
SNP Genotyping Characterizes the Genome Composition of the New Baisary Fat-Tailed Sheep Breed

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Background
2. Materials and Methods
2.1. Farm Locations
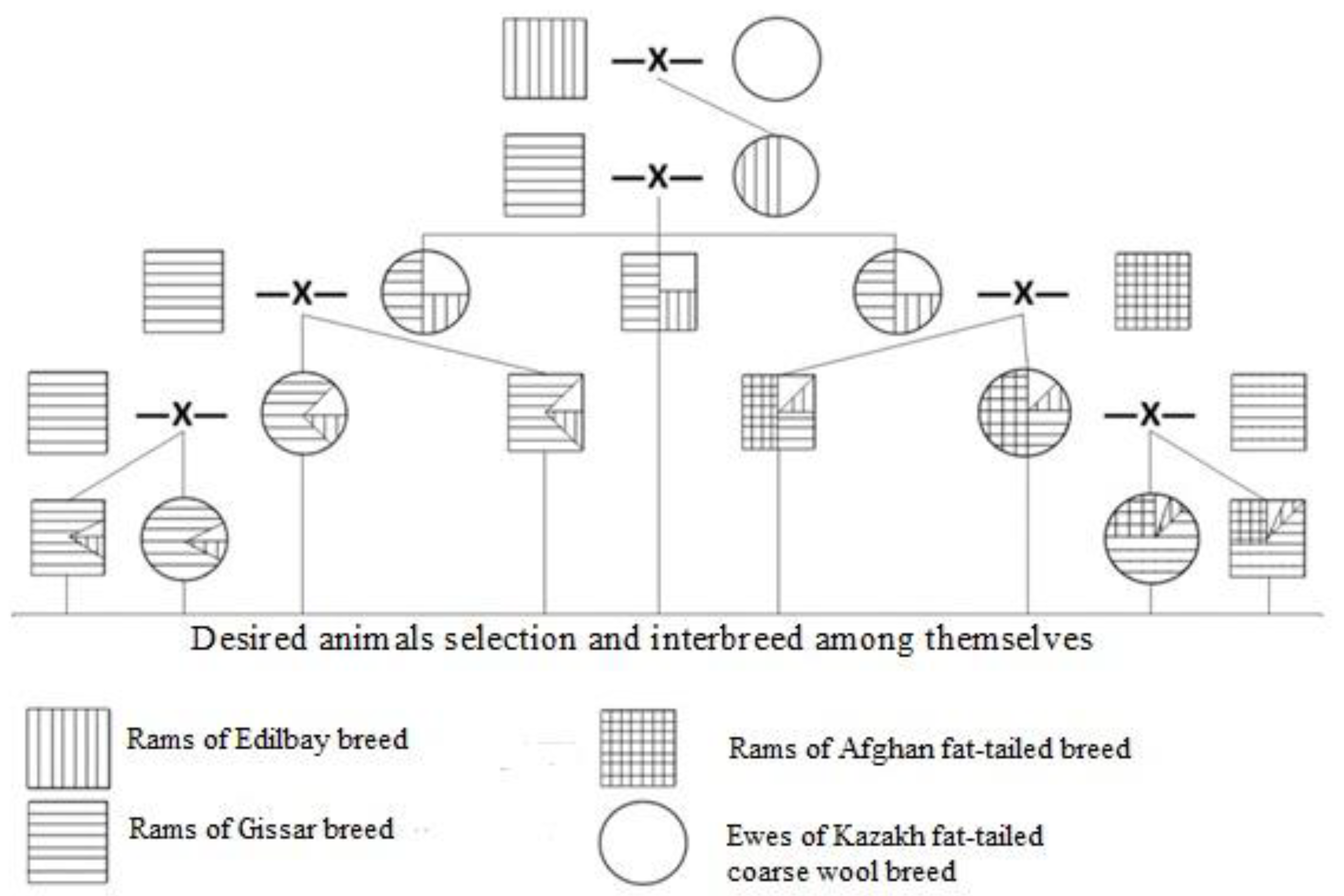
2.2. Mating Scheme
2.3. Animal Selection Criteria
2.4. Animal Sampling and SNP Genotyping
2.5. SNP Quality Control
2.6. Genetic Diversity and Population Structure Analysis
3. Results
3.1. Productive Qualities of a New Baisary Sheep of Meat-Fat Direction
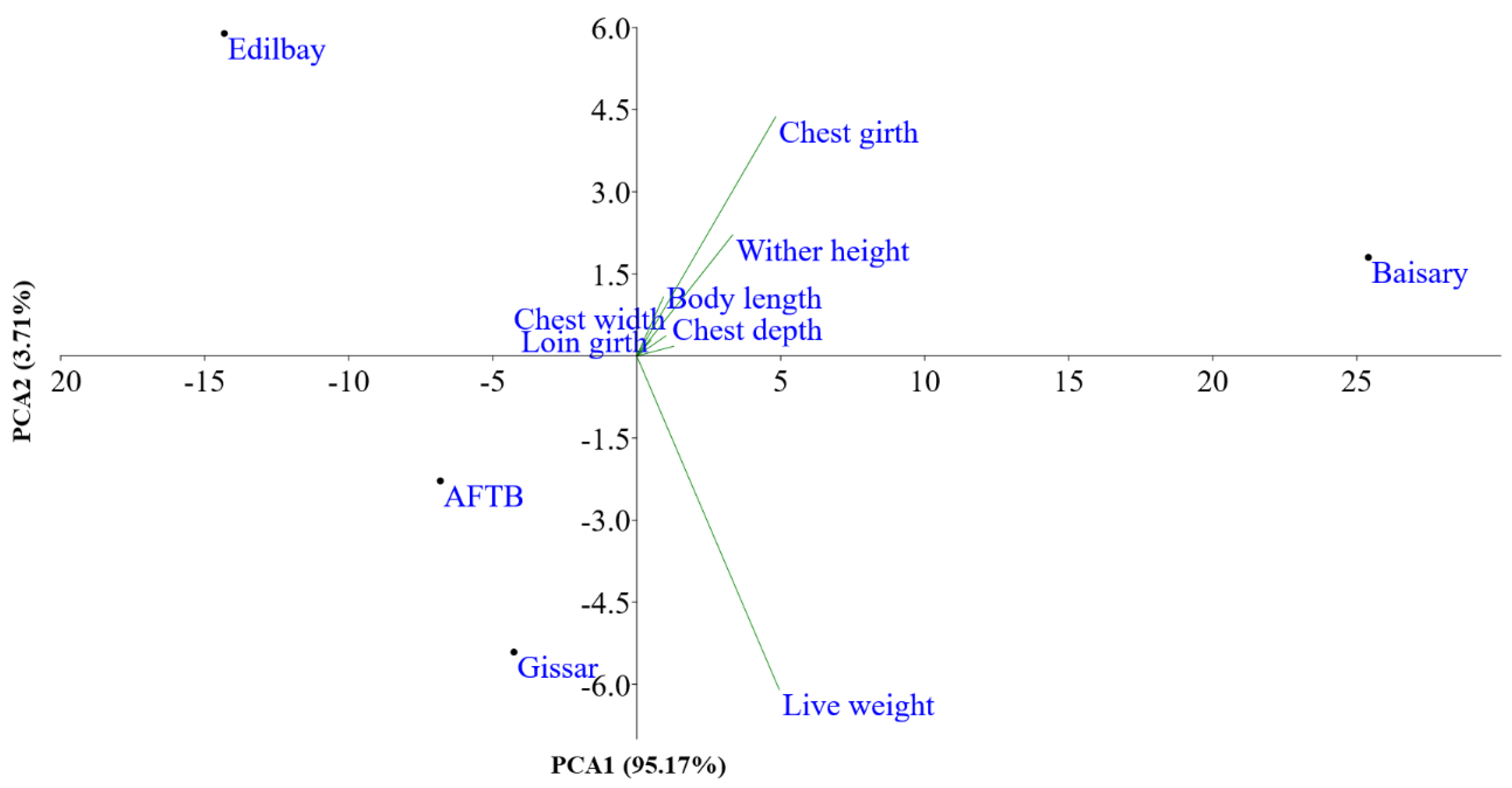
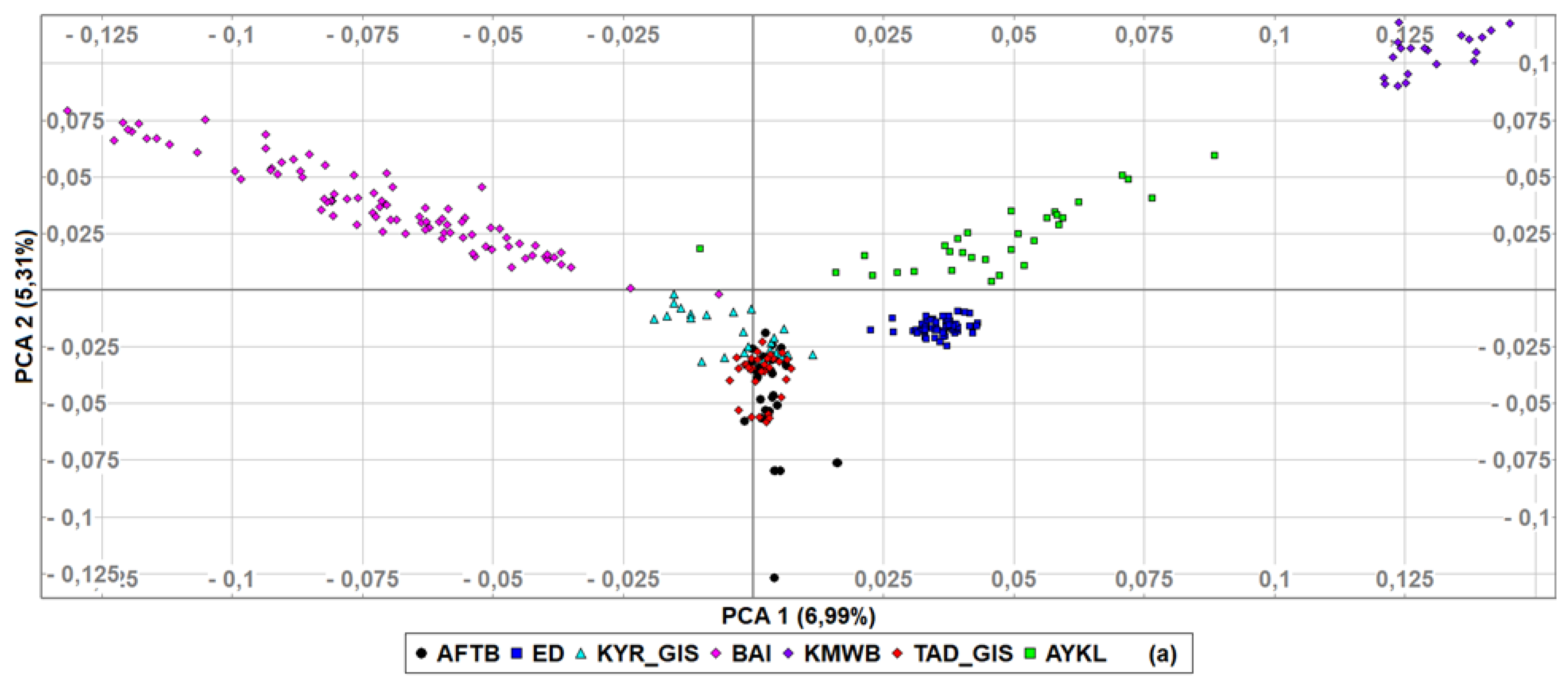
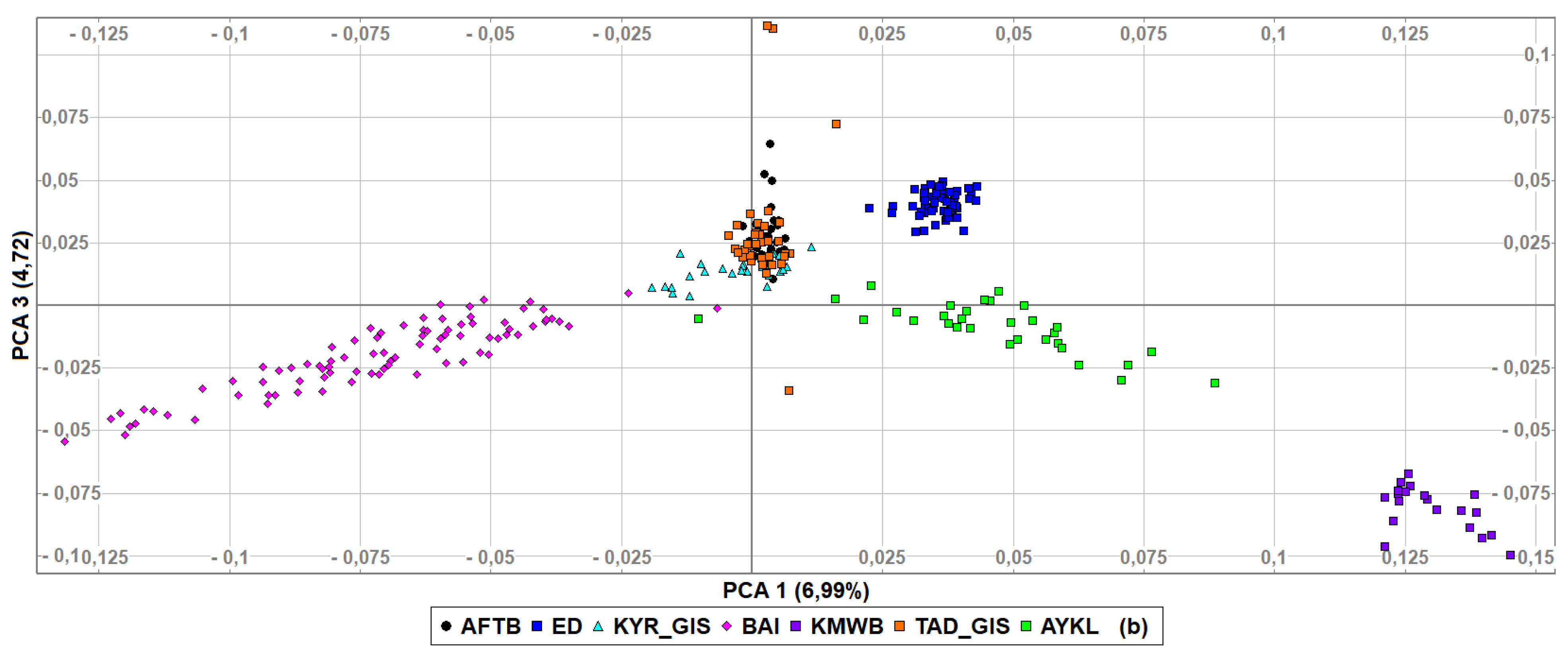
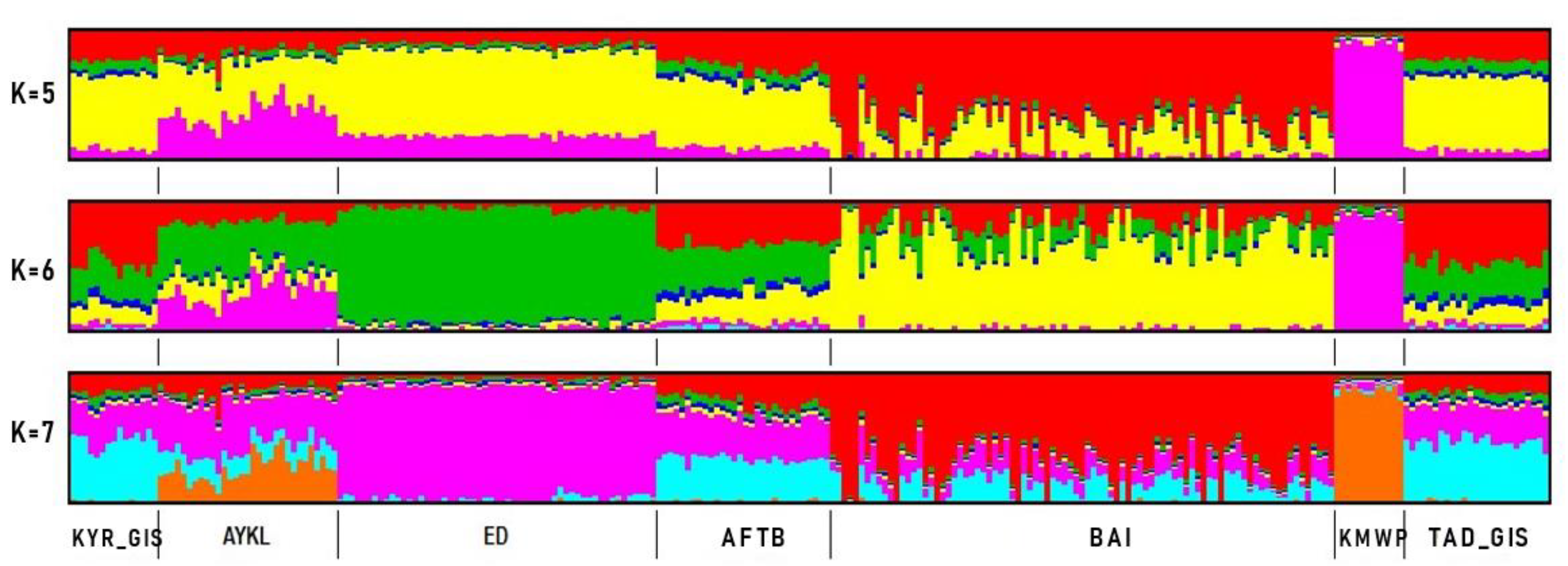
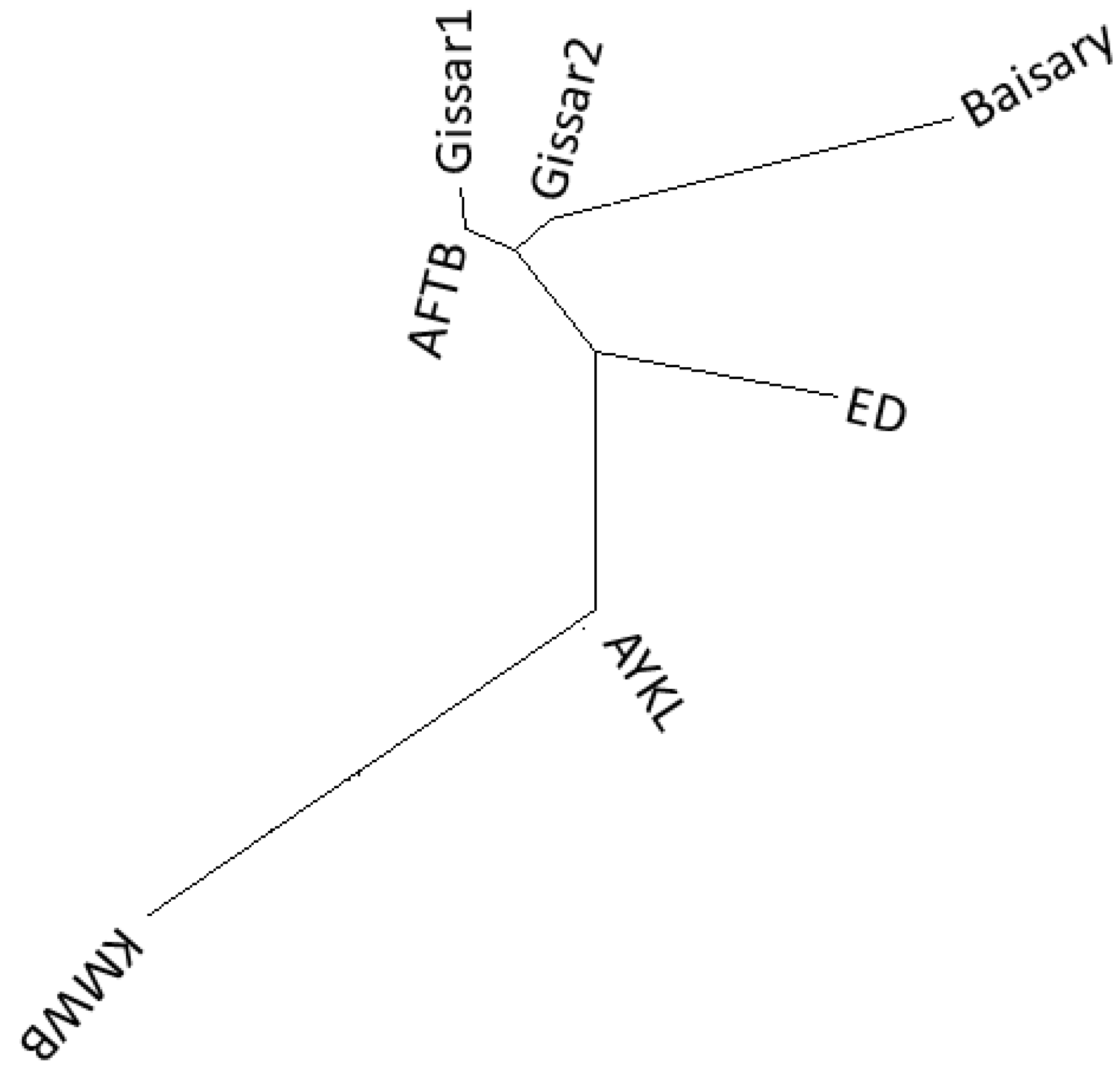
3.2. Genetic Diversity Indices and Baisary Sheep Breed’s Relationships with Ancestral Breeds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Makhatov, B.M.; Meldebekov, A.M.; Kulmanova, G.A. State and prospects for the development of sheep breeding in Kazakhstan. OshSU Bull. 2021, 1, 382–389. [Google Scholar]
- Sabdenov, K.S.; Makhatov, B.M.; Nurzhanova, K.K.; Burambayeva, N.B. Modern technology for the production of sheep products: Textbook. KazNAU.-Almaty Aitұmar 2015, 416. [Google Scholar]
- Ombaev, A.M.; Kanseitov, T.; Kanseitova, E.T.; Abzhalov, S.A. New Ordabasy breed of meat and greasy sheep. Bull. NAS RK 2014, 1, 75–79. [Google Scholar]
- Dossybayev, K.; Orazmbetova, Z.; Mussayeva, A.; Saitou, N.; Zhapbasov, R.; Makhatov, B.; Bekmanov, B. Genetic diversity of different breeds of Kazakh sheep using microsatellite analysis. Arch. Anim. Breed. 2019, 62, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Pozharskiy, A.; Khamzina, A.; Gritsenko, D.; Khamzina, Z.; Kassymbekova, S.; Karimov, N.; Karymsakov, T.; Tlevlesov, N. SNP genotyping and population analysis of five indigenous Kazakh sheep breeds. Livest. Sci. 2020, 241, 104252. [Google Scholar] [CrossRef]
- Dossybayev, K.; Mussayeva, A.; Bekmanov, B.; Kulataev, B. Analysis of Genetic Diversity in three Kazakh Sheep using 12 Microsatellites. Int. J. Eng. Technol. 2018, 7, 122. [Google Scholar] [CrossRef] [Green Version]
- Iskakova, Z.; Alibayev, N.; Burabaev, A.; Yessirkepov, M.; Marzanov, N. Characteristics of gene pool of various sheep breeds of the Republic of Kazakhstan. Eurasia J. Biosci. 2020, 14, 2395–2402. [Google Scholar]
- FAO. The State of Food and Agriculture, Livestock in the Balance; Viale delle Terme di Caracalla: Rome, Italy, 2009; Available online: http://www.fao.org/3/a-i0680e.pdf (accessed on 26 August 2021).
- Romanov, M.N.; Zinovieva, N.A.; Griffin, D.K. British Sheep Breeds as a Part of World Sheep Gene Pool Landscape: Looking into Genomic Applications. Animals 2021, 11, 994. [Google Scholar] [CrossRef]
- Kijas, J.W.; Townley, D.; Dalrymple, B.P.; Heaton, M.P.; Maddox, J.F.; McGrath, A.; Wilson, P.; Ingersoll, R.G.; McCulloch, R.; McWilliam, S.; et al. A genome wide survey of SNP variation reveals the genetic structure of sheep breeds. PLoS ONE 2009, 4, e4668. [Google Scholar] [CrossRef] [Green Version]
- Kijas, J.W.; Lenstra, J.A.; Hayes, B.; Boitard, S.; Porto Neto, L.R. Genome-Wide Analysis of the World’s Sheep Breeds Reveals High Levels of Historic Mixture and Strong Recent Selection. PLoS Biol. 2012, 10, e1001258. [Google Scholar] [CrossRef] [Green Version]
- Tobar, C.; Melissa, K.; Alvarez, L.; Alvarez Franco, D.C.; Ángela, L. Genome-wide association studies in sheep from Latin America. Review. Rev. Mex. Cienc. Pecu. 2020, 11, 859–883. [Google Scholar]
- Zhang, H.; Wang, Z.; Wang, S.; Li, H. Progress of genome wide association study in domestic animals. J. Anim. Sci. Biotechnol. 2012, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Kawȩcka, A.; Gurgul, A.; Miksza-Cybulska, A. The use of SNP microarrays for biodiversity studies of sheep—A review. Ann. Anim. Sci. 2016, 16, 975–987. [Google Scholar] [CrossRef] [Green Version]
- Deniskova, T.; Dotsev, A.; Lushihina, E.; Shakhin, A.; Kunz, E.; Medugorac, I.; Reyer, H.; Wimmers, K.; Khayatzadeh, N.; Sölkner, J.; et al. Population Structure and Genetic Diversity of Sheep Breeds in the Kyrgyzstan. Front. Genet. 2019, 10, 1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Keenan, K.; McGinnity, P.; Cross, T.F.; Crozier, W.W.; Prodohl, P.A. DiveRsity: An R package for the estimation of population genetics parameters and their associated errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef] [Green Version]
- Szpiech, Z.A.; Jackobson, N.A.; Rosenberg, N.A. ADZE: A rarefaction approach for counting alleles private to combinations of populations. Bioinformatics 2008, 24, 2498–2504. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Buchmann, R.; Hazelhurst, S. Genesis Manual, University of the Witwatersrand, Johannesburg. 2014. Available online: http://www.bioinf.wits.ac.za/software/genesis/Genesis.pdf (accessed on 30 May 2022).
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Azhimetov, N.N.; Parzhanov, Z.A.; Alibayev, N.N.; Myrzakulov, A.S. A new Ordabasy breed of sheep—A breakthrough technology for the production of lamb and mutton: Theory and practice. Eurasia J. Biosci. 2020, 14, 1193–1201. [Google Scholar]
- Zonabend König, E.; Ojango, J.M.; Audho, J.; Mirkena, T.; Strandberg, E.; Okeyo, A.M.; Philipsson, J. Live weight, conformation, carcass traits and economic values of ram lambs of Red Maasai and Dorper sheep and their crosses. Trop Anim. Health Prod. 2017, 49, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebreyowhens, W.; Mengistu, R.; Awet, E. Improving live body weight gain of local sheep through crossbreeding with high yielding exotic Dorper sheep under smallholder farmers. Int. J. Livest. Prod. 2017, 8, 67–71. [Google Scholar]
- Momani Shaker, M.; Abdullah, A.Y.; Kridli, R.T.; Šáda, I.; Sovják, R. Effect of crossing indigenous Awassi sheep breed with mutton and prolific sire breeds on growth performance of lambs in subtropical region. Czech J. Anim. Sci. 2002, 47, 247–252. [Google Scholar]
- Momani Shaker, M.; Abdullah, A.Y.; Kridli, R.T.; Bláha, J.; Šáda, I. Influence of the nutrition level on fattening and carcass characteristics of Awassi ram lambs. Czech J. Anim. Sci. 2003, 47, 466–474. [Google Scholar]
- Dawson, L.E.; Carson, R.A.F.; Mcclinton, L.O.W. Comparison of productivity of Texel and Rouge de í Ouest ewes and their crosses. Anim. Sci. 2002, 75, 459–468. [Google Scholar] [CrossRef]
- Edea, Z.; Dessie, T.; Dadi, H.; Do, K.-T.; Kim, K.-S. Genetic Diversity and Population Structure of Ethiopian Sheep Populations Revealed by High-Density SNP Markers. Front. Genet. 2017, 8, 218. [Google Scholar] [CrossRef] [Green Version]
- ATMAgro. Available online: http://atmagro.ru/2015/04/24/kurdyuchnye-ovcy/ (accessed on 24 April 2015).
- Farsykhanov, S.I. Gissar Breed of Sheep; Dushanbe: Irfon, Tajikistan, 1981; p. 238. [Google Scholar]
- Calafell, F.; Shuster, A.; Speed, W.; Kidd, J.; Kidd, K. Short tandem repeat polymorphism evolution in humans. Eur. J. Hum. Genet. 1998, 6, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, K.B.; Schurr, T.G.; Long, J.C.; Rosenberg, N.A.; Crawford, M.H.; Tarskaia, L.A.; Osipova, L.P.; Zhadanov, S.I.; Smith, D.G. A private allele ubiquitous in the Americas. Biol. Lett. 2007, 3, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Kalinowski, S.T. Counting alleles with rarefaction: Private alleles and hierarchical sampling designs. Conserv. Genet. 2004, 5, 539–543. [Google Scholar] [CrossRef]
- Deniskova, T.; Dotsev, A.; Selionova, M.; Brem, G.; Zinovieva, N. Biodiversity of Russian Local Sheep Breeds Based on Pattern of Runs of Homozygosity. Diversity 2021, 13, 360. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurements | Edilbay (ED n = 6) | Gissar (Giss1 n = 4) | Afghan Fat-Tailed Breed (AFTB n = 3) | |||
|---|---|---|---|---|---|---|
| Mean ± SE | σ | Mean ± SE | σ | Mean ± SE | σ | |
| Live weight | 115.0 ± 1.24 | 3.03 | 130.0 ± 0.41 | 0.81 | 126.0 ± 0.58 | 2.64 |
| Wither height | 86.5 ± 0.85 | 2.07 | 87.7 ± 0.75 | 1.50 | 87.0 ± 0.57 | 1.00 |
| Chest depth | 39.0 ± 0.93 | 2.28 | 40.5 ± 0.29 | 0.57 | 39.7 ± 0.33 | 0.58 |
| Chest width | 24 ± 1.06 | 2.61 | 24.5 ± 0.75 | 0.58 | 25.3 ± 0.67 | 1.15 |
| Body length | 90 ± 0.73 | 1.79 | 89.8 ± 0.85 | 1.71 | 89.3 ± 1.20 | 2.08 |
| Chest girth | 107 ± 0.73 | 1.79 | 106.8. ± 0.48 | 0.96 | 107.3 ± 0.88 | 1.53 |
| Loin girth | 9.22 ± 0.30 | 0.75 | 9.33 ± 0.25 | 0.5 | 9.7 ± 0.88 | 1.53 |
| Measurements | Adult Rams (n = 10) | 1.5 Year-Old Rams (n = 12) | 4 Month-Old Rams (n = 35) | |||
|---|---|---|---|---|---|---|
| Mean ± SE | σ | Mean ± SE | σ | Mean ± SE | σ | |
| Live weight | 143 ± 3.33 | 5.77 | 78 ± 2.91 | 5.88 | 41.4 ± 0.74 | 4.38 |
| Wither height | 102.0 ± 1.53 | 2.64 | 86.25 ± 1.32 | 2.63 | 71.9 ± 0.71 | 4.20 |
| Chest depth | 45.3 ± 0.67 | 1.15 | 35.5 ± 0.65 | 1.29 | 26.9 ± 0.45 | 2.69 |
| Chest width | 29.0 ± 0.58 | 1.0 | 22.8 ± 1.43 | 2.87 | 17.9 ± 0.27 | 1.57 |
| Body length | 94.0 ± 3.06 | 5.29 | 86.8 ± 3.90 | 7.80 | 59.4 ± 0.60 | 3.53 |
| Chest girth | 129 ± 2.64 | 4.58 | 102.5 ± 1.85 | 3.7 | 78.6 ± 0.62 | 3.68 |
| Loin girth | 11.6 ± 0.33 | 0.57 | 10.3 ± 0.25 | 0.5 | 9.5 ± 0.60 | 0.79 |
| Breeds | n | Ar | pAr | Ho | He | FIS (Cl = 95%) |
|---|---|---|---|---|---|---|
| Afghan fat-tailed breed (AFTB) | 38 | 1.99 | 0.0034 | 0.39 | 0.4 | 0.0043 (−0.026 to 0.0073) |
| Edilbay (ED) | 55 | 1.99 | 0.0036 | 0.4 | 0.39 | −0.0115 (−0.0254 to −0.017) |
| Baisary (BAI) | 87 | 1.98 | 0.0034 | 0.39 | 0.38 | −0.0099 (−0.0233 to −0.0068) |
| Kazakh meat-wool breed (KMWB) | 20 | 1.99 | 0.0047 | 0.39 | 0.4 | 0.0023 (−0.0517 to 0) |
| Gissar (Giss1) | 38 | 1.99 | 0.0030 | 0.39 | 0.39 | 0.0026 (−0.0266 to −0.00080) |
| Breed | n | Total Number of ROH | ROH Length | ROH Number | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | Min. | Max. | Mean | Min. | Max. | |||
| AFTB | 38 | 5288 | 233.171 ± 56.45 | 143.56 | 503.64 | 139.16 ± 14.2 | 91 | 165 |
| ED | 55 | 7493 | 213.6 ± 13.94 | 176.54 | 284.6 | 136.23 ± 6.05 | 121 | 153 |
| BAI | 87 | 12,170 | 267.23 ± 60.77 | 165.42 | 597.55 | 139.89 ± 7.2 | 118 | 160 |
| KMWB | 20 | 2412 | 229.84 ± 71.94 | 145.58 | 574 | 120.6 ± 18.7 | 96 | 152 |
| Gissar1 | 38 | 5515 | 259.46 ± 73.54 | 183.5 | 602.31 | 145.13 ± 8.28 | 128 | 169 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhumadillayev, N.; Dossybayev, K.; Khamzina, A.; Kapasuly, T.; Khamzina, Z.; Tlevlesov, N. SNP Genotyping Characterizes the Genome Composition of the New Baisary Fat-Tailed Sheep Breed. Animals 2022, 12, 1468. https://doi.org/10.3390/ani12111468
Zhumadillayev N, Dossybayev K, Khamzina A, Kapasuly T, Khamzina Z, Tlevlesov N. SNP Genotyping Characterizes the Genome Composition of the New Baisary Fat-Tailed Sheep Breed. Animals. 2022; 12(11):1468. https://doi.org/10.3390/ani12111468
Chicago/Turabian StyleZhumadillayev, Narzhan, Kairat Dossybayev, Aigerim Khamzina, Tilek Kapasuly, Zhangylsyn Khamzina, and Nurlan Tlevlesov. 2022. "SNP Genotyping Characterizes the Genome Composition of the New Baisary Fat-Tailed Sheep Breed" Animals 12, no. 11: 1468. https://doi.org/10.3390/ani12111468