts of copper from copper sulfate or copper proteinate. J. Anim. Sci. 1999, 77, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Council, N.R. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; The National Academies Press: Washington, DC, USA, 2007; p. 384. [Google Scholar] [CrossRef]
- Van Ravenswaay, R.O.; Henry, P.; Ammerman, C. Effects of time and dietary iron on tissue iron concentration as an estimate of relative bioavailability of supplemental iron sources for ruminants. Anim. Feed Sci. Technol. 2001, 90, 185–198. [Google Scholar] [CrossRef]
- Henry, P.R.; Ammerman, C.B.; Littell, R.C. Relative bioavailability of manganese from a manganese-methionine complex and inorganic sources for ruminants. J. Dairy Sci. 1992, 75, 3473–3478. [Google Scholar] [CrossRef]
- Wong-Valle, J.; Henry, P.R.; Ammerman, C.B.; Rao, P.V. Estimation of the relative bioavailability of manganese sources for sheep. J. Anim. Sci. 1989, 67, 2409–2414. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.K.; Hidroglou, M. Effects of dietary chelated and sequestered zinc and zinc sulfate on growing lambs fed a purified diet. Can. J. Anim. Sci. 1977, 57, 93–99. [Google Scholar] [CrossRef]
- Spears, J.W.; Samsell, L.J. Relative availability of zinc in zinc methionine and zinc oxide for ruminants. J. Anim. Sci. 1986, 63 (Suppl. 1), 402. [Google Scholar]
- Hassan, A. Effect of Supplementation of Chelated Zinc on Milk Production in Ewes. Food Nutr. Sci. 2011, 2, 706–713. [Google Scholar] [CrossRef] [Green Version]
- Rojas, L.X.; McDowell, L.R.; Cousins, R.J.; Martin, F.G.; Wilkinson, N.S.; Johnson, A.B.; Velasquez, J.B. Relative bioavailability of two organic and two inorganic zinc sources fed to sheep. J. Anim. Sci. 1995, 73, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Grešáková, Ľ.; Tokarčíková, K.; Čobanová, K. Bioavailability of Dietary Zinc Sources and Their Effect on Mineral and Antioxidant Status in Lambs. Agriculture 2021, 11, 1093. [Google Scholar] [CrossRef]
- Zhu, Z.; Yan, L.; Hu, S.; An, S.; Lv, Z.; Wang, Z.; Wu, Y.; Zhu, Y.; Zhao, M.; Gu, C.; et al. Effects of the different levels of dietary trace elements from organic or inorganic sources on growth performance, carcass traits, meat quality, and faecal mineral excretion of broilers. Arch. Anim. Nutr. 2019, 73, 324–337. [Google Scholar] [CrossRef]
- Vieira, R.; Ferket, P.; Malheiros, R.; Hannas, M.; Crivellari, R.; Moraes, V.; Elliott, S. Feeding low dietary levels of organic trace minerals improves broiler performance and reduces excretion of minerals in litter. Br. Poult. Sci. 2020, 61, 574–582. [Google Scholar] [CrossRef]
- Qiu, J.L.; Zhou, Q.; Zhu, J.M.; Lu, X.T.; Liu, B.; Yu, D.Y.; Lin, G.; Ao, T.; Xu, J.M. Organic trace minerals improve eggshell quality by improving the eggshell ultrastructure of laying hens during the late laying period. Poult. Sci. 2020, 99, 1483–1490. [Google Scholar] [CrossRef]
- Stefanello, C.; Santos, T.C.; Murakami, A.E.; Martins, E.N.; Carneiro, T.C. Productive performance, eggshell quality, and eggshell ultrastructure of laying hens fed diets supplemented with organic trace minerals. Poult. Sci. 2014, 93, 104–113. [Google Scholar] [CrossRef]
- Świątkiewicz, S.; Arczewska-Wlosek, A.; Józefiak, D. The efficacy of organic minerals in poultry nutrition: Review and implications of recent studies. World’s Poult. Sci. J. 2014, 70, 475–486. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Vieira, B.S.; Applegate, T.J. Influence of dietary zinc, copper, and manganese on the intestinal health of broilers under Eimeria challenge. Front. Vet. Sci. 2020, 7, 32064270. [Google Scholar] [CrossRef] [Green Version]
- Das, T.; Mondal, M.; Biswas, P.; Bairagi, B.; Samanta, C.C. Influence of Level of Dietary Inorganic and Organic Copper and Energy Level on the Performance and Nutrient Utilization of Broiler Chickens. Asian-Australas. J. Anim. Sci. 2010, 23, 82–89. [Google Scholar] [CrossRef]
- Jegede, A.V.; Oduguwa, O.O.; Bamgbose, A.M.; Fanimo, A.O.; Nollet, L. Growth response, blood characteristics and copper accumulation in organs of broilers fed on diets supplemented with organic and inorganic dietary copper sources. Br. Poult. Sci. 2011, 52, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Kwiecień, M.; Winiarska-Mieczan, A.; Zawiślak, K.; Sroka, S. Effect of copper glycinate chelate on biomechanical, morphometric and chemical properties of chicken femur. Ann. Anim. Sci. 2014, 14, 127–139. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.B.; Seo, Y.M.; Shin, K.S.; Rhee, A.R.; Han, J.; Paik, I.K. Effects of supplemental copper-methionine chelate and copper-soy proteinate on the performance, blood parameters, liver mineral content, and intestinal microflora of broiler chickens. J. Appl. Poult. Res. 2011, 20, 21–32. [Google Scholar] [CrossRef]
- Shamsudeen, P.; Shrivastava, H.O.P. Ramsingh. Biointeraction of chelated and inorganic copper with aflatoxin on growth performance of broiler chicken. Int. J. Vet. Sci. 2013, 2, 106–110. [Google Scholar]
- Ao, T.; Pierce, J. The replacement of inorganic mineral salts with mineral proteinates in poultry diets. World’s Poult. Sci. J. 2013, 69, 5–16. [Google Scholar] [CrossRef]
- Norvell, M.J.; Thomas, M.C.; Goatcher, W.D.; Gable, D.A.; Calvert, C.C. Some effects of high dietary levels of various salts of copper in broiler chickens. In Proceedings of the University of Missouri’s Annual Conference on Trace Substances in Environmental Health, Columbia, MO, USA, 11 June 1974; pp. 367–372. [Google Scholar]
- Aoyagi, S.; Baker, D.H. Bioavailability of copper in analytical-grade and feed-grade inorganic copper sources when fed to provide copper at levels below the chick’s requirement. Poult. Sci. 1993, 72, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Lu, L.; Li, S.; Xie, J.; Zhang, L.; Wang, R.; Luo, X. Copper in organic proteinate or inorganic sulfate form is equally bioavailable for broiler chicks fed a conventional corn-soybean meal diet. Biol. Trace Elem. Res. 2012, 147, 142–148. [Google Scholar] [CrossRef]
- Elvehjem, C.A.; Hart, E.B. The relation of iron and copper to hemoglobin syntheisi in the chick. J. Biol. Chem. 1929, 84, 131–141. [Google Scholar] [CrossRef]
- Elvehjem, C.A.; Hart, E.B.; Sherman, W.C. The availability of iron from different sources for hemoglobin formation. J. Biol. Chem. 1933, 103, 63–70. [Google Scholar] [CrossRef]
- Cao, J.; Luo, X.G.; Henry, P.R.; Ammerman, C.B.; Littell, R.C.; Miles, R.D. Effect of dietary iron concentration, age, and length of iron feeding on feed intake and tissue iron concentration of broiler chicks for use as a bioassay of supplemental iron sources. Poult. Sci. 1996, 75, 495–504. [Google Scholar] [CrossRef]
- Ma, X.Y.; Liu, S.B.; Lu, L.; Li, S.F.; Xie, J.J.; Zhang, L.Y.; Zhang, J.H.; Luo, X.G. Relative bioavailability of iron proteinate for broilers fed a casein-dextrose diet. Poult. Sci. 2014, 93, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Y.; Lu, L.; Luo, X.G. The chemical characteristics of organic iron sources and their relative bioavailabilities for broilers fed a conventional corn-soybean meal diet. J. Anim. Sci. 2016, 94, 2378–2396. [Google Scholar] [CrossRef] [Green Version]
- Schaible, P.; Bandemer, S.L.; Davidson, J. The Manganese Content of Feedstufls and Its Relation to Poultry Nutrition; Michigan State University: East Lansing, MI, USA, 1938; p. 32. [Google Scholar]
- Southern, L.L.; Baker, D.H. Excess manganese ingestion in the chick. Poult. Sci. 1983, 62, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Gallup, W.D.; Norris, L.C. The Amount of Manganese Required to Prevent Perosis in the Chick. Poult. Sci. 1939, 18, 76–82. [Google Scholar] [CrossRef]
- Black, J.R.; Ammerman, C.B.; Henry, P.R.; Miles, R.D. Biological availability of manganese sources and effects of high dietary manganese on tissue mineral composition of broiler-type chicks. Poult. Sci. 1984, 63, 1999–2006. [Google Scholar] [CrossRef] [PubMed]
- Wong-Valle, J.; Ammerman, C.B.; Henry, P.R.; Rao, P.V.; Miles, R.D. Bioavailability of manganese from feed grade manganese oxides for broiler chicks. Poult. Sci. 1989, 68, 1368–1373. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Waldroup, P. Evaluation of Mintrex® manganese as a source of manganese for young broilers. Int. J. Poult. Sci 2006, 5, 708–713. [Google Scholar]
- Watson, L.T.; Ammerman, C.B.; Miller, S.M.; Harms, R.H. Biological availability to chicks of manganese from different inorganic sources. Poult. Sci. 1971, 50, 1693–1700. [Google Scholar] [CrossRef]
- Henry, P.R.; Ammerman, C.B.; Miles, R.D. Bioavailability of manganese sulfate and manganese monoxide in chicks as measured by tissue uptake of manganese from conventional dietary levels. Poult. Sci. 1986, 65, 983–986. [Google Scholar] [CrossRef]
- Luo, X. Studies on the Optimal Manganese Level and Its Bioavailability in a Practical Diet for Broiler Chicks; Chinese Academy of Agricultural Science: Beijing, China, 1989. [Google Scholar]
- Smith, M.O.; Sherman, I.L.; Miller, L.C.; Robbins, K.R.; Halley, J.T. Relative biological availability of manganese from manganese proteinate, manganese sulfate, and manganese monoxide in broilers reared at elevated temperatures. Poult. Sci. 1995, 74, 702–707. [Google Scholar] [CrossRef]
- Korol, W.; Wójcik, S.; Matyka, S.; Hansen, T.S. Availability of manganese from different manganese oxides and their effect on performance of broiler chickens. J. Anim. Feed Sci. 1996, 5, 273–279. [Google Scholar] [CrossRef]
- Watson, L.T.; Ammerman, C.B.; Miller, S.M.; Harms, R.H. Biological Assay of Inorganic Manganese for Chicks. Poult. Sci. 1970, 49, 1548–1554. [Google Scholar] [CrossRef]
- Henry, P.; Ammerman, C.; Miles, R. Bioavailability of manganese monoxide and manganese dioxide for broiler chicks. Nutr. Rep. Int. 1987, 36, 425–433. [Google Scholar]
- Scheideler, S.E. Interaction of dietary calcium, manganese, and manganese source (Mn oxide or Mn methionine complex) on chick performance and manganese utilization. Biol. Trace. Elem. Res. 1991, 29, 217–228. [Google Scholar] [CrossRef]
- Fly, A.D.; Izquierdo, O.A.; Lowry, K.R.; Baker, D.H. Manganese bioavailability in a Mn-methionine chelate. Nutr. Res. 1989, 9, 901–910. [Google Scholar] [CrossRef]
- Henry, P.R.; Ammerman, C.B.; Miles, R.D. Relative bioavailability of manganese in a manganese-methionine complex for broiler chicks. Poult. Sci. 1989, 68, 107–112. [Google Scholar] [CrossRef]
- Baker, D.H.; Halpin, K.M. Efficacy of a manganese-protein chelate compared with that of manganese sulfate for chicks. Poult. Sci. 1987, 66, 1561–1563. [Google Scholar] [CrossRef]
- Ao, T.; Pierce, J.L.; Pescatore, A.J.; Ford, M.J.; Cantor, A.H.; Dawson, K.A.; Paul, M. Evaluation of organic Mn (Bioplex Mn®) as a Mn source for chicks. Poult. Sci. 2008, 87, 172–173. [Google Scholar]
- Saldanha, M.M.; Araújo, I.C.S.; Triguineli, M.V.; Vaz, D.P.; Ferreira, F.N.A.; Albergaria, J.D.S.; Fontes, D.O.; Lara, L.J.C. Relative bioavailability of manganese in relation to proteinate and sulfate sources for broiler chickens from one to 20 d of age. Poult. Sci. 2020, 99, 5647–5652. [Google Scholar] [CrossRef]
- Wang, F.; Lu, L.; Li, S.; Liu, S.; Zhang, L.; Yao, J.; Luo, X. Relative bioavailability of manganese proteinate for broilers fed a conventional corn-soybean meal diet. Biol. Trace Elem. Res. 2012, 146, 181–186. [Google Scholar] [CrossRef]
- Liao, X.D.; Wang, G.; Lu, L.; Zhang, L.Y.; Lan, Y.X.; Li, S.F.; Luo, X.G. Effect of manganese source on manganese absorption and expression of related transporters in the small intestine of broilers. Poult. Sci. 2019, 98, 4994–5004. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Luo, X.G.; Lu, L.; Liu, B.; Yu, S.X. Effect of Manganese Source on Manganese Absorption by the Intestine of Broilers1 1Supported by the National Basic Research Program of China (project no. 2004CB117501), National Natural Science Foundation of China (project no. 30270968), and Chinese Academy of Agricultural Sciences Foundation for First-Place Outstanding Scientists. Poult. Sci. 2006, 85, 1947–1952. [Google Scholar] [CrossRef]
- Ji, F.; Luo, X.G.; Lu, L.; Liu, B.; Yu, S.X. Effects of manganese source and calcium on manganese uptake by in vitro everted gut sacs of broilers’ intestinal segments. Poult. Sci. 2006, 85, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.-P.; Lu, L.; Wang, R.-L.; Xi, L.; Zhang, L.-Y.; Luo, X.-G. Manganese source affects manganese transport and gene expression of divalent metal transporter 1 in the small intestine of broilers. Br. J. Nutr. 2012, 108, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Yi, J.; Chen, Y.; Huang, M.; Zhu, N. Relative Bioavailability of Broiler Chickens Fed with Zinc Hydroxychloride and Sulfate Sources for Corn-Soybean Meal. Biol. Trace Elem. Res. 2021, 200, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, S.F.; Luo, X.G.; Lu, L.; Crenshaw, T.D.; Bu, Y.; Liu, B.; Kuang, X.; Shao, G.Z.; Yu, S.X. Bioavailability of organic manganese sources in broilers fed high dietary calcium. Anim. Feed Sci. Technol. 2005, 123, 703–715. [Google Scholar] [CrossRef]
- Liu, S.; Li, S.; Lu, L.; Xie, J.; Zhang, L.; Wang, R.; Luo, X. The effectiveness of zinc proteinate for chicks fed a conventional corn-soybean meal diet. J. Appl. Poult. Res. 2013, 22, 396–403. [Google Scholar] [CrossRef]
- Luo, X.; Li, S.; Liu, B.; Bu, Y.; Kuang, X.; Yu, S. Bioavailabilities of manganese sources based on heart manganese-containing superoxide dismutase gene expression for broilers. Wei Sheng Yan Jiu J. Hyg. Res. 2004, 33, 681–686. [Google Scholar]
- Miles, R.; Henry, P.; Sampath, V.; Shivazad, M.; Comer, C. Relative Bioavailability of Novel Amino Acid Chelates of Manganese and Copper for Chicks. J. Appl. Poult. Res. 2003, 12, 417–423. [Google Scholar] [CrossRef]
- Ashmead, D. The Role of Metal Amino Acid Chelate; Academic Press: San Diego, CA, USA, 1993. [Google Scholar]
- Huang, Y.; Lu, L.; Xie, J.; Li, S.; Li, X.; Liu, S.; Zhang, L.; Xi, L.; Luo, X. Relative bioavailabilities of organic zinc sources with different chelation strengths for broilers fed diets with low or high phytate content. Anim. Feed Sci. Technol. 2013, 179, 144–148. [Google Scholar] [CrossRef]
- Roberson, R.H.; Schaible, P.J. The Availability to the Chick of Zinc as the Sulfate, Oxide or Carbonate. Poult. Sci. 1960, 39, 835–837. [Google Scholar] [CrossRef]
- Edwards, H.M.; Baker, D.H. Zinc bioavailability in soybean meal. J. Anim. Sci. 2000, 78, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Ao, T.; Pierce, J.L.; Pescatore, A.J.; Cantor, A.C.; Dawson, K.A.; Ford, M.J. Effects of feeding reduced levels of organic minerals (Bioplex) on the development of white layer pullets. Poult. Sci. 2009, 88, 197. [Google Scholar]
- Wedekind, K.J. The bioavailability of zinc-methionine relative to zinc sulfate is affected by calcium level. Poult. Sci. 1994, 73, 114. [Google Scholar]
- Cao, J.; Henry, P.R.; Davis, S.R.; Cousins, R.J.; Miles, R.D.; Littell, R.C.; Ammerman, C.B. Relative bioavailability of organic zinc sources based on tissue zinc and metallothionein in chicks fed conventional dietary zinc concentrations. Anim. Feed Sci. Technol. 2002, 101, 161–170. [Google Scholar] [CrossRef]
- Ao, T.; Pierce, J.L.; Power, R.F.G.; Dawson, K.A.; Pescatore, A.J.; Cantor, A.H.; Ford, M.J. Evaluation of Bioplex Zn® as an organic zinc source for chicks. Int. J. Poult. Sci. 2006, 5, 808–811. [Google Scholar]
- Sahraei, M.; Janmmohamadi, H.; Taghizadeh, A.; Ali Moghadam, G.; Abbas Rafat, S. Estimation of the Relative Bioavailability of Several Zinc Sources for Broilers Fed a Conventional Corn-Soybean Meal Diet. J. Poult. Sci. 2013, 50, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Apgar, G.A.; Kornegay, E.T.; Lindemann, M.D.; Notter, D.R. Evaluation of copper sulfate and a copper lysine complex as growth promoters for weanling swine. J. Anim. Sci. 1995, 73, 2640–2646. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, S.; Bailoni, L.; Ramanzin, M.; Vincenzi, R.; Simonetto, A.; Bittante, G. Effect of proteinate or sulphate mineral sources on trace elements in blood and liver of piglets. Anim. Sci. 2000, 71, 131–139. [Google Scholar] [CrossRef]
- Creech, B.L.; Spears, J.W.; Flowers, W.L.; Hill, G.M.; Lloyd, K.E.; Armstrong, T.A.; Engle, T.E. Effect of dietary trace mineral concentration and source (inorganic vs. chelated) on performance, mineral status, and fecal mineral excretion in pigs from weaning through finishing. J. Anim. Sci. 2004, 82, 2140–2147. [Google Scholar] [CrossRef] [Green Version]
- Peters, J.C.; Mahan, D.C. Effects of dietary organic and inorganic trace mineral levels on sow reproductive performances and daily mineral intakes over six parities. J. Anim. Sci. 2008, 86, 2247–2260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veum, T.; Shannon, M.; Wu, C.; Bollinger, D.; Ellersieck, M. Copper proteinate in weanling pig diets for enhancing growth performance and reducing fecal copper excretion compared with copper sulfate. J. Anim. Sci. 2004, 82, 1062–1070. [Google Scholar] [CrossRef] [PubMed]
- Burkett, J.; Stalder, K.; Powers, W.; Bregendahl, K.; Pierce, J.; Baas, T.; Bailey, T.; Shafer, B. Effect of inorganic and organic trace mineral supplementation on the performance, carcass characteristics, and fecal mineral excretion of phase-fed, grow-finish swine. Asian-Australas. J. Anim. Sci. 2009, 22, 1279–1287. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Choi, S.C.; Chae, B.J.; Lee, J.K.; Acda, S.P. Evaluation of Metal-Amino Acid Chelates and Complexes at Various Levels of Copper and Zinc in Weanling Pigs and Broiler Chicks. Asian-Australas. J. Anim. Sci. 2001, 14, 1734–1740. [Google Scholar] [CrossRef]
- Ma, L.; Hou, C.; He, J.; Qiu, J.; Lu, X.; Guo, Y.; Liu, B.; Lin, G.; Xue, Y.; Dongyou, Y. Effect of compound organic trace minerals on growth performance, serum indexes and micromineral excretion in fattening pigs. J. Zhejiang Univ. (Agric. Life Sci.) 2018, 44, 181–189. [Google Scholar] [CrossRef]
- Pierce, J.; Shafer, B.; Stalder, K.; Burkett, J. Nutritional means to lower trace mineral excretion from swine and poultry without compromising performance. Poult. Sci. 2005, 84, 1–6. [Google Scholar]
- Taylor-Pickard, J.A.; Nollet, L.; Geers, R. Performance, carcass characteristics and economic benefits of total replacement of inorganic minerals by organic forms in growing pig diets. J. Appl. Anim. Nutr. 2013, 2, e3. [Google Scholar] [CrossRef]
- Zhang, W.F.; Tian, M.; Song, J.S.; Chen, F.; Lin, G.; Zhang, S.H.; Guan, W.T. Effect of replacing inorganic trace minerals at lower organic levels on growth performance, blood parameters, antioxidant status, immune indexes, and fecal mineral excretion in weaned piglets. Trop. Anim. Health Prod. 2021, 53, 121. [Google Scholar] [CrossRef]
- Ma, L.; He, J.; Lu, X.; Qiu, J.; Hou, C.; Liu, B.; Lin, G.; Yu, D. Effects of low-dose organic trace minerals on performance, mineral status, and fecal mineral excretion of sows. Asian-Australas. J. Anim. Sci. 2019, 33, 132–138. [Google Scholar] [CrossRef]
- Shannon, M.; Boren, C.A.; Wu, C.; Huntington, C.; Bollinger, D.; Veum, T. Evaluation of various inclusion rates of organic zinc either as polysaccharide or proteinate complex on the growth performance, plasma, and excretion of nursery pigs. J. Anim. Sci. 2004, 82, 1359–1366. [Google Scholar] [CrossRef]
- Veum, T.L.; Bollinger, D.W.; Ellersieck, M.R. Proteinated trace mineral and condensed fish protein digest in weanling pig diets. J. Anim. Sci. 1995, 73, 308. [Google Scholar]
- Lin, G.; Guo, Y.; Liu, B.; Wang, R.; Su, X.; Yu, D.; He, P. Optimal dietary copper requirements and relative bioavailability for weanling pigs fed either copper proteinate or tribasic copper chloride. J. Anim. Sci. Biotechnol. 2020, 11, 54. [Google Scholar] [CrossRef] [PubMed]
- Lebel, A.; Matte, J.J.; Guay, F. Effect of mineral source and mannan oligosaccharide supplements on zinc and copper digestibility in growing pigs. Arch. Anim. Nutr. 2014, 68, 370–384. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, Y.L.; Zhao, J.M.; Vazquez-Añón, M.; Stein, H.H. Digestibility and retention of zinc, copper, manganese, iron, calcium, and phosphorus in pigs fed diets containing inorganic or organic minerals. J. Anim. Sci. 2014, 92, 3407–3415. [Google Scholar] [CrossRef]
- Bertechini, A.; Fassani, É.; Brito, J.; Barrios, P. Effects of dietary mineral bioplex in pregnant and lactating sow diets on piglet performance and physiological characteristics. Rev. Bras. Zootec. 2012, 41, 624–629. [Google Scholar] [CrossRef] [Green Version]
- Baker, D.H.; Stein, H.H. Bioavailability of minerals and vitamins in feedstuffs. In Sustainable Swine Nutrition, 1st ed.; Chiba, L.I., Ed.; John Wiled & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 340–364. [Google Scholar]
- Chiba, L.I. Sustainable Swine Nutrition; Wiley: Hoboken, NJ, USA, 2012. [Google Scholar]
- Pickett, R.A.; Plumlee, M.P.; Beeson, W.M. Availability of dietary iron in diffent compounds for young pigs. J. Anim. Sci. 1961, 20, 946. [Google Scholar]
- Spears, J.W.; Schoenherr, W.D.; Kegley, E.B.; Flowers, W.L.; Alhusen, H.D. Efficacy of iron methionine as a source of iron for nursing pigs. J. Anim. Sci. 1992, 70, 243. [Google Scholar]
- Lewis, A.; Miller, P.S.; Wolverton, C. Bioavailability of Iron in Two Different Sources for Weanling Pigs; Animal Science Department, University of Nebraska: Lincoln, NE, USA, 1996. [Google Scholar]
- Brady, P.S.; Ku, P.K.; Ullrey, D.E.; Miller, E.R. Evaluation of an amino acid-iron chelate hematinic for the baby pig. J. Anim. Sci. 1978, 47, 1135–1140. [Google Scholar] [CrossRef] [Green Version]
- Ashmead, H.D.W. Amino Acid Chelation in Human and Animal Nutrition; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Egeli, A.; Framstad, T.; Morberg, H. Clinical Biochemistry, Haematology and Body Weight in Piglets. Acta Vet. Scand. 1998, 39, 381–393. [Google Scholar] [CrossRef]
- Kayongo-Male, H.; Ullrey, D.E.; Miller, E.R. Manganese (Mn) nutrition of the pig. 2 The availability of Mn from different sources to the growing pig. Bull. Anim. Health Prod. Afr. Bull. Sante Prod. Anim. Afr. 1980, 28, 145–153. [Google Scholar]
- Lisgara, Μ.; Skampardonis, V.; Leontides, L. Effect of diet supplementation with chelated zinc, copper and manganese on hoof lesions of loose housed sows. Porc. Health Manag. 2016, 2, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, M.C.; Hill, G.M. Trace Mineral Supplementation for the Intestinal Health of Young Monogastric Animals. Front. Vet. Sci. 2019, 6, 73. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.R.; Ku, P.K.; Hitchcock, J.P.; Magee, W.T. Availability of Zinc from Metallic Zinc Dust for Young Swine. J. Anim. Sci. 1981, 52, 312–315. [Google Scholar] [CrossRef]
- Hahn, J.D.; Baker, D.H. Growth and plasma zinc responses of young pigs fed pharmacologic levels of zinc. J. Anim. Sci. 1993, 71, 3020–3024. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Kornegay, E.T.; Schell, T. Influence of dietary lysine on the utilization of zinc from zinc sulfate and a zinc-lysine complex by young pigs. J. Anim. Sci. 1998, 76, 1064–1074. [Google Scholar] [CrossRef] [Green Version]
- Swinkels, J.W.G.M.; Kornegay, E.T.; Zhou, W.; Lindemann, M.D.; Webb, K.E., Jr.; Verstegen, M.W.A. Effectiveness of a zinc amino acid chelate and zinc sulfate in restoring serum and soft Tissue zinc concentrations when fed to zinc-depleted pigs. J. Anim. Sci. 1996, 74, 2420–2430. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Guo, Y. Beneficial effects of tetrabasic zinc chloride for weanling piglets and the bioavailability of zinc in tetrabasic form relative to ZnO. Anim. Feed Sci. Technol. 2007, 135, 75–85. [Google Scholar] [CrossRef]
- Martell, A.E.; Smith, R.M. Critical Stability Constants: Second Supplement; Springer: New York, NY, USA, 2013. [Google Scholar]
- Martell, A.E.; Hancock, R.D. Stability Constants and Their Measurement. In Metal Complexes in Aqueous Solutions. Modern Inorganic Chemistry; Springer: Boston, MA, USA, 1996. [Google Scholar]
- Sun, X.; Sarteshnizi, R.A.; Boachie, R.T.; Okagu, O.D.; Abioye, R.O.; Pfeilsticker Neves, R.; Ohanenye, I.C.; Udenigwe, C.C. Peptide–Mineral Complexes: Understanding Their Chemical Interactions, Bioavailability, and Potential Application in Mitigating Micronutrient Deficiency. Foods 2020, 9, 1402. [Google Scholar] [CrossRef]
- Gonzalez, P.; Bossak-Ahmad, K.; Vileno, B.; Wezynfeld, N.E.; El Khoury, Y.; Hellwig, P.; Hureau, C.; Bal, W.; Faller, P. Triggering Cu-coordination change in Cu(ii)-Ala-His-His by external ligands. Chem. Commun. 2019, 55, 8110–8113. [Google Scholar] [CrossRef]
- Mena, S.; Mirats, A.; Caballero, A.B.; Guirado, G.; Barrios, L.A.; Teat, S.J.; Rodriguez-Santiago, L.; Sodupe, M.; Gamez, P. Drastic Effect of the Peptide Sequence on the Copper-Binding Properties of Tripeptides and the Electrochemical Behaviour of Their Copper(II) Complexes. Chem. Eur. J. 2018, 24, 5153–5162. [Google Scholar] [CrossRef] [Green Version]
- Wienk, K.J.H.; Marx, J.J.M.; Beynen, A.C. The concept of iron bioavailability and its assessment. Eur. J. Nutr. 1999, 38, 51–75. [Google Scholar] [CrossRef] [PubMed]
- Holwerda, R.A.; Albin, R.C.; Madsen, F.C. Chelation effectivness of zinc proteinates demonstrated. Feedstuffs 1995, 67, 12–23. [Google Scholar]
- Yu, Y.; Lu, L.; Wang, R.L.; Xi, L.; Luo, X.G.; Liu, B. Effects of zinc source and phytate on zinc absorption by in situ ligated intestinal loops of broilers. Poult. Sci. 2010, 89, 2157–2165. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Cao, S.; Ma, X.; Li, X.; Liao, X.; Zhang, L.; Zhang, M.; Zhang, R.; Hou, S.; Luo, X.; et al. Organic iron absorption and expression of related transporters in the small intestine of broilers. Poult. Sci. 2021, 100, 101182. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lu, L.; Hao, S.; Wang, Y.; Zhang, L.; Liu, S.; Liu, B.; Li, K.; Luo, X. Dietary manganese modulates expression of the manganese-containing superoxide dismutase gene in chickens. J. Nutr. 2011, 141, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.G.; Li, S.F.; Lu, L.; Liu, B.; Kuang, X.; Shao, G.Z.; Yu, S.X. Gene expression of manganese-containing superoxide dismutase as a biomarker of manganese bioavailability for manganese sources in broilers. Poult. Sci. 2007, 86, 888–894. [Google Scholar] [CrossRef]
- Monteiro, S.C.; Lofts, S.; Boxall, A.B.A. Pre-Assessment of Environmental Impact of Zinc and Copper Used in Animal Nutrition. EFSA J. 2010, 7, 74E. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Choct, M. Trace mineral nutrition for broiler chickens and prospects of application of organically complexed trace minerals: A review. Anim. Prod. Sci. 2009, 49, 269–282. [Google Scholar] [CrossRef]
- Leeson, S.; Caston, L. Using minimal supplements of trace minerals as a method of reducing trace mineral content of poultry manure. Anim. Feed Sci. Technol. 2008, 142, 339–347. [Google Scholar] [CrossRef]
- Uchida, K.; Mandebvu, P.; Ballard, C.; Sniffen, C.; Carter, M. Effect of feeding a combination of zinc, manganese and copper amino acid complexes, and cobalt glucoheptonate on performance of early lactation high producing dairy cows. Anim. Feed. Sci. Technol. 2001, 93, 193–203. [Google Scholar] [CrossRef]
- Ballantine, H.T.; Socha, M.T.; Tomlinson, D.A.D.J.; Johnson, A.B.; Fielding, A.S.; Shearer, J.K.; Van Amstel, S.R. Effects of feeding complexed zinc, manganese, copper, and cobalt to late gestation and lactating dairy cows on claw integrity, reproduction, and lactation performance. Prof. Anim. Sci. 2002, 18, 211–218. [Google Scholar] [CrossRef]
- Case, C.; Shannon, M. Effect of feeding organic and inorganic sources of additional zinc on growth performance and zinc balance in nursery pigs. J. Anim. Sci. 2002, 80, 1917–1924. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Lu, X.; Ma, L.; Hou, C.; He, J.; Liu, B.; Yu, D.; Lin, G.; Xu, J. Low-dose of organic trace minerals reduced fecal mineral excretion without compromising performance of laying hens. Asian-Australas. J. Anim. Sci. 2020, 33, 588–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ao, T.; Pierce, J.L.; Pescatore, A.J.; Cantor, A.H.; Dawson, K.A.; Ford, M.J.; Paul, M. Effects of feeding different concentration and forms of zinc on the performance and tissue mineral status of broiler chicks. Br. Poult. Sci. 2011, 52, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Boruta, A.; Swierczewska, E.; Glebocka, K.; Nollet, L. Trace organic minerals as a replacement of inorganic sources for layers: Effects on productivity and mineral excretion. In Proceedings of World Poultry Science Association, Proceedings of the 16th European Symposium on Poultry Nutrition, Strasbourg, France, 26–30 August 2007; pp. 491–494. [Google Scholar]
- Abdallah, A.G.; El-Husseiny, O.M.; Abdel-Latif, K.O. Influence of some dietary organic mineral supplementations on broiler performance. Int. J. Poult. Sci. 2009, 8, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Acda, S.; Chae, B. A review on the applications of organic trace minerals in pig nutrition. Pak. J. Nutr. 2002, 1, 25–30. [Google Scholar] [CrossRef]
- Holder, V.B.; Jennings, J.S.; Covey, T.L. Effect of total replacement of trace minerals with Bioplex® proteinated minerals on the health and performance of lightweight, high-risk feedlot cattle. J. Anim. Sci. 2016, 94, 120. [Google Scholar] [CrossRef]
- Pomport, P.H.; Warren, H.E.; Taylor-Pickard, J. Effect of total replacement of inorganic with organic sources of key trace minerals on performance and health of high producing dairy cows. J. Appl. Anim. Nutr. 2021, 9, 23–30. [Google Scholar] [CrossRef]
- Gelsinger, S.L.; Pino, F.; Jones, C.M.; Gehman, A.M.; Heinrichs, A.J. Effects of a dietary organic mineral program including mannan oligosaccharides for pregnant cattle and their calves on calf health and performance. Prof. Anim. Sci. 2016, 32, 205–213. [Google Scholar] [CrossRef]
- Alltech. A Nutritional Revolution: 7 Changes in Nutritional Dogma on Alternative Sources of Trace Minerals, Alltech: USA. 2018. Available online: https://www.alltech.com/blog/nutritional-revolution-7-changes-nutritional-dogma-alternative-sources-trace-minerals (accessed on 14 May 2022).
- Broom, L.J. Necrotic enteritis; current knowledge and diet-related mitigation. World’s Poult. Sci. J. 2017, 73, 281–292. [Google Scholar] [CrossRef]
- Wade, B.; Keyburn, A. The true cost of necrotic enteritis. World Poult. 2015, 31, 16–17. [Google Scholar]
- He, B.; King, W.; Graugnard, D.; Dawson, K.A.; Bortoluzzi, C.; Applegate, T. Zinc source influences the gene expression of zinc transporters in the jejunum and cecal tonsils of broilers challenged with Eimeria maxima and Clostridium perfringens. Poult. Sci. 2019, 98, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, V.; Nollet, L.; Kovac, G. Effect of Dietary Supplementation of Trace Elements on the Growth Performance and Their Distribution in the Breast and Thigh Muscles Depending on the Age of Broiler Chickens. Acta Vet. Brno 2010, 79, 203–209. [Google Scholar] [CrossRef]
- Aksu, T.; Özsoy, B.; Aksu, D.; Yoruk, M.; Gul, M. The effects of lower levels of organically complexed zinc, copper and manganese in broiler diets on performance, mineral concentration of tibia and mineral excretion. Kafkas Univ. Vet. Fak. Derg. 2011, 17, 141–146. [Google Scholar]
- Aksu, D.; Aksu, T.; Özsoy, B.; Baytok, E. The effects of replacing inorganic with a lower level of organically complexed minerals (Cu, Zn and Mn) in broiler diets on lipid peroxidation and antioxidant defense systems. Asian-Australas. J. Anim. Sci. 2010, 23, 1066–1072. [Google Scholar] [CrossRef]
- Aksu, T.; Aksu, M.; Yoruk, M.A.; Karaoglu, M. Effects of organically-complexed minerals on meat quality in chickens. Br. Poult. Sci. 2011, 52, 558–563. [Google Scholar] [CrossRef]
- Leeson, S. A new look at trace mineral nutrition of poultry: Can we reduce environmental burden of poultry manure? In Proceedings of The Nutritional Biotechnology in the Feed and Food Industries, Proceedings of the 19th Annual Symposium, Lexington, KY, USA, 11–14 May 2003; pp. 125–131. [Google Scholar]
- Yuan, J.; Xu, Z.; Huang, C.; Zhou, S.; Guo, Y. Effect of dietary Mintrex-Zn/Mn on performance, gene expression of Zn transfer proteins, activities of Zn/Mn related enzymes and fecal mineral excretion in broiler chickens. Anim. Feed Sci. Technol. 2011, 168, 72–79. [Google Scholar] [CrossRef]
- Delles, R.M.; Xiong, Y.L.; True, A.D.; Ao, T.; Dawson, K.A. Dietary antioxidant supplementation enhances lipid and protein oxidative stability of chicken broiler meat through promotion of antioxidant enzyme activity. Poult. Sci. 2014, 93, 1561–1570. [Google Scholar] [CrossRef]
- Delles, R.M.; Naylor, A.; Kocher, A.; Dawson, K.A.; Samuel, R.S. Diets with organic trace minerals (Bioplex®) and yeast protein (NuPro®) improved the water-holding capacity of pork loin meat. J. Anim. Sci. 2016, 94, 66. [Google Scholar] [CrossRef]





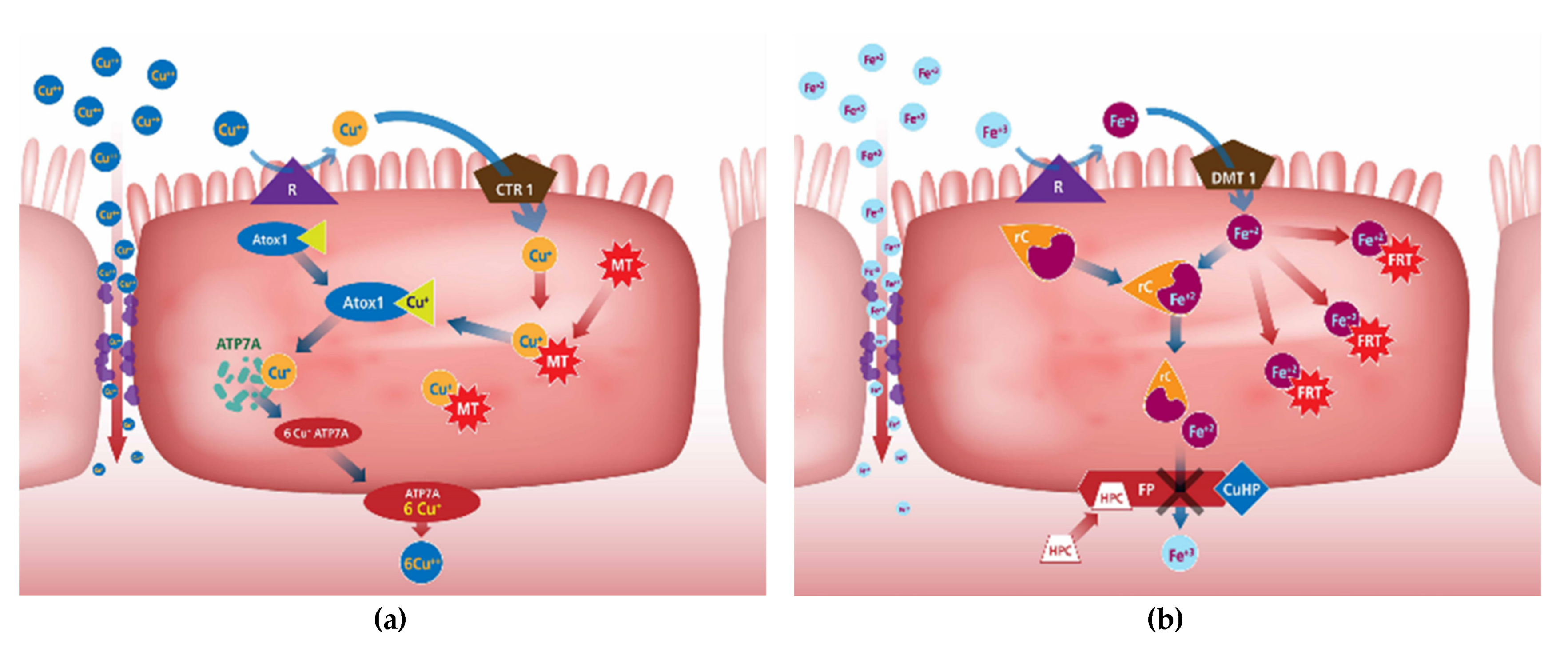{kind=link}
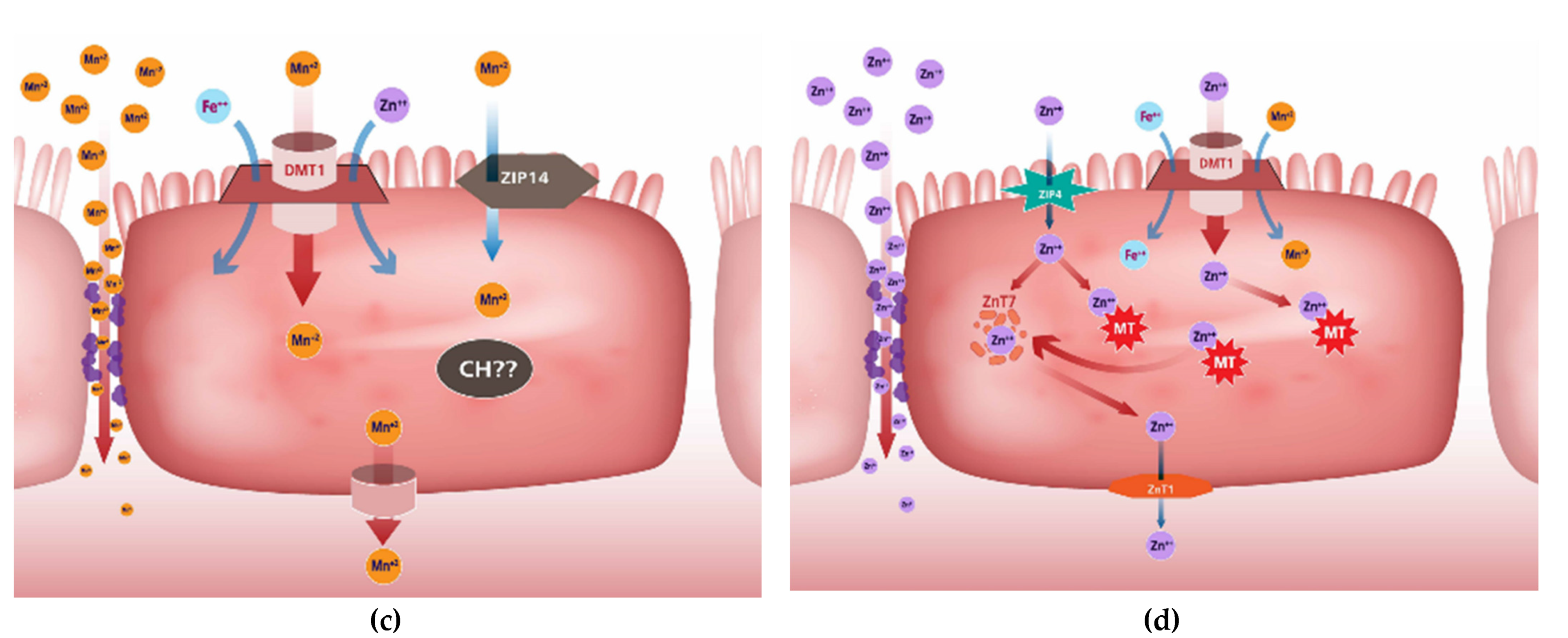{kind=link}
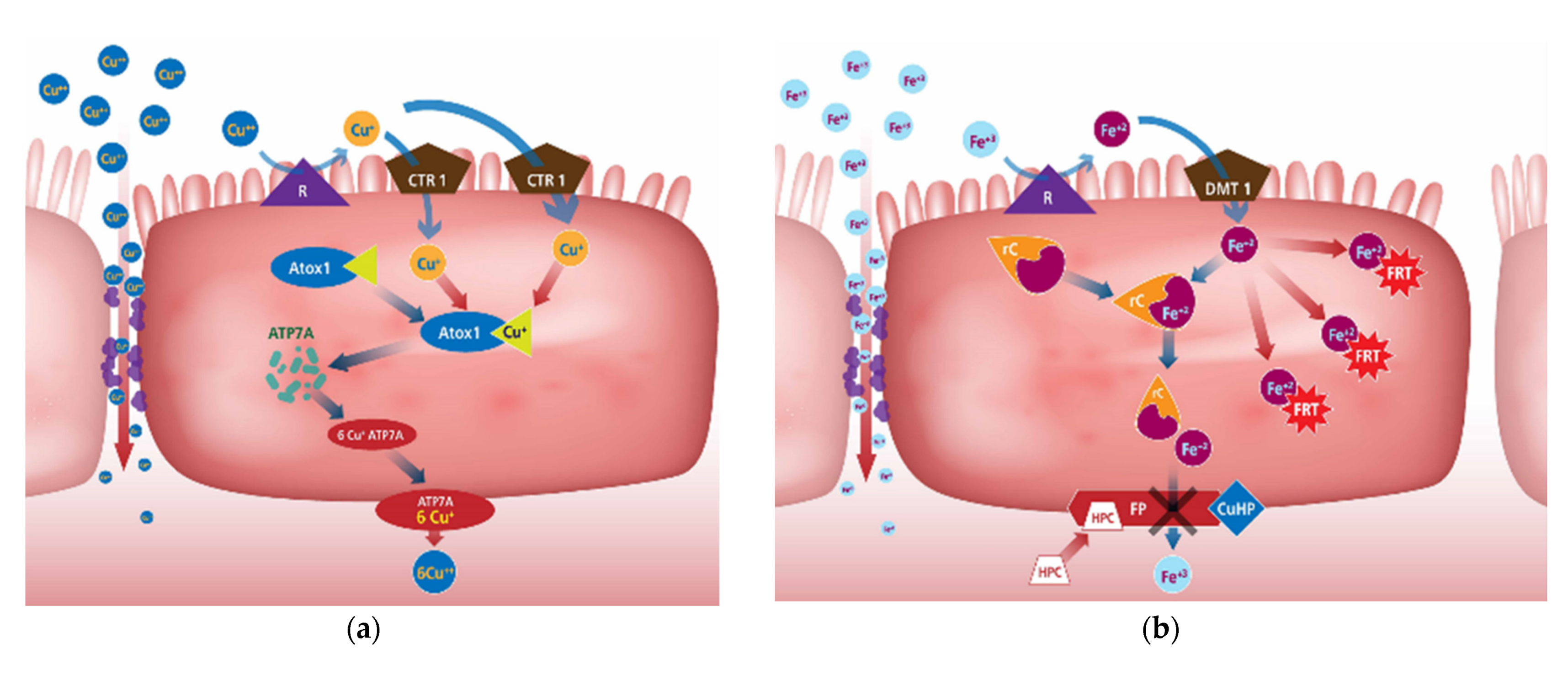{kind=link}
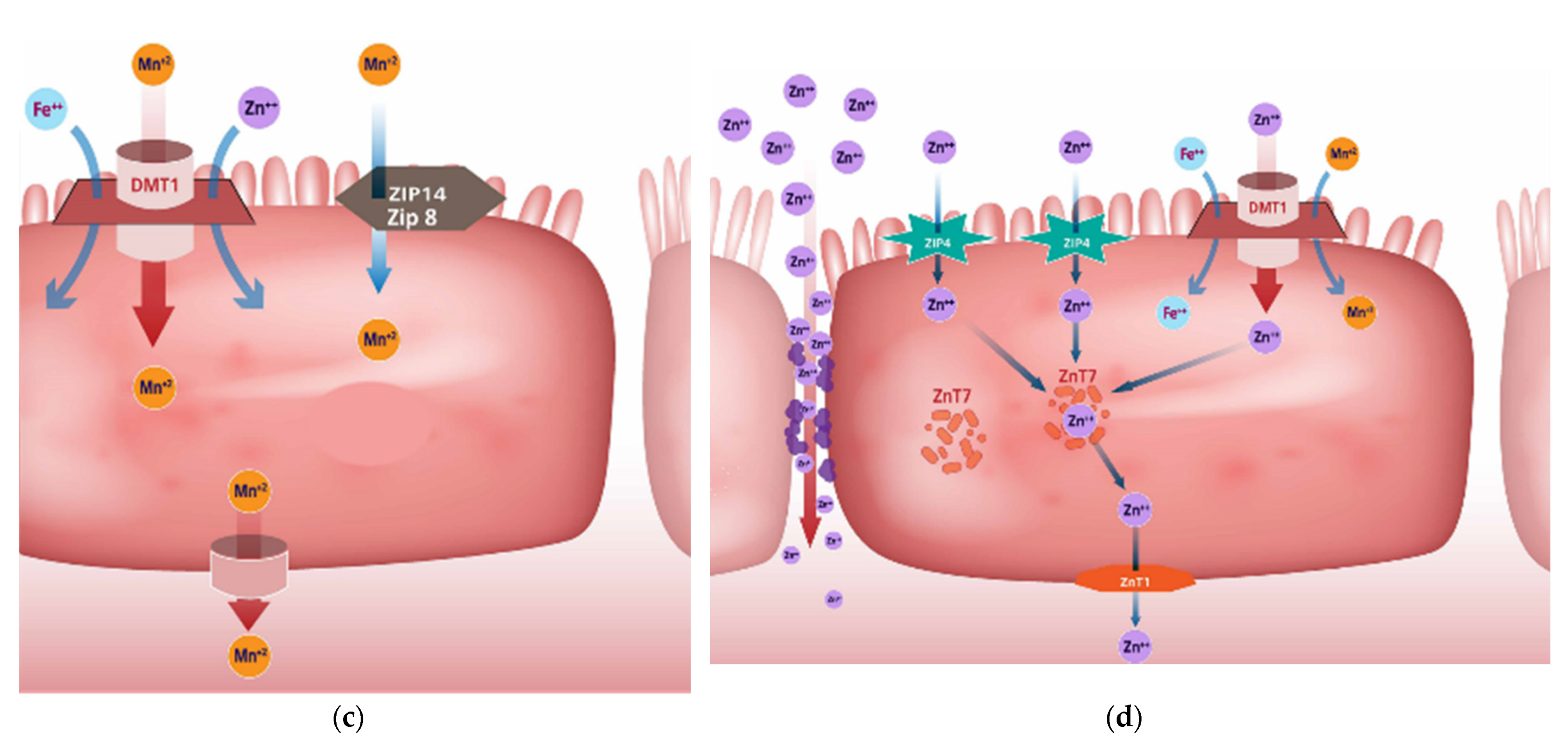{kind=link}
| Mineral | Function | Signs of Deficiency | EU Maximum Inclusion Levels: Maximum Content of Element in mg kg−1 of Complete Feed with a Moisture Content of 12% |
|---|---|---|---|
| Copper | Involved in metabolic reactions including cellular respiration, tissue pigmentation, haemoglobin formation (caeruloplasmin) and connective tissue development [9,10]. Essential component of several metalloenzymes [11,12]. Protects against oxidative stress [12,13]. | Muscle weakness, iron-deficient anaemia, hypopigmentation, bone changes resembling scurvy, defective connective tissue synthesis, hair abnormalities, impaired myelinisation of nerve tissues and neurological defects, altered lipid metabolism and cardiac malfunction [14,15,16]. | Bovines: Bovines before the start of rumination: 15 (total), Other bovines: 30 (total) Ovines: 15 (total) Caprines: 35 (total) Piglets: Suckling and weaned up to 4 weeks after weaning: 150 (total), from 5th week after weaning up to 8 weeks after weaning: 100 (total) Crustaceans: 50 (total) Other animals: 25 (total) [17] |
| Iron | Important for physiological function—haemoglobin, in which the heme portion functions to carry oxygen from the lungs to the tissues, mitochondrial Fe enzymes essential for oxidative production of cellular energy through Krebs cycle, transport of oxygen by myoglobin to cells and tissue of muscle. Important for immune function and lipid metabolism. | Supressed growth and blood volume [18]. Decreased animal performance, loss of appetite and weight, spasmodic breathing and ultimately death [19]. | Ovine: 500 (total (1)), Bovines and poultry: 450 (total (1)) Piglets up to 1 week before weaning: 250 mg/day (total (1)) Pet animals: 600 (total (1)) Other species: 750 (total (1)) [20] |
| Manganese | Constituent of multiple enzymes. Component of the organic matrix of bone and is essential for cartilage development. Involved in the metabolism of calcium and carbohydrates. Necessary for the utilisation of biotin, vitamin B1 and vitamin C [21]. Metabolic association between manganese and choline which affects fat metabolism in the liver [22]. | Impaired growth, skeletal abnormalities, abnormal reproduction function, ataxia in newborns, impaired carbohydrate and lipid metabolism and impaired mucopolysaccharide synthesis [23]. Poultry specific issues include: Perosis (slipped tendon), thin eggshell quality, chondrodystrophy in embryonic chicks, reduced egg production and hatchability | Fish: 100 (total) Other species: 150 (total) [24] |
| Zinc | Activates several enzymes. Component of many important metalloenzymes. Critically involved in cell replication and in the development of cartilage and bone [25]. Involved in protein synthesis, carbohydrate metabolism and many other biochemical reactions [26,27]. | Retarded growth, decreased feed intake, abnormal skeletal formation, alopecia, dermatitis, abnormal wool/hair/feather growth and impaired reproduction. Fetal abnormalities. Reduced egg hatchability [25]. Parakeratosis, diarrhoea and thymic atrophy [28]. | Dogs and cats: 200 (total) Salmonids and milk replacers for calves: 180 (total) Piglets, sows, rabbits and all fish other than salmonids: 150 (total) Other species and categories: 120 (total) [29]. |
| AAFCO | EU | ||
|---|---|---|---|
| Metal Proteinate (57.23) | The product resulting from the chelation of a soluble salt with amino acids and/or partially hydrolysed protein. It must be declared as an ingredient as the specific metal proteinate, e.g., copper proteinate, zinc proteinate etc. | Metal chelate of protein hydrolysates | A powder with a minimum content of x% metal where x = 10% copper, iron, manganese and zinc. Minimum of 50% copper, iron, manganese and 85% zinc chelated. Chemical formula: M(x)1–3. nH2O, M = metal, x = anion of protein hydrolysates containing any amino acid from soya protein hydrolysate. |
| Metal Polysaccharide Complex (57.29) | The product resulting from complexing of a soluble salt with a polysaccharide solution declared as an ingredient as the specific metal complex, e.g., copper polysaccharide complex, zinc polysaccharide complex etc. | ||
| Metal Amino Acid Chelate (57.142) | The product resulting from the reaction of a metal ion from a soluble metal salt with amino acids with a mole ratio of 1 mole of metal to 1 to 3 (preferably 2) moles of amino acids to form coordinate covalent bonds. The average weight of the hydrolysed amino acids must be approximately 150 and the resulting molecular weight of the chelate must not exceed 800. The minimum metal content must be declared. When used as a commercial feed ingredient, it must be declared as a specific metal amino acid chelate, e.g., copper amino acid chelate, zinc amino acid chelate etc. | Metal chelate of amino acids hydrate | Metal amino acid complex where the metal and the amino acids derived from soya protein are chelated via coordinate covalent bonds, as a powder with a minimum content of 10% copper and zinc, 9% iron and 8% manganese. Chemical formula: M(x)1–3. nH2O, M = metal, x = anion of any amino acid from soya protein hydrolysate. Maximum of 10% of the molecules exceeding 1500 Da. |
| Metal Amino Acid Complex (57:150) | The product resulting from complexing a soluble metal salt with an amino acid(s). Mineral metal content must be declared. When used as a commercial feed ingredient, it must be declared as a specific metal amino acid complex, e.g., copper amino acid complex, zinc amino acid complex etc. | ||
| Metal (specific amino acid) complex (57.151) | The product resulting from complexing a soluble metal salt with a specific amino acid. Minimum metal content must be declared. When used as a commercial feed ingredient, it must be declared as a specific metal, specific amino acid complex, e.g., copper lysine, zinc methionine etc. | Metal chelate of glycine hydrate (liquid)Metal chelate of glycine hydrate (solid) | A liquid with a minimum content of 6% copper or 7% zinc.Chemical formula: M(x)1–3. nH2O, M = Cu or Zn, x = anion of glycine A powder with a minimum content of 15% copper, iron, zinc and manganese and a maximum of 13% moisture for copper and 10% moisture for iron, zinc and manganese.Chemical formula: M(x)1–3. nH2O, M = metal, x = anion of glycine |
| Factor | Sub-Factor |
|---|---|
| Animal | Age Breed Health status Monogastric or ruminant Physiological state (e.g., growth, bone development, pregnancy, lactation, disease) Previous nutrition Production (performance) level and type of production Sex Species |
| Chemical aspects | Bond strength Chemical form and purity of the mineral sources Differences in dissociation rates of the mineral form from the ligand Particle size of the mineral Processing conditions/manufacturing method Solubility Stability |
| Dietary | Chemical composition of the diet (proximate analysis and mineral contents) Feedstuff composition of the diet and presence of dietary antagonists Level of supplementation of the minerals tested Overall diet digestibility Presence of antimicrobial growth promoters or (organic) acids Vitamin content |
| Environmental | Environmental stress Feeding method (dry or wet feeding; soaking) Housing and equipment Level of feeding expressed as energy level times maintenance requirement for energy Level of mineral intake Water supply level and quality |
| Evaluation | Reference/Standard source Model used for evaluation (dose-response; linear or non-liner) Choice of response criteria Direct or indirect measurement Duration of preliminary and test period Experimental design Levels of supplementation Number of replicates |
| Source | Cattle | Poultry | Sheep | Swine |
|---|---|---|---|---|
| Cupric sulphate | 100 | 100 | 100 | 100 |
| Copper acetate | 100 | |||
| Copper amino acid complex/chelate | 96–128 | 100 | ||
| Copper carbonate | 86 | 97 | ||
| Copper chelate of HMTBa | 111–112 | |||
| Copper chloride | 98 | 96 | ||
| Copper chloride, basic | 102–112 | |||
| Copper citrate | 101 | 74–99 | ||
| Copper EDTA | 91–104 | 96 | ||
| Copper glycine/glycinate | 131–157 2 | 96 | ||
| Copper lysine | 89–153 3 | 92–124 | 68–97 | 73–101 |
| Copper methionine | 88–117 | 150–152 | 100–107 | |
| Copper oxide | 81 | |||
| Copper proteinate | 82–147 | 79–111 | 103 | 114–263 6 |
| Cupric acetate | 93–188 2 | 93 5 | ||
| Cupric carbonate, basic | 113 | |||
| Cupric carbonate | 54–68 4 | 121 5 | 62–111 | |
| Cupric chloride | 102–121 | 106–110 | 102–123 | |
| Cupric chloride, tribasic (TBCC) | 87–196 2 | 70–134 | 97 | |
| Cupric oxide | 0–64 | 0–69 | 22–48 5 | 0–104 7 |
| Cupric sulphide | 25 | 11–35 | 0–69 | |
| Cuprous acetate | 100 | 98–110 | ||
| Cuprous chloride | 81–145 | |||
| Cuprous iodide | 46–82 | |||
| Cuprous oxide | 92–98 |
| Source | Cattle | Poultry | Sheep | Swine |
|---|---|---|---|---|
| Ferrous sulphate heptahydrate | 100 | 100 | 100 | 100 |
| Ferric ammonium citrate | 98–115 | 102 | ||
| Ferric chloride | 26–78 | |||
| Ferric choline citrate | 102 | 118–144 | ||
| Ferric citrate | 107 | 70–76 | 89–192 | |
| Ferric EDTA | 93 | |||
| Ferric glycerophosphate | 86–100 | |||
| Ferric orthophosphate | 4–36 | |||
| Ferric oxide | 0–82 | 12 | ||
| Ferric phytate | 47 | |||
| Ferric polyphosphate | 84–91 | |||
| Ferric pyrophosphate | 45 | |||
| Ferric sulphate | 37–104 | |||
| Ferrous ammonium sulphate | 99–100 | |||
| Ferrous carbonate–low 2 | 0–25 | 0–10 | 0–29 | 8–45 |
| Ferrous carbonate–high 2 | 79 | 55–88 | 13–112 | 55–101 |
| Ferrous chloride | 98–106 | |||
| Ferrous EDTA | 97–100 | 90–91 | ||
| Ferrous fumarate | 71–133 | |||
| Ferrous gluconate | 97 | |||
| Ferrous sulphate, anhydrous | 65–100 | |||
| Ferrous sulphate monohydrate | 91–103 | 87–101 | ||
| Ferrous tartrate | 70–83 | |||
| Iron methionine | 86–129 | 68–183 | ||
| Fe-ZnSO4.H2O | 112–126 | |||
| Iron proteinate | 96–174 | 123 | ||
| Iron, reduced | 8–66 | 27–86 | ||
| Sodium iron pyrophosphate | 2–30 | 29–81 | ||
| Zn-FeSO4.H2O | 93–96 |
| Source | Cattle | Poultry | Sheep | Swine |
|---|---|---|---|---|
| Manganese sulphate | 100 | 100 | 100 | |
| Manganese amino acid complex/chelate | 84–148 | |||
| Manganese carbonate | 32–101 | 20–93 | 95 | |
| Manganese chelate of HMTBa | 116–154 2 | |||
| Manganese dioxide | 29–106 | 25–67 | ||
| Manganese methionine | 95–174 2 | 93–164 | ||
| Manganese oxide | 46–103 | 31–91 | 96 | |
| Manganese propionate | 139 | |||
| Manganese proteinate | 86–163 | |||
| Manganous chloride | 93–102 |
| Source | Cattle | Poultry | Sheep | Swine |
|---|---|---|---|---|
| Fe-ZnSO4.H2O | 107 | |||
| Zinc acetate | ||||
| Zin amino acid complex/chelate | 76–164 | 102–110 | 102 | |
| Zinc chloride | 42 | 88–107 | ||
| Zinc chloride, basic | 108–119 | |||
| Zinc chloride, tetrabasic (TBZC) | 102–111 | 122–159 | ||
| Zinc sulphate (incl: basic & tribasic) | 100 | 76–124 | 83–99 | |
| Zinc aspartate | ||||
| Zinc carbonate | 58 | 78–123 | 105–106 | 98 |
| Zinc, chelated | 91–125 | |||
| Zinc citrate | 128 | |||
| Zinc EDTA | 110–118 | 17 | ||
| Zinc, elemental | 102 | |||
| Zinc glycine | 82–335 | |||
| Zinc lysine | 100 | 106–111 | 114 | 24–110 |
| Zinc methionine | 98–133 | 77–292 | 95–134 | 60–116 |
| Zinc methionine hydroxy analog (ZnHMTBa) | 161–441 | |||
| Zinc oxide | 98–101 | 22–108 | 74–106 | 50–110 |
| Zinc picolinate | 31–104 | |||
| Zinc polysaccharide complex | 144 | 94 | ||
| Zinc propionate | 116–119 | |||
| Zinc proteinate | 70–200 | 56–254 | ||
| Zinc, sequestered | 97–108 | |||
| Zn-FeSO4.H2O | 99 |
| Source | RV1, % | Qf | Standard | Response Criterion | MethodCalc. 2 | Type Diet | Added Level, mg kg−1 | Reference |
|---|---|---|---|---|---|---|---|---|
| Mn amino acid complex A (M) (6.48% Mn) | 114–273 | 45.3 | MnSO4.7H2O | Plasma Mn, Abs | N—21 mg kg−1 | 90 | Ji et al. [273] | |
| Mn amino acid complex B (S) (7.86% Mn) | 129–360 | 115.4 | MnSO4.7H2O | Abs, Plasma | N—21 mg kg−1 | 90 | Ji et al. [273] | |
| MnSO4 + Gly | 111–318 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [273] | ||
| MnSO4 + Met | 150–305 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [273] | ||
| Mn-Gly chelate | 139–333 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [273] | ||
| Mn-Met chelate | 170–373 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [273] | ||
| Mn-Met complex E (W) (8.27% Mn) | 110–160 | 3.2 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [273] | |
| Mn amino acid complex A (M) (6.48% Mn) | 98–182 | 45.3 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | |
| Mn amino acid complex B (S) (7.86% Mn) | 102–213 | 115.4 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | |
| MnSO4 + Gly | 52–90 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | ||
| MnSO4 + Met | 75–194 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | ||
| Mn-Gly chelate | 82–159 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | ||
| Mn-Met chelate | 161–230 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | ||
| Mn-Met complex E (W) (8.27% Mn) | 80–168 | 3.2 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | |
| Mn amino acid complex A (M) (6.48% Mn) d31, normal Ca | 133–164 | 45.3 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | |
| Mn amino acid complex A (M) (6.48% Mn) d31, high Ca | 100–117 | 45.3 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | |
| Mn amino acid complex B (S) (7.86% Mn) d31, normal Ca | 145–191 | 115.4 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | |
| Mn amino acid complex B (S) (7.86% Mn) d31, high Ca | 107–165 | 115.4 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | |
| Mn-Met complex E (W) (8.27% Mn) d31, normal Ca | 108–182 | 3.2 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | |
| Mn-Met complex E (W) (8.27% Mn) d31, high Ca | 106–143 | 3.2 | MnSO4.7H2O | Abs | N—21 mg kg−1 | 90 | Ji et al. [274] | |
| Mn AA complex (OW) | 103–113 | 2.35 | MnSO4.7H2O RG | Plasma Mn | N—14 mg kg−1 | 110 | Liao et al. [272] | |
| Mn AA chelate (OM) | 125–141 | 61.9 | MnSO4.H2O RG | Plasma Mn | N—14 mg kg−1 | 110 | Liao et al. [272] | |
| Mn AA proteinate (OS) | 136–169 | 147 | MnSO4.H2O RG | Plasma Mn | N—14 mg kg−1 | 110 | Liao et al. [272] | |
| Mn methionine E (W) (8.27% Mn) | 102–103 | 3.2 | MnSO4.H2O RG | Bone, Heart MnSOD mRNA, Bone | N—16 mg kg−1 | 120 | Luo et al. [334] | |
| Mn amino acid B (M) (6.48% Mn) | 98–110 | 45.3 | MnSO4.H2O RG | Bone, Heart MnSOD mRNA | N—16 mg kg−1 | 120 | Luo et al. [334] | |
| Mn amino acid C (S) (7.86% Mn) | 99–102 | 115.4 | MnSO4.H2O RG | Bone, Heart MnSOD mRNA | N—16 mg kg−1 | 120 | Luo et al. [334] | |
| Mn AA A (W) | 99–105 | 2.35 | MnSO4.H2O RG | Heart MnSOD mRNA, Heart Mn, MnSOD protein conc., MnSOD activity | N—16 mg kg−1 | 100–200 | Li et al. [333] | |
| Mn AA B (M) | 104–118 | 16.85 | MnSO4.H2O RG | Heart MnSOD mRNA, Heart Mn, MnSOD protein conc., MnSOD activity | N—16 mg kg−1 | 100–200 | Li et al. [333] | |
| Mn AA C (S) | 102–112 | 147 | MnSO4.H2O RG | Heart MnSOD mRNA, Heart Mn, MnSOD protein conc., MnSOD activity | N—16 mg kg−1 | 100–200 | Li et al. [333] | |
| Mn AA (M) (9.06% Mn) (d7, d14) | 133, 136 | 16.85 | MnSO4.H2O RG | Plasma Mn | N—13 mg kg−1 | 100 | Bai et al. [275] | |
| Mn AA (S) (10.18% Mn) (d7, d14) | 146, 175 | 147 | MnSO4.H2O RG | Plasma Mn | N—13 mg kg−1 | 100 | Bai et al. [275] | |
| Zinc amino acid complex C (W) (11.93% Zn) | 105–162 | 6.48 | ZnSO4.7H2O RG | Abs | N—90 mg kg−1 | 40 | Yu et al. [331] | |
| Zn-Gly chelate | 109–160 | ZnSO4.7H2O RG | Abs | N—90 mg kg−1 | 40 | Yu et al. [331] | ||
| Zn-Met chelate | 109–146 | ZnSO4.7H2O RG | Abs | N—90 mg kg−1 | 40 | Yu et al. [331] | ||
| Zinc proteinate A (S) (18.61% Zn) | 112–196 | 944.02 | ZnSO4.7H2O RG | Abs | N—90 mg kg−1 | 40 | Yu et al. [331] | |
| Zinc proteinate B (M) (13.27% Zn) | 108–189 | 30.73 | ZnSO4.7H2O RG | Abs | N—90 mg kg−1 | 40 | Yu et al. [331] | |
| ZnSO4.7H2O + Gly | 77–97 | ZnSO4.7H2O RG | Abs | N—90 mg kg−1 | 40 | Yu et al. [331] | ||
| ZnSO4.7H2O + Met | 88–99 | ZnSO4.7H2O RG | Abs | N—90 mg kg−1 | 40 | Yu et al. [331] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byrne, L.; Murphy, R.A. Relative Bioavailability of Trace Minerals in Production Animal Nutrition: A Review. Animals 2022, 12, 1981. https://doi.org/10.3390/ani12151981
Byrne L, Murphy RA. Relative Bioavailability of Trace Minerals in Production Animal Nutrition: A Review. Animals. 2022; 12(15):1981. https://doi.org/10.3390/ani12151981
Chicago/Turabian StyleByrne, Laurann, and Richard A. Murphy. 2022. "Relative Bioavailability of Trace Minerals in Production Animal Nutrition: A Review" Animals 12, no. 15: 1981. https://doi.org/10.3390/ani12151981