
In Vitro and In Vivo Assessment of Dietary Supplementation of Both Natural or Nano-Zeolite in Goat Diets: Effects on Ruminal Fermentation and Nutrients Digestibility

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Feed Additives
2.2. Basal Diet
2.3. The In Vitro Experiment
2.3.1. Gas Production Procedure
2.3.2. Rumen Degradability
2.3.3. Rumen Fermentation Characteristics
2.4. In Vivo Experiment
2.4.1. Animals and Experimental Design
2.4.2. Rumen Fermentation Parameters
2.4.3. Apparent Nutrients Digestibility
2.5. Statistical Analyses
3. Results
3.1. In Vitro Experiment
3.2. In Vivo Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Grabherr, H.; Spolders, M.; Furll, M.; Flachwosky, G. Effect of several doses of zeolite A on feed intake, energy metabolism and on mineral metabolism in dairy cows around calving. J. Anim. Physiol. Anim. Nutr. 2009, 93, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Connor, E. Invited review: Improving feed efficiency in dairy production: Challenges and possibilities. Animal 2015, 9, 395–408. [Google Scholar] [CrossRef] [Green Version]
- Baglieri, A.; Reyneri, A. Organically modified clays as binders of fumonisins in feedstocks. J. Environ. Sci. Health 2013, 48, 776–783. [Google Scholar] [CrossRef]
- Jaynes, W.F.; Zartman, R.E.; Hudnall, W.H. Aflatoxin B1 adsorption by clays from water and corn meal. Appl. Clay Sci. 2007, 36, 197–205. [Google Scholar] [CrossRef]
- Valpotić, H.; Gračner, D.; Turk, R.; Đuričić, D.; Vince, S.; Folnožić, I.; Lojkić, M.; Žura Žaja, I.; Bedrica, L.; Maćešić, N.; et al. Zeolite clinoptilolite nanoporous feed additive for animals of veterinary importance: Potentials and limitations. Period. Biol. 2017, 119, 159–172. [Google Scholar] [CrossRef]
- Trckova, M.; Matlova, L.; Dvorska, L.; Pavlik, I. Kaolin, bentonite and zeolites as feed supplements for animals: Health advantages and risks. Vet. Med. Czech. 2004, 49, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Cole, N.A.; Todd, R.W.; Parker, D.B. Use of fat and zeolite to reduce ammonia emissions from beef cattle feed yards. In Proceedings of the International Symposium on Air Quality and Waste Management for Agriculture, (M31), Broomfield, CO, USA, 16–19 September 2007; ASABE: Broomfield, CO, USA, 2007; p. 41. [Google Scholar]
- Ghaemnia, L.; Bojarpour, M.; Mirzadeh, K.; Chaji, M.; Eslami, M. Effects of different levels of zeolite on digestibility and some blood parameters in Arabic lambs. J. Anim. Vet. Adv. 2010, 9, 779–781. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, M.; Nanekarani, S. The effects of calcic and potassic clinoptilolite on ruminal parameters in Lori breed sheep. APCBEE Procedia. 2012, 4, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Karatzia, M.A.; Pourliotis, K.; Katsoulos, P.D.; Karatzias, H. Effects of in-feed inclusion of clinoptilolite on blood serum concentrations of aluminium and inorganic phosphorus and on ruminal pH and volatile fatty acid concentrations in dairy cows. Biol. Trace Elem. Res. 2011, 142, 159–166. [Google Scholar] [CrossRef]
- Spotti, M.; Fracchiolla, M.L.; Arioli, F.; Caloni, F.; Pompa, G. Aflatoxin B 1 binding to sorbents in bovine ruminal fluid. Vet. Res. Commun. 2005, 29, 507–515. [Google Scholar] [CrossRef]
- Kuzma, J.; Romanchek, J.; Kokotovich, A. Upstream oversight assessment for agrifood nanotechnology: A case studies approach. Risk Anal. 2008, 28, 1081–1098. [Google Scholar] [CrossRef]
- Mahler, G.J.; Esch, M.B.; Tako, E.; Southard, T.L.; Archer, S.D.; Glahn, R.P.; Shuler, M.L. Oral exposure to polystyrene nano-particles affects iron absorption. Nat. Nanotechnol. 2012, 7, 264–271. [Google Scholar] [CrossRef]
- Zhang, D. Processing of advanced materials using high-energy mechanical milling. Prog. Mater. Sci. 2004, 49, 537–560. [Google Scholar] [CrossRef]
- Zhang, X.F.; Liu, Z.G.; Shen, W.; Gurunathan, S. Silver nanoparticles: Synthesis, characterization, properties, applications, and therapeutic approaches. Int. J. Mol. Sci. 2016, 17, 1534. [Google Scholar] [CrossRef]
- Thomas, P.; Turnbull, J.; Roberts, J.; Patel, A.; Brain, A. Response to: ‘Failed supraglottic airway’: An algorithm for suboptimally placed supraglottic airway devices based on videolaryngoscopy. Br. J. Anaesth. 2017, 119, 1243. [Google Scholar] [CrossRef]
- Varenne, F.; Botton, J.; Merlet, J.C.; Vachon, J.; Geiger, S.; Infante, I.C.; Chehimi, M.M.; Vauthier, C. Standardization and validation of a protocol of zeta potential measurements by electrophoretic light scattering for nanomaterial characterization. Colloids Surf. 2015, 486, 218–231. [Google Scholar] [CrossRef]
- NRC. National Research Council, Nutrient Requirements of Small Ruminants, 7th ed.; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- AOAC. Official Methods of Analysis, 20th ed.; Association of Official Agricultural Chemists: Arlington, VA, USA, 2006. [Google Scholar]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Bueno, I.C.S.; Cabral Filhoa, S.L.S.; Gobboa, S.P.; Louvandininb, H.; Vittia, D.M.S.S.; Abdallaa, A.L. Influence of inoculum source in a gas production method. Anim. Feed Sci. Technol. 2005, 123, 95–105. [Google Scholar] [CrossRef]
- Soltan, Y.A.; Hashem, N.M.; Morsy, A.S.; El-Azrak, K.M.; Nour El-Din, A.; Sallam, S.M. Comparative effects of Moringa oleifera root bark and monensin supplementations on ruminal fermentation, nutrient digestibility and growth performance of growing lambs. Anim. Feed Sci. Technol. 2018, 235, 189–201. [Google Scholar] [CrossRef]
- Salama, H.S.A.; El-Zaiat, H.M.; Sallam, S.M.A.; Soltan, Y.A. Agronomic and qualitative characterization of multi-cut berseem clover (Trifolium alexandrinum L.) cultivars. J. Sci. Food Agric. 2020, 100, 3857–3865. [Google Scholar] [CrossRef] [PubMed]
- Soltan, Y.A.; Morsy, A.S.; Sallam, S.M.A.; Louvandini, H.; Abdalla, A.L. Comparative in vitro evaluation of forage legumes (prosopis, acacia, atriplex, and leucaena) on ruminal fermentation and methanogenesis. J. Anim. Feed Sci. 2012, 21, 759–772. [Google Scholar] [CrossRef]
- El-Zaiat, H.M.; Abdalla, A.L. Potentials of patchouli (Pogostemon cablin) essential oil on ruminal methanogenesis, feed degradability, and enzyme activities in vitro. Environ. Sci. Pollut. Res. 2019, 26, 30220–30228. [Google Scholar] [CrossRef] [PubMed]
- Longo, C.; Bueno, I.C.S.; Nozella, E.F.; Goddoy, P.B.; Cabral-Filho, S.L.S.; Abdalla, A.L. The influence of head-space and inoculum dilution on in vitro ruminal methane measurements. Int. Congr. Ser. 2006, 1293, 62–65. [Google Scholar] [CrossRef]
- Blümmel, M.; Makkar, H.P.S.; Becker, K. In vitro gas production: A technique revisited. J. Anim. Physiol. Anim. Nutr. 1997, 77, 24–34. [Google Scholar] [CrossRef]
- Dehority, B.A.; Damron, W.S.; McLaren, J.B. Occurrence of the rumen ciliate Oligoisotricha bubali in domestic cattle (Bos taurus). Appl. Environ. Microbiol. 1983, 45, 1394–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmquist, D.L.; Conrad, H.R. Origin of plasma fatty acids in lactating cows fed high grain or high fat diets. J. Dairy Sci. 1971, 54, 1025–1033. [Google Scholar] [CrossRef]
- Konitzer, K.; Voigt, S. Direktbestimmung von Ammonium in Blut-und Gewebsextrakten mit der Phenol-Hypochlorit-reaktion. Clin. Chim. Acta 1963, 8, 5–11. [Google Scholar] [CrossRef]
- SAS. SAS User’s guide: Statistics; Version 9; SAS Institute Inc.: Cary, NC, USA, 2002. [Google Scholar]
- Hua, M.; Zhang, S.; Pan, B.; Zhang, W.; Lv, L.; Zhang, Q. Heavy metal removal from water/wastewater by nano-sized metal oxides: A review. J. Hazard. Mater. 2012, 212, 317–331. [Google Scholar] [CrossRef]
- Boadi, D.; Benchaar, C.; Chiquette, J.; Massé, D. Mitigation strategies to reduce enteric methane emissions from dairy cows: Update review. Can. J. Anim. Sci. 2004, 84, 319–335. [Google Scholar] [CrossRef]
- Sarker, N.C.; Keomanivong, F.; Borhan, M.; Rahman, S.; Swanson, K. In vitro evaluation of nano zinc oxide (nZnO) on mitigation of gaseous emissions. J. Anim. Sci. Technol. 2018, 60, 27. [Google Scholar] [CrossRef] [Green Version]
- Adegbeye, M.J.; Elghandour, M.M.; Barbabosa-Pliego, A.; Monroy, J.C.; Mellado, M. Nano-particles in equine nutrition: Mechanism of action and application as feed additives. J. Equine Vet. Sci. 2019, 78, 29–37. [Google Scholar] [CrossRef]
- Galindo, J.A.; Elias, A.; Cardero, J. The addition zeolite to silage diets. 1. Effect of zeolite level on rumen cellulolisis of cows fed silage. Cuban J. Agric. Sci. 1982, 16, 277. [Google Scholar]
- Câmara, L.R.A.; Valadares, S.C.; Leão, M.I.; Valadares, R.F.D.; Dias, M.; Gomide, A.P.C.; Barros, A.C.W.; Nascimento, V.A.; Ferreira, D.J.; Faé, J.T.; et al. Zeolite in the diet of beef cattle. Arq. Bras. Med. Vet. Zootec. 2012, 64, 631–639. [Google Scholar] [CrossRef] [Green Version]
- McCollum, F.T.; Galyean, M.L. Effects of clinoptilolite on rumen fermentation, digestion and feedlot performance in beef steers fed high concentrate diets. J. Anim. Sci. 1983, 56, 517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehority, B.A. Rumen Microbiology; Nottingham University Press: Nottingham, UK, 2003. [Google Scholar]
- Sweeney, T.F.; Cervantes, A.; Bull, L.S.; Hemken, R.W. Zeo-Agriculture. Use of Natural Zeolites in Agriculture and Aquaculture; Pond, W.G., Mumpton, F.A., Eds.; Westview Press Inc.: Boulder, CO, USA, 1984; p. 183. [Google Scholar]
- Marden, J.P.; Julien, C.; Monteils, V.; Auclair, E.; Moncoulon, R.; Bayourthe, C. How does live yeast differ from sodium bicarbonate to stabilize ruminal pH in highyielding dairy cows? J. Dairy Sci. 2008, 91, 3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dschaak, C.M.; Eun, J.-S.; Young, A.J.; Stott, R.D.; Peterson, S. Effects of supplementation of natural zeolite on intake, digestion, ruminal fermentation, and lactational performance of dairy cows. Prof. Anim. Sci. 2010, 26, 647–654. [Google Scholar] [CrossRef]
- Mumpton, F.A. La roca magica: Uses of Natural Previous Zeolites in Agriculture and Industry. Proc. Natl. Acad. Sci. USA 1999, 96, 3463–3470. [Google Scholar] [CrossRef] [Green Version]
- Erwanto, I.R.; Zakaria, W.A.; Prayuwidayati, M. The use of ammoniated zeolite to improve rumen metabolism in ruminant. Anim. Prod. Sci. 2012, 13, 138–142. [Google Scholar]
- Kardaya, D.; Sudrajat, D.; Dihansih, E. Efficacy of dietary urea-impregnated zeolite in improving rumen fermentation characteristics of local lamb. Media Paternak. 2012, 35, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Khachlouf, K.; Hamed, H.; Gdoura, R.; Gargouri, A. Effects of zeolite supplementation on dairy cow production and ruminal parameters–a review. Ann. Anim. Sci. 2018, 18, 857–877. [Google Scholar] [CrossRef] [Green Version]
- Soltan, Y.A.; Morsy, A.S.; Hashem, N.M.; Sallam, S.M. Boswellia sacra resin as a phytogenic feed supplement to enhance ruminal fermentation, milk yield, and metabolic energy status of early lactating goats. Anim. Feed Sci. Technol. 2021, 277, 114963. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
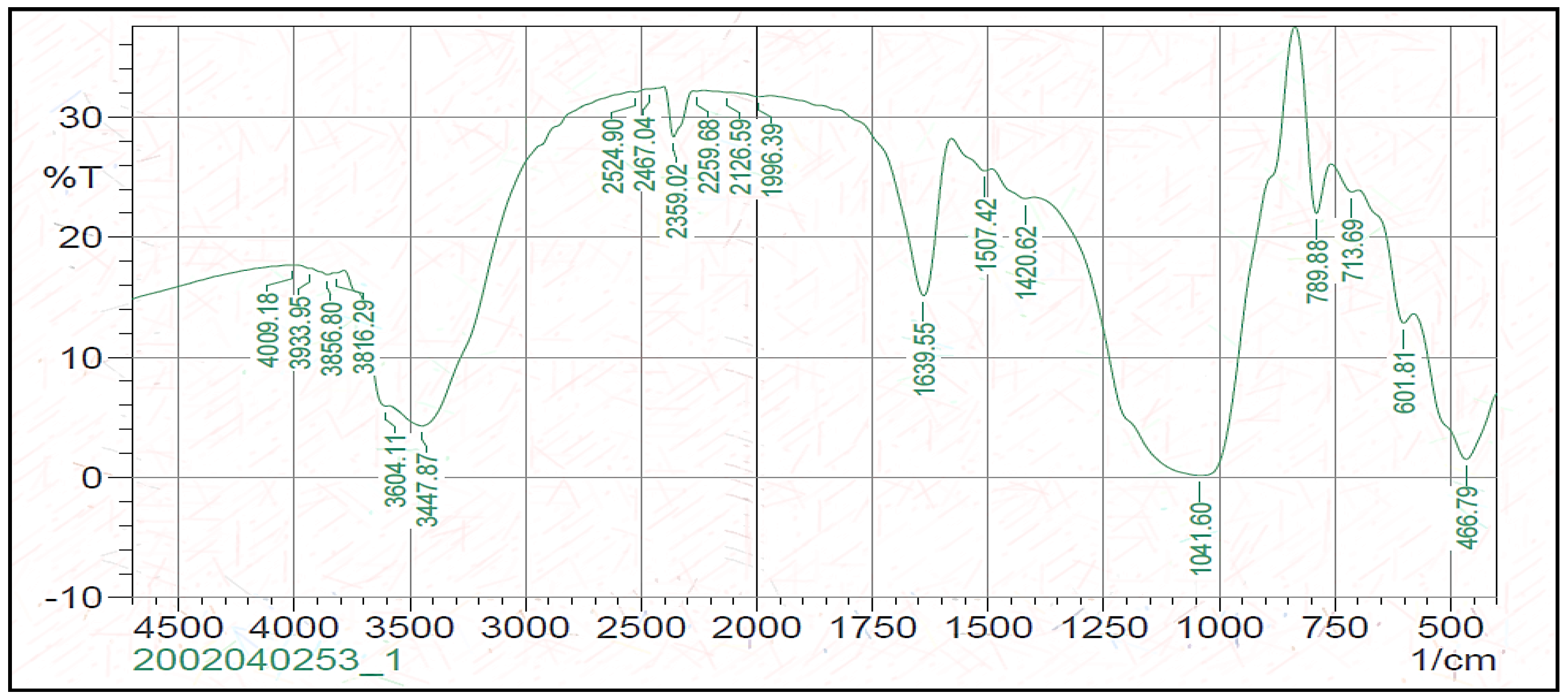{kind=link}
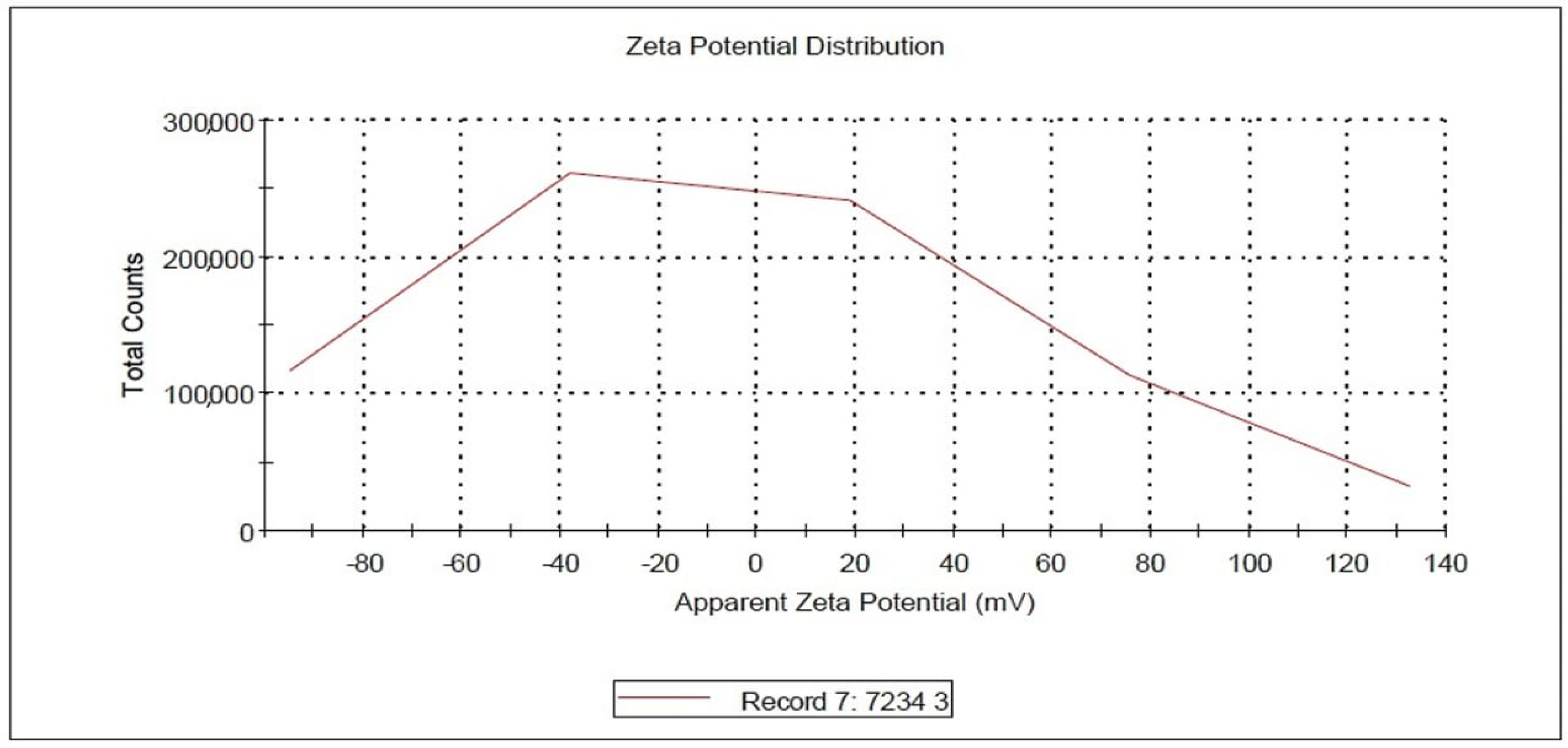{kind=link}
| Item | Zeolite Characteristics |
|---|---|
| Chemical composition | |
| SiO2 | 650.0–713.0 g/kg |
| Al2O3 | 115.0–131.0 g/kg |
| CaO | 27.0–52.0 g/kg |
| K2O | 22.0–34.0 g/kg |
| Fe2O3 | 7.00–19.0 g/kg |
| MgO | 6.00–12.0 g/kg |
| Na2O | 2.00–13.0 g/kg |
| TiO2 | 1.00–3.00 g/kg |
| Si/Al ratio | 4.80–5.40 |
| Physical properties | |
| Softing point | 1260 °C |
| Melting point | 1340 °C |
| Flow temperature | 1420 °C |
| Specific gravity | 2200–2440 kg/m3 |
| Volume density | 1600–1800 kg/m3 |
| Porosity | 24–32% |
| Compactness | 70% |
| Whitens | 70% |
| Appearance | Gray-green |
| Item | Basal Diet (g/kg Dry Matter) |
|---|---|
| Ingredients | |
| Berseem clover hay | 500 |
| Ground yellow corn | 345 |
| Soybean meal | 150 |
| Mineral and vitamin mixture 1 | 5.00 |
| Chemical composition (g/kg DM) | |
| Organic matter | 924 |
| Crude protein | 131 |
| Ether extract | 20.0 |
| Neutral detergent fiber | 718 |
| Acid detergent fiber | 343 |
| Acid detergent lignin | 60 |
| Hemicellulose | 375 |
| Cellulose | 283 |
| Treatment | GP (mL/g DM Incubated) | Methane | TDOM (g/kg) | Partitioning Factor (mg TDOM/mL GP) | |
|---|---|---|---|---|---|
| (mL/g DM Incubated) | (mL/g TDOM) | ||||
| Control | 133 | 7.7 | 10.8 | 709 | 1.10 |
| Nano zeolite (g/kg DM) | |||||
| 0.20 | 143 | 10.51 | 14.33 | 730 | 1.11 |
| 0.30 | 129 | 10.12 | 13.80 | 733 | 1.13 |
| 0.40 | 142 | 6.74 | 9.13 | 756 | 1.18 |
| 0.50 | 153 | 6.94 | 9.90 | 734 | 1.09 |
| 1.00 | 141 | 7.32 | 10.2 | 742 | 0.97 |
| Contrast 1 | |||||
| SEM | 1.40 | 0.22 | 0.38 | 8.19 | 0.01 |
| Linear | 0.36 | 0.01 | 0.05 | 0.09 | 0.56 |
| Quadratic | 0.004 | 0.04 | 0.10 | 0.46 | 0.86 |
| Natural zeolite (g/kg DM) | |||||
| 10 | 138 | 4.99 | 7.40 | 710 | 1.15 |
| 20 | 137 | 3.98 | 5.50 | 741 | 1.13 |
| 30 | 140 | 5.15 | 7.25 | 711 | 1.16 |
| Contrast 2 | |||||
| SEM | 0.42 | 0.06 | 0.11 | 2.45 | 0.002 |
| Linear | 0.25 | 0.002 | 0.001 | 0.09 | 0.62 |
| Quadratic | 0.36 | 0.16 | 0.24 | 0.36 | 0.50 |
| Treatment | Protozoal Count (×105/mL) | ||||||
|---|---|---|---|---|---|---|---|
| Diplodinium | Entodinium | Epidinium | Eudiplodinium | Isotricha | Ophryscolex | Total | |
| Control | 8.49 | 1.42 | 0.412 | 0.150 | 0.26 | 0.150 | 10.9 |
| Nano zeolite (g/kg DM) | |||||||
| 0.20 | 10.4 | 1.20 | 0.07 | 0.11 | 0.15 | 0.22 | 12.1 |
| 0.30 | 8.62 | 1.12 | 0.15 | 0.37 | 0.15 | 0.11 | 10.5 |
| 0.40 | 12.9 | 1.53 | 0.07 | 0.30 | 0.41 | 0.15 | 15.4 |
| 0.50 | 10.0 | 1.46 | 0.01 | 0.67 | 0.30 | 0.07 | 12.5 |
| 1.00 | 11.5 | 1.39 | 0.03 | 0.90 | 0.41 | 0.11 | 14.3 |
| Contrast 1 | |||||||
| SEM | 0.39 | 0.143 | 0.045 | 0.082 | 0.098 | 0.052 | 0.44 |
| Linear | 0.86 | 0.28 | 0.01 | 0.18 | 0.49 | 0.71 | 0.67 |
| Quadratic | 0.009 | 0.75 | 0.03 | 0.31 | 0.68 | 0.28 | 0.05 |
| Natural zeolite (g/kg DM) | |||||||
| 10 | 11.7 | 1.20 | 0.07 | 0.11 | 0.94 | 0.11 | 14.2 |
| 20 | 13.5 | 1.01 | 0.11 | 0.83 | 0.71 | 0.30 | 16.5 |
| 30 | 11.6 | 1.20 | 0.04 | 0.22 | 0.37 | 0.26 | 13.7 |
| Contrast 2 | |||||||
| SEM | 0.124 | 0.043 | 0.013 | 0.024 | 0.029 | 0.015 | 0.1334 |
| Linear | <0.001 | 0.20 | 0.04 | 0.001 | 0.09 | 0.21 | <0.001 |
| Quadratic | 0.36 | 0.95 | 0.13 | 0.03 | 0.05 | 0.27 | 0.61 |
| Item | SCFAs (% of Total SCFAs) | Total SCFAs (mM) | pH | NH3-N (mg/100 mL) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Acetate | Propionate | Butyrate | Iso-butyrate | Valerate | Iso-Valerate | C2:C3 | ||||
| Control | 59.1 | 22.2 | 11.6 | 0.297 | 2.02 | 3.31 | 2.66 | 90.0 | 6.34 | 20.7 |
| Nano zeolite (g/kg DM) | ||||||||||
| 0.20 | 58.4 | 24.9 | 11.1 | 0.276 | 1.96 | 3.30 | 2.34 | 103 | 6.31 | 21.1 |
| 0.30 | 58.7 | 24.8 | 10.9 | 0.282 | 1.95 | 3.37 | 2.37 | 99.0 | 6.32 | 19.3 |
| 0.40 | 58.6 | 24.4 | 11.3 | 0.309 | 1.97 | 3.26 | 2.36 | 94.0 | 6.26 | 16.3 |
| 0.50 | 60.3 | 24.7 | 10.2 | 0.209 | 1.71 | 2.85 | 2.44 | 95.0 | 6.25 | 19.8 |
| 1.00 | 59.7 | 23.1 | 11.4 | 0.310 | 1.99 | 3.39 | 2.59 | 89.0 | 6.32 | 20.6 |
| Contrast 1 | ||||||||||
| SEM | 0.631 | 0.316 | 0.622 | 0.033 | 0.074 | 0.146 | 0.041 | 3.150 | 0.012 | 0.475 |
| Linear | 0.71 | 0.007 | 0.56 | 0.82 | 0.68 | 0.87 | 0.01 | 0.29 | 0.54 | 0.27 |
| Quadratic | 0.65 | 0.06 | 0.86 | 0.82 | 0.88 | 0.91 | 0.07 | 0.24 | 0.49 | 0.33 |
| Natural zeolite (g/kg DM) | ||||||||||
| 10 | 63.7 | 21.6 | 9.88 | 0.18 | 1.61 | 3.03 | 2.82 | 83.0 | 6.36 | 20.3 |
| 20 | 59.4 | 24.8 | 10.7 | 0.21 | 1.75 | 2.94 | 2.39 | 100 | 6.33 | 17.9 |
| 30 | 58.2 | 23.4 | 12.6 | 0.32 | 1.95 | 3.40 | 2.49 | 103 | 6.34 | 19.2 |
| Contrast 2 | ||||||||||
| SEM | 0.19 | 0.09 | 0.186 | 0.010 | 0.022 | 0.044 | 0.012 | 0.94 | 0.004 | 0.14 |
| Linear | 0.86 | 0.001 | 0.64 | 0.43 | 0.16 | 0.47 | 0.10 | 0.14 | 0.83 | 0.004 |
| Quadratic | 0.03 | 0.08 | 0.421 | 0.44 | 0.11 | 0.82 | 0.02 | 0.05 | 0.15 | 0.13 |
| Items | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| Control | Natural Zeolite | Nano Zeolite | |||
| Total SCFAs, mM | 72.3 b | 74.3 b | 86.8 a | 2.48 | 0.02 |
| SCFAs (% of total SCFAs) | |||||
| Acetic | 64.3 | 65.8 | 61.6 | 0.93 | 0.20 |
| Propionic | 17.1 | 15.7 | 16.0 | 0.56 | 0.50 |
| Isobutyric | 2.13 | 2.30 | 1.93 | 0.10 | 0.41 |
| Butyric | 9.92 b | 10.2 b | 14.3 a | 0.66 | 0.001 |
| Isovaleric | 4.37 | 3.68 | 4.45 | 0.42 | 0.76 |
| Valeric | 2.24 ab | 2.46 a | 1.74 b | 0.12 | 0.03 |
| C2:C3 | 3.83 | 4.25 | 3.95 | 0.19 | 0.69 |
| pH | 5.50 b | 5.96 a | 5.72 ab | 0.07 | 0.005 |
| NH3-N, (mg/100 mL) | 6.28 a | 5.16 b | 3.90 c | 0.30 | 0.001> |
| Protozoa, (×105/mL) | |||||
| Diplodinium | 10.33 | 12.3 | 11.4 | 0.39 | 0.113 |
| Entodinium | 1.37 | 1.50 | 1.11 | 0.09 | 0.212 |
| Epidinium | 1.07 | 1.13 | 1.17 | 0.11 | 0.932 |
| Eudiplodinium | 0.57 | 0.70 | 0.63 | 0.08 | 0.793 |
| Isotrica | 0.23 c | 0.77 a | 0.50 b | 0.06 | <0.001 |
| Ophryscolex | 0.17 | 0.13 | 0.40 | 0.06 | 0.123 |
| Total | 13.7 | 16.8 | 15.3 | 0.65 | 0.147 |
| Items | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| Control | Natural Zeolite | Nano Zeolite | |||
| DMI (g/day) | 1167 | 1184 | 1179 | 13.21 | 0.904 |
| Digestibility (g/kg) | |||||
| Dry matter | 443 | 446 | 445 | 0.13 | 0.56 |
| Organic matter | 435 | 459 | 445 | 0.61 | 0.31 |
| Ether extract | 556 | 565 | 608 | 1.42 | 0.30 |
| Crude protein | 387 | 386 | 425 | 0.90 | 0.12 |
| Neutral detergent fiber | 429 | 436 | 425 | 0.43 | 0.62 |
| Acid detergent fiber | 355 | 362 | 341 | 0.66 | 0.45 |
| Hemicellulose | 505 | 513 | 513 | 0.44 | 0.764 |
| Cellulose | 472 | 467 | 447 | 0.75 | 0.389 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Nile, A.; Elazab, M.; El-Zaiat, H.; El-Azrak, K.E.-D.; Elkomy, A.; Sallam, S.; Soltan, Y. In Vitro and In Vivo Assessment of Dietary Supplementation of Both Natural or Nano-Zeolite in Goat Diets: Effects on Ruminal Fermentation and Nutrients Digestibility. Animals 2021, 11, 2215. https://doi.org/10.3390/ani11082215
El-Nile A, Elazab M, El-Zaiat H, El-Azrak KE-D, Elkomy A, Sallam S, Soltan Y. In Vitro and In Vivo Assessment of Dietary Supplementation of Both Natural or Nano-Zeolite in Goat Diets: Effects on Ruminal Fermentation and Nutrients Digestibility. Animals. 2021; 11(8):2215. https://doi.org/10.3390/ani11082215
Chicago/Turabian StyleEl-Nile, Amr, Mahmoud Elazab, Hani El-Zaiat, Kheir El-Din El-Azrak, Alaa Elkomy, Sobhy Sallam, and Yosra Soltan. 2021. "In Vitro and In Vivo Assessment of Dietary Supplementation of Both Natural or Nano-Zeolite in Goat Diets: Effects on Ruminal Fermentation and Nutrients Digestibility" Animals 11, no. 8: 2215. https://doi.org/10.3390/ani11082215