
Schizochytrium sp. (T18) Oil as a Fish Oil Replacement in Diets for Juvenile Rainbow Trout (Oncorhynchus mykiss): Effects on Growth Performance, Tissue Fatty Acid Content, and Lipid-Related Transcript Expression

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Microbial Oil
2.2. Experimental Diet Formulation and Composition
2.3. Experimental Fish
2.4. Tissue Sampling
2.5. Growth Performance
2.6. Whole Body Nutrient Composition
2.7. Total Lipid and FA Content of Tissues
2.8. RNA Preparation
2.9. Real-Time Quantitative Polymerase Chain Reaction (qPCR)
2.9.1. qPCR Overview
2.9.2. Primer Design
2.9.3. cDNA Synthesis and qPCR Parameters
2.9.4. Primer Quality Assurance Testing
2.9.5. Endogenous Control (Normalizer) Selection
2.9.6. Experimental qPCR Analyses
2.10. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Fatty Acid Content of the Liver
3.3. Fatty Acid Content of Muscle
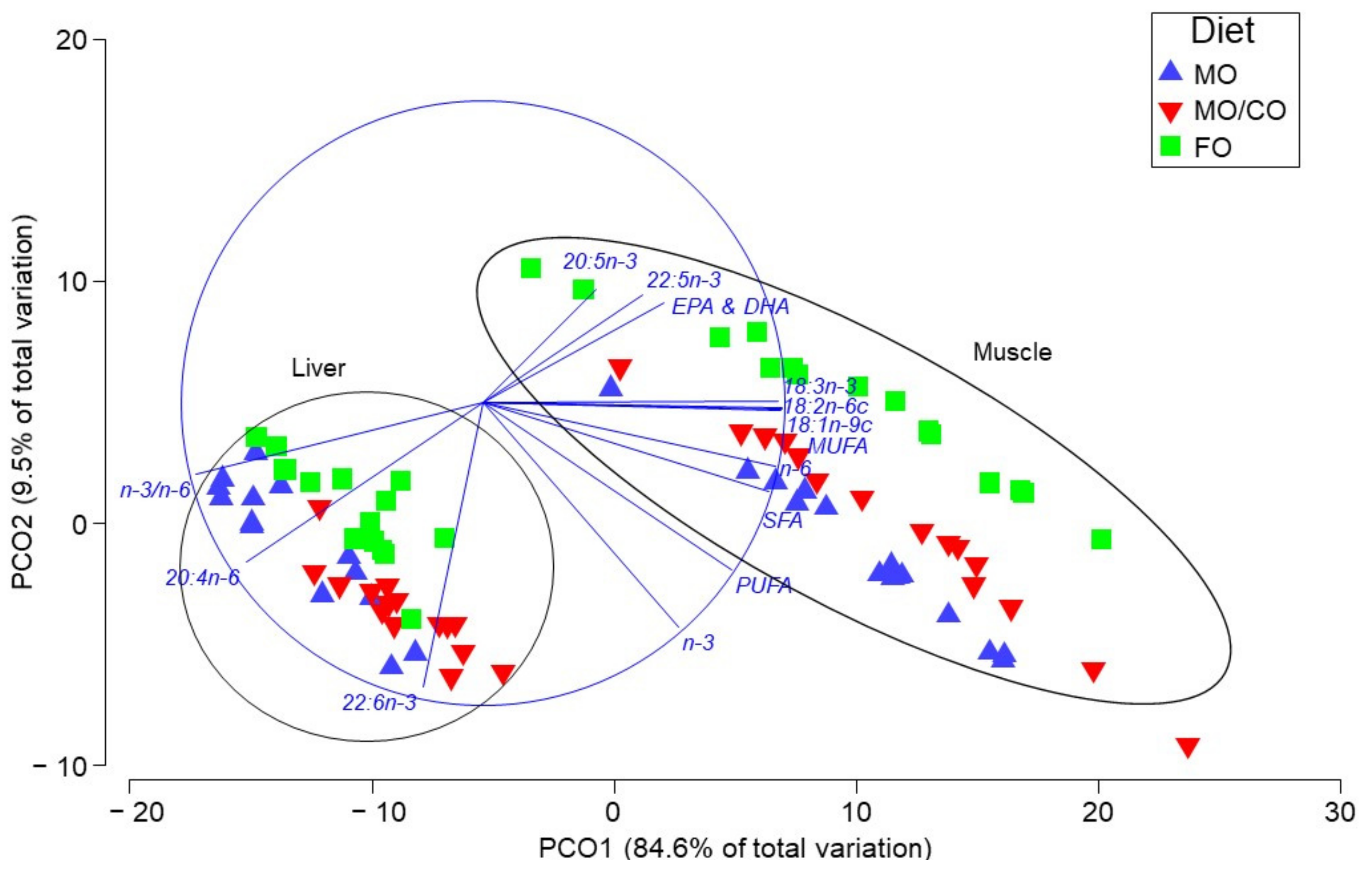
3.4. Multivariate Analyses of Fatty Acid Data
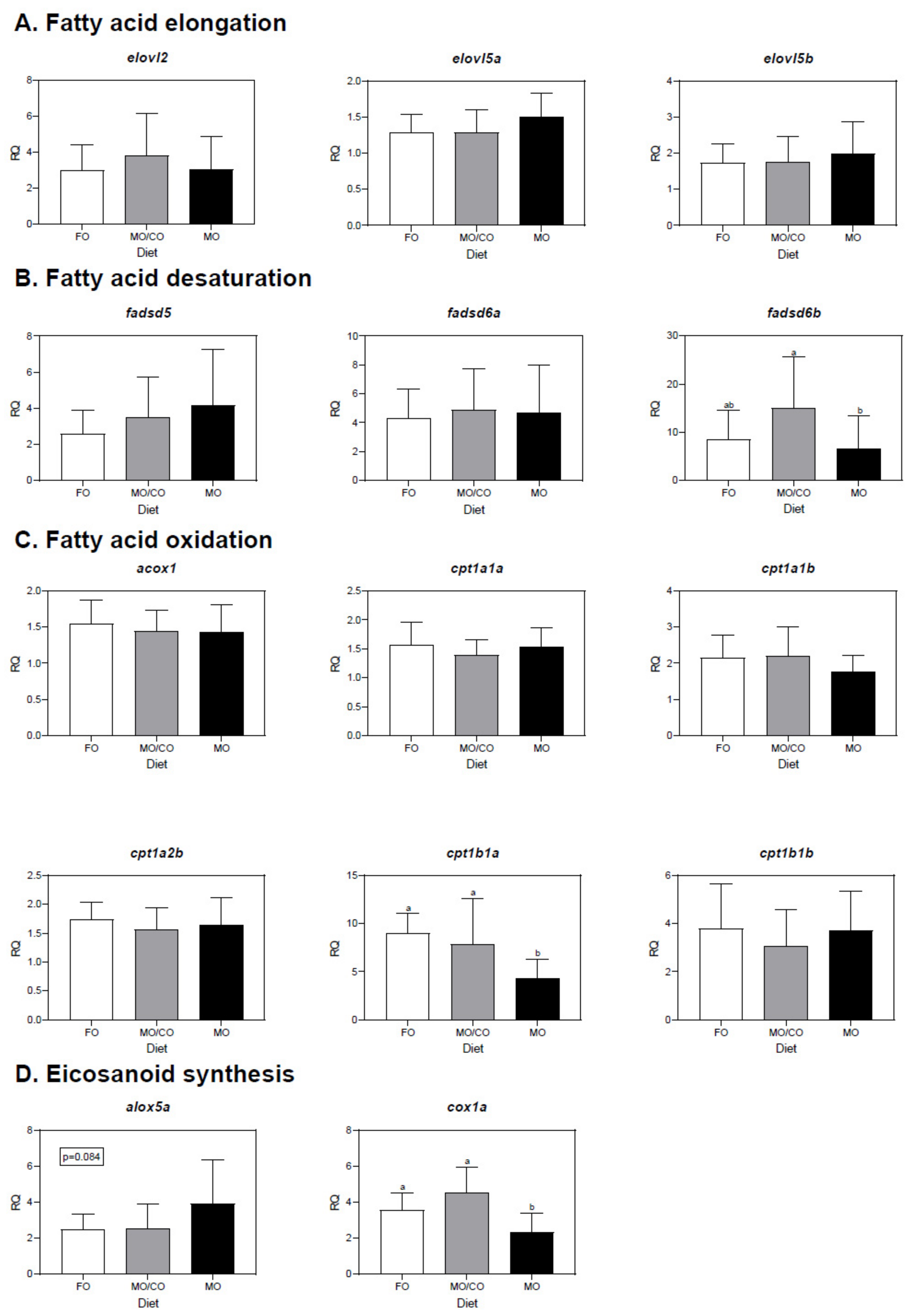
3.5. Liver Transcript Expression Analysis
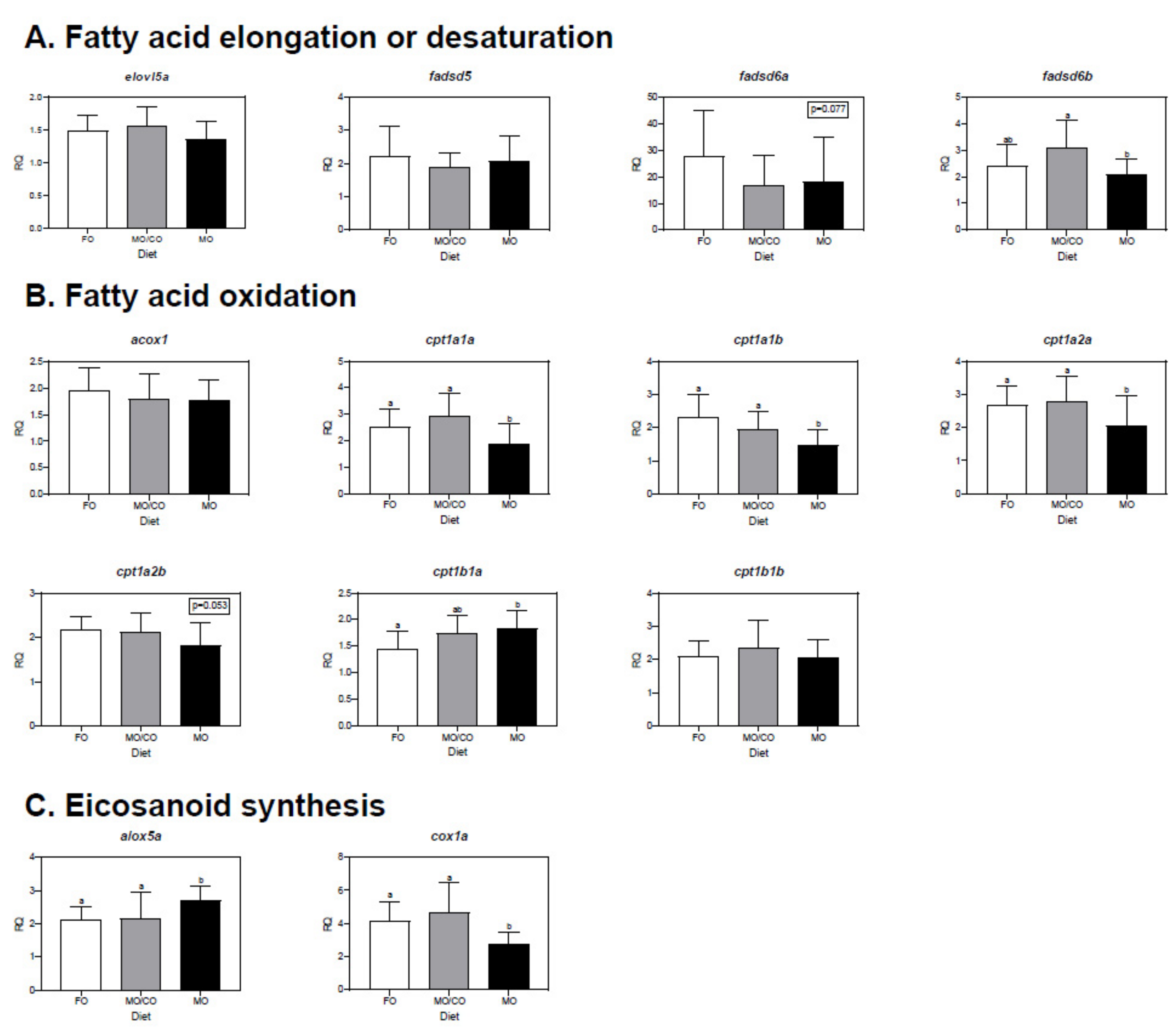
3.6. Muscle Transcript Expression Analysis
3.7. Multivariate Statistics and Correlations of Fatty Acid and Transcript Datat
4. Discussion
4.1. Impact on Growth Performance and Whole Body Nutrient Content
4.2. Impact on Liver FA Content
4.3. Impact on Muscle FA Content
4.4. Multivariate Analysis of FA profiles
4.5. Transcript Expression in Liver
4.6. Transcript Expression in Muscle
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tocher, D.R.; Betancor, M.B.; Sprague, M.; E Olsen, R.; A Napier, J. Omega-3 Long-Chain Polyunsaturated Fatty Acids, EPA and DHA: Bridging the Gap between Supply and Demand. Nutrients 2019, 11, 89. [Google Scholar] [CrossRef] [Green Version]
- Colombo, S.M.; Rodgers, T.F.M.; Diamond, M.L.; Bazinet, R.P.; Arts, M.T. Projected declines in global DHA availability for human consumption as a result of global warming. Ambio 2020, 49, 865–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tocher, D.R.; Bell, J.G.; Dick, J.R.; Crampton, V.O. Effects of dietary vegetable oil on Atlantic salmon hepatocyte fatty acid desaturation and liver fatty acid compositions. Lipids 2003, 38, 723–732. [Google Scholar] [CrossRef] [PubMed]
- NRC (National Research Council); Committee on the Nutrient Requirements of Fish Shrimp. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Da Silva, T.L.; Moniz, P.; Silva, C.; Reis, A. The Dark Side of Microalgae Biotechnology: A Heterotrophic Biorefinery Platform Directed to ω-3 Rich Lipid Production. Microorganisms 2019, 7, 670. [Google Scholar] [CrossRef] [Green Version]
- Tocher, D.R. Omega-3 long-chain polyunsaturated fatty acids and aquaculture in perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- Bazan, N.G.; Molina, M.F.; Gordon, W.C. Docosahexaenoic Acid Signalolipidomics in Nutrition: Significance in Aging, Neuroinflammation, Macular Degeneration, Alzheimer’s, and Other Neurodegenerative Diseases. Annu. Rev. Nutr. 2011, 31, 321–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, L.B.S.; Ciappolino, V.; Agostoni, C. DHA Effects in Brain Development and Function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Ryan, A.S.; Astwood, J.D.; Gautier, S.; Kuratko, C.N.; Nelson, E.B.; Salem, N. Effects of long-chain polyunsaturated fatty acid supplementation on neurodevelopment in childhood: A review of human studies. Prostaglandins Leukot. Essent. Fat. Acids 2010, 82, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Jacob-Lopes, E.; Maroneze, M.M.; Queiroz, M.I.; Zepka, L.Q. (Eds.) Handbook of Microalgae-Based Processes and Products: Fun-damentals and Advances in Energy, Food, Feed, Fertilizer, and Bioactive Compounds; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Nichols, D.S. Prokaryotes and the input of polyunsaturated fatty acids to the marine food web. FEMS Microbiol. Lett. 2003, 219, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Yarnold, J.; Karan, H.; Oey, M.; Hankamer, B. Microalgal Aquafeeds As Part of a Circular Bioeconomy. Trends. Plant. Sci.. 2019, 24, 959–970. [Google Scholar] [CrossRef] [Green Version]
- Adarme-Vega, T.C.; Lim, D.K.Y.; Timmins, M.; Vernen, F.; Felicitas, V.; Schenk, P.M. Microalgal biofactories: A promising approach towards sustainable omega-3 fatty acid production. Microb. Cell Factories 2012, 11, 96. [Google Scholar] [CrossRef] [Green Version]
- Tibbetts, S.M.; Scaife, M.A.; Armenta, R.E. Apparent digestibility of proximate nutrients, energy and fatty acids in nutritionally-balanced diets with partial or complete replacement of dietary fish oil with microbial oil from a novel Schizochytrium sp. (T18) by juvenile Atlantic salmon (Salmo salar L.). Aquaculture 2020, 520, 735003. [Google Scholar] [CrossRef]
- Desvilettes, C.; Bec, A. Formation and Transfer of Fatty Acids in Aquatic Microbial Food Webs: Role of Heterotrophic Protists. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M., Eds.; Springer: New York, NY, USA, 2009; pp. 25–42. [Google Scholar]
- Gong, Y.; Wan, X.; Jiang, M.; Hu, C.; Hu, H.; Huang, F. Metabolic engineering of microorganisms to produce omega-3 very long-chain polyunsaturated fatty acids. Prog. Lipid Res. 2014, 56, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Karageorgou, D.; Rova, E.; Katapodis, P.; Rova, U.; Christakopoulos, P.; Matsakas, L. An Overview of Potential Oleaginous Microorganisms and Their Role in Biodiesel and Omega-3 Fatty Acid-Based Industries. Microorganisms 2020, 8, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, Z.; Ratledge, C. (Eds.) Single Cell Oils: Microbial and Algal Oils; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Sprague, M.; Walton, J.; Campbell, P.; Strachan, F.; Dick, J.R.; Bell, J.G. Replacement of fish oil with a DHA-rich algal meal derived from Schizochytrium sp. on the fatty acid and persistent organic pollutant levels in diets and flesh of Atlantic salmon (Salmo salar, L.) post-smolts. Food Chem. 2015, 185, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Parrish, C.C.; Guerra, N.I.; Armenta, R.E.; Colombo, S.M. Extracted microbial oil from a novel Schizochytrium sp. (T18) as a sustainable high DHA source for Atlantic salmon feed: Impacts on growth and tissue lipids. Aquaculture 2021, 534, 736249. [Google Scholar] [CrossRef]
- Leyland, B.; Leu, S.; Boussiba, S. Are Thraustochytrids algae? Fungal Biol. 2017, 121, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Damare, V.S. Advances in isolation and preservation strategies of ecologically important marine protists, the thraustochytrids. In Advances in Biological Science Research; Elsevier: Amsterdam, The Netherlands, 2019; pp. 500. [Google Scholar]
- Morabito, C.; Bournaud, C.; Maës, C.; Schuler, M.; Cigliano, R.A.; Dellero, Y.; Maréchal, E.; Amato, A.; Rébeillé, F. The lipid metabolism in thraustochytrids. Prog. Lipid Res. 2019, 76, 101007. [Google Scholar] [CrossRef] [PubMed]
- Metz, J.G.; Roessler, P.; Facciotti, D.; Levering, C.; Dittrich, F.; Lassner, M.; Valentine, R.; Lardizabal, K.; Domergue, F.; Yamada, A.; et al. Production of Polyunsaturated Fatty Acids by Polyketide Synthases in Both Prokaryotes and Eukaryotes. Science 2001, 293, 290–293. [Google Scholar] [CrossRef]
- Ratledge, C.; Streekstra, H.; Cohen, Z.; Fichtali, J. Downstream Processing, Extraction, and Purification of Single Cell Oils. In Single Cell Oils; Elsevier: Amsterdam, The Netherlands, 2010; pp. 179–197. [Google Scholar]
- Tibbetts, S.M. The potential for ‘next-generation’, microalgae-based feed ingredients for salmonid aquaculture in context of the blue revolution. In Microalgal Biotechnology; Jacob- Lopes, E., Ed.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Hart, B.; Schurr, R.; Narendranath, N.; Kuehnle, A.; Colombo, S. Digestibility of Schizochytrium sp. whole cell biomass by Atlantic salmon (Salmo salar). Aquaculture 2021, 533, 736156. [Google Scholar] [CrossRef]
- Miller, M.R.; Nichols, P.D.; Carter, C.G. Replacement of fish oil with thraustochytrid Schizochytrium sp. L oil in Atlantic salmon parr (Salmo salar L) diets. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2007, 148, 382–392. [Google Scholar] [CrossRef]
- Santigosa, E.; Constant, D.; Prudence, D.; Wahli, T.; Verlhac-Trichet, V. A novel marine algal oil containing both EPA and DHA is an effective source of omega-3 fatty acids for rainbow trout ( Oncorhynchus mykiss ). J. World Aquac. Soc. 2020, 51, 649–665. [Google Scholar] [CrossRef]
- FAO. FAO Yearbook. Fishery and Aquaculture Statistics 2018. 2020. Available online: http://www.fao.org/fishery/static/Yearbook/YB2018_USBcard/root/aquaculture/b23.pdf (accessed on 15 April 2021).
- DFO (Department of Fisheries and Oceans Canada). Canadian Aquaculture Production and Value. 2018. Available online: https://www.dfo-mpo.gc.ca/stats/aqua/aqua18-eng.htm (accessed on 15 April 2021).
- USDA (United States Department of Agriculture). Trout Production. Released February 27, 2020; National Agricultural Statistics Service (NASS), Agricultural Statistics Board, United States Department of Agriculture: Washington, DC, USA, 2020; pp. 1–10. [Google Scholar]
- Hixson, S.M.; Parrish, C.C.; Anderson, D.M. Changes in Tissue Lipid and Fatty Acid Composition of Farmed Rainbow Trout in Response to Dietary Camelina Oil as a Replacement of Fish Oil. Lipids 2013, 49, 97–111. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipides from animal tis-sues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Christie, W.W.; Han, X. Preparation of derivatives of fatty acids. Lipid Anal. 2012, 145–158. [Google Scholar] [CrossRef]
- Collins, S.A.; Xie, S.; Hall, J.R.; White, M.B.; Rise, M.L.; Anderson, D.M. Evaluation of enzyme- and Rhizopus oligosporus -treated high oil residue camelina meal on rainbow trout growth performance and distal intestine histology and inflammatory biomarker gene expression. Aquaculture 2018, 483, 27–37. [Google Scholar] [CrossRef]
- Xue, X.; Hixson, S.M.; Hori, T.S.; Booman, M.; Parrish, C.C.; Anderson, D.M.; Rise, M.L. Atlantic salmon (Salmo salar) liver transcriptome response to diets containing Camelina sativa products. Comp. Biochem. Physiol. Part D Genom. Proteom. 2015, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Katan, T.; Caballero-Solares, A.; Taylor, R.G.; Rise, M.L.; Parrish, C.C. Effect of plant-based diets with varying ratios of ω6 to ω3 fatty acids on growth performance, tissue composition, fatty acid biosynthesis and lipid-related gene expression in Atlantic salmon (Salmo salar). Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 30, 290–304. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Solares, A.; Xue, X.; Parrish, C.C.; Foroutani, M.B.; Taylor, R.G.; Rise, M.L. Changes in the liver transcriptome of farmed Atlantic salmon (Salmo salar) fed experimental diets based on terrestrial alternatives to fish meal and fish oil. BMC Genom. 2018, 19, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, T.-O.; Herschman, H.R. Two inducible, functional cyclooxygenase-2 genes are present in the rainbow trout genome. J. Cell. Biochem. 2007, 102, 1486–1492. [Google Scholar] [CrossRef] [PubMed]
- Morash, A.J.; Bureau, D.P.; McClelland, G.B. Effects of dietary fatty acid composition on the regulation of carnitine palmito-yltransferase (CPT) I in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. 2009, 152, 85–93. [Google Scholar] [CrossRef]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinform. 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kõressaar, K.; Lepamets, M.; Kaplinski, L.; Raime, K.; Andreson, R.; Remm, M. Primer3_masker: Integrating masking of tem-plate sequence with primer design software. Bioinformatics. 2018, 34, 1937–1938. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, J.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruohonen, K. Individual measurements and nested designs in aquaculture experiments: A simulation study. Aquaculture 1998, 165, 149–157. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; Lanois, A.J.; Livesey, E.D.; Bernhard, K.P.; Coley, M.L. Towards Sustainable Aquafeeds: Complete Substitution of Fish Oil with Marine Microalga Schizochytrium sp. Improves Growth and Fatty Acid Deposition in Juvenile Nile Tilapia (Oreochromis niloticus). PLoS ONE 2016, 11, e0156684. [Google Scholar] [CrossRef] [Green Version]
- Roy, J.; Mercier, Y.; Tonnet, L.; Burel, C.; Lanuque, A.; Surget, A.; Larroquet, L.; Corraze, G.; Terrier, F.; Panserat, S.; et al. Rainbow trout prefer diets rich in omega-3 long chain polyunsaturated fatty acids DHA and EPA. Physiol. Behav. 2020, 213, 112692. [Google Scholar] [CrossRef] [PubMed]
- Soengas, J.L. Integration of Nutrient Sensing in Fish Hypothalamus. Front. Neurosci. 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Kousoulaki, K.; Mørkøre, T.; Nengas, I.; Berge, R.; Sweetman, J. Microalgae and organic minerals enhance lipid retention efficiency and fillet quality in Atlantic salmon (Salmo salar L.). Aquaculture 2016, 451, 47–57. [Google Scholar] [CrossRef]
- Bélanger, A.; Sarker, P.; Bureau, D.; Chouinard, Y.; Vandenberg, G. Apparent Digestibility of Macronutrients and Fatty Acids from Microalgae (Schizochytrium sp.) Fed to Rainbow Trout (Oncorhynchus mykiss): A Potential Candidate for Fish Oil Substitution. Animals 2021, 11, 456. [Google Scholar] [CrossRef] [PubMed]
- Jordal, A.-E.O.; Torstensen, B.E.; Tsoi, S.; Tocher, D.R.; Lall, S.P.; Douglas, S.E. Dietary Rapeseed Oil Affects the Expression of Genes Involved in Hepatic Lipid Metabolism in Atlantic Salmon (Salmo salar L.). J. Nutr. 2005, 135, 2355–2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchini, G.M.; Francis, D.S.; Senadheera, S.P.S.D.; Thanuthong, T.; De Silva, S.S. Fish oil replacement with different vegetable oils in Murray cod: Evidence of an “omega-3 sparing effect” by other dietary fatty acids. Aquaculture 2011, 315, 250–259. [Google Scholar] [CrossRef]
- Colombo, S.M.; Parrish, C.C.; Wijekoon, M.P.A. Optimizing long chain-polyunsaturated fatty acid synthesis in salmonids by balancing dietary inputs. PLOS ONE 2018, 13, e0205347. [Google Scholar] [CrossRef]
- Bell, J.; Ashton, I.; Secombes, C.; Weitzel, B.; Dick, J.; Sargent, J. Dietary lipid affects phospholipid fatty acid compositions, eicosanoid production and immune function in Atlantic salmon (Salmo salar). Prostaglandins, Leukot. Essent. Fat. Acids 1996, 54, 173–182. [Google Scholar] [CrossRef]
- Betiku, O.; Barrows, F.; Ross, C.; Sealey, W. The effect of total replacement of fish oil with DHA-Gold® and plant oils on growth and fillet quality of rainbow trout (Oncorhynchus mykiss) fed a plant-based diet. Aquac. Nutr. 2015, 22, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Bélanger-Lamonde, A.; Sarker, P.K.; Ayotte, P.; Bailey, J.L.; Bureau, D.P.; Chouinard, P.Y.; Dewailly, É.; Leblanc, A.; Weber, J.-P.; Vandenberg, G.W. Algal and Vegetable Oils as Sustainable Fish Oil Substitutes in Rainbow Trout Diets: An Approach to Reduce Contaminant Exposure. J. Food Qual. 2018, 2018, 1–12. [Google Scholar] [CrossRef]
- Hixson, S.M.; Parrish, C.C.; Anderson, D.M. Full substitution of fish oil with camelina (Camelina sativa) oil, with partial substitution of fish meal with camelina meal, in diets for farmed Atlantic salmon (Salmo salar) and its effect on tissue lipids and sensory quality. Food Chem. 2014, 157, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Toyes-Vargas, E.A.; Parrish, C.C.; Viana, M.T.; Carreón-Palau, L.; Magallón-Servín, P.; Magallón-Barajas, F.J. Replacement of fish oil with camelina (Camelina sativa) oil in diets for juvenile tilapia (var. GIFT Oreochromis niloticus) and its effect on growth, feed utilization and muscle lipid composition. Aquaculture 2020, 523, 735177. [Google Scholar] [CrossRef]
- Huyben, D.; Rimoldi, S.; Ceccotti, C.; Montero, D.; Betancor, M.; Iannini, F.; Terova, G. Effect of dietary oil fromCamelina sativaon the growth performance, fillet fatty acid profile and gut microbiome of gilthead Sea bream (Sparus aurata). PeerJ 2020, 8, e10430. [Google Scholar] [CrossRef]
- Calder, P.C. Very long chain omega-3 (n-3) fatty acids and human health. Eur. J. Lipid Sci. Technol. 2014, 116, 1280–1300. [Google Scholar] [CrossRef]
- WHO. Fats and Fatty Acids in Human Nutrition. Report on an Expert Consultation; Food and Nutrition Paper; FAO: Geneva, Switzerland, 2008; p. 3. [Google Scholar]
- GOED (Global Organisation for EPA and DHA), 2014. Global Recommendations for EPA and DHA Intake. Available online: http://issfal.org/GlobalRecommendationsSummary19Nov2014Landscape_-3-.pdf (accessed on 15 April 2021).
- American Heart Association. Fish and Omega-3 Fatty Acids. 2017. Available online: https://www.heart.org/en/healthy-living/healthy-eating/eat-smart/fats/fish-and-omega-3-fatty-acids (accessed on 15 April 2021).
- Bell, J.G.; Henderson, R.J.; Tocher, D.R.; Sargent, J.R. Replacement of dietary fish oil with increasing levels of linseed oil: Modification of flesh fatty acid compositions in Atlantic salmon (Salmo salar) using a fish oil finishing diet. Lipids 2004, 39, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Monroig, Ó.; Zheng, X.; Morais, S.; Leaver, M.J.; Taggart, J.B.; Tocher, D.R. Multiple genes for functional ∆6 fatty acyl de-saturases (Fad) in Atlantic salmon (Salmo salar L.): Gene and cDNA characterization, functional expression, tissue distribution and nutritional regulation. Biochim. Biophys. Acta 2010, 1801, 1072–1081. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.; Larsen, B.K.; Granby, K.; Cunha, S.C.; Monteiro, C.; Fernandes, J.O.; Nunes, M.L.; Marques, A.; Dias, J.; Cunha, I.; et al. Diets supplemented with Saccharina latissima influence the expression of genes related to lipid metabolism and oxidative stress modulating rainbow trout (Oncorhynchus mykiss) fillet composition. Food Chem. Toxicol. 2020, 140, 111332. [Google Scholar] [CrossRef]
- Emery, J.A.; Norambuena, F.; Trushenski, J.; Turchini, G.M. Uncoupling EPA and DHA in Fish Nutrition: Dietary Demand is Limited in Atlantic Salmon and Effectively Met by DHA Alone. Lipids 2016, 51, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Rombenso, A.N.; Trushenski, J.T.; Jirsa, D.; Drawbridge, M. Docosahexaenoic acid (DHA) and arachidonic acid (ARA) are essential to meet LC-PUFA requirements of juvenile California Yellowtail (Seriola dorsalis). Aquaculture 2016, 463, 123–134. [Google Scholar] [CrossRef]
- Trushenski, J.; Schwarz, M.; Bergman, A.; Rombenso, A.; Delbos, B. DHA is essential, EPA appears largely expendable, in meeting the n−3 long-chain polyunsaturated fatty acid requirements of juvenile cobia Rachycentron canadum. Aquaculture 2012, 326-329, 81–89. [Google Scholar] [CrossRef]
- Stubhaug, I.; Lie, Ø.; Torstensen, B.E. b-Oxidation capacity in liver increases during parr-smolt transformation of Atlantic salmon fed vegetable oil and fish oil. J. Fish Biol. 2006, 69, 504–517. [Google Scholar] [CrossRef]
- Tocher, D.R. Metabolism and Functions of Lipids and Fatty Acids in Teleost Fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Vestergren, A.S.; Trattner, S.; Pan, J.; Johnsson, P.; Kamal-Eldin, A.; Brännäs, E.; Moazzami, A.A.; Pickova, J. The effect of combining linseed oil and sesamin on the fatty acid composition in white muscle and on expression of lipid-related genes in white muscle and liver of rainbow trout (Oncorhynchus mykiss). Aquac. Int. 2013, 21, 843–859. [Google Scholar] [CrossRef]
- Figueiredo-Silva, A.C.; Kaushik, S.; Terrier, F.; Schrama, J.W.; Médale, F.; Geurden, I. Link between lipid metabolism and voluntary food intake in rainbow trout fed coconut oil rich in medium-chain TAG. Br. J. Nutr. 2011, 107, 1714–1725. [Google Scholar] [CrossRef] [Green Version]
- Sissener, N.; Araujo, P.; Sæle, Ø.; Rosenlund, G.; Stubhaug, I.; Sanden, M. Dietary 18:2n-6 affects EPA (20:5n-3) and ARA (20:4n-6) content in cell membranes and eicosanoid production in Atlantic salmon (Salmo salar L.). Aquaculture 2020, 522, 735098. [Google Scholar] [CrossRef]
- Orr, S.K.; Palumbo, S.; Bosetti, F.; Mount, H.T.; Kang, J.X.; Greenwood, C.E.; Ma, D.W.L.; Serhan, C.N.; Bazinet, R.P. Unesterified docosahexaenoic acid is protective in neuroinflammation. J. Neurochem. 2013, 127, 378–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katan, T.; Xue, X.; Caballero-Solares, A.; Taylor, R.G.; Rise, M.L.; Parrish, C.C. Influence of Dietary Long-Chain Polyunsaturated Fatty Acids and ω6 to ω3 Ratios on Head Kidney Lipid Composition and Expression of Fatty Acid and Eicosanoid Metabolism Genes in Atlantic Salmon (Salmo salar). Front. Mol. Biosci. 2020, 7. [Google Scholar] [CrossRef]
- Arts, M.T.; Kohler, C.C. Health and condition in fish: The influence of lipids on membrane competency and immune response. In Lipids in Aquatic Ecosystems; J.B. Metzler, 2009; pp. 237–256.
- Salini, M.J.; Wade, N.M.; Araújo, B.C.; Turchini, G.M.; Glencross, B.D. Eicosapentaenoic Acid, Arachidonic Acid and Eicosanoid Metabolism in Juvenile Barramundi Lates calcarifer. Lipids 2016, 51, 973–988. [Google Scholar] [CrossRef]
- Caballero-Solares, A.; Hall, J.R.; Xue, X.; Eslamloo, K.; Taylor, R.G.; Parrish, C.C.; Rise, M.L. The dietary replacement of marine ingredients by terrestrial animal and plant alternatives modulates the antiviral immune response of Atlantic salmon (Salmo salar). Fish Shellfish. Immunol. 2017, 64, 24–38. [Google Scholar] [CrossRef]
- Caballero-Solares, A.; Xue, X.; Cleveland, B.M.; Foroutani, M.B.; Parrish, C.C.; Taylor, R.G.; Rise, M.L. Diet-Induced Physiological Responses in the Liver of Atlantic Salmon (Salmo salar) Inferred Using Multiplex PCR Platforms. Mar. Biotechnol. 2020, 22, 511–525. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| FA | MO 1 (%) | MO (µg/mg) |
|---|---|---|
| 14:0 | 13.1 | 193.0 |
| 15:0 | 1.5 | 21.9 |
| 16:0 | 30.7 | 453.5 |
| 16:1n-7c | 4.5 | 66.5 |
| 16:1n-7t | <0.1 | 0.3 |
| 17:0 | 0.3 | 4.2 |
| 17:1n-7 | <0.1 | 0.2 |
| 18:0 | 0.9 | 13.6 |
| 18:1n-9c | 0.6 | 9.0 |
| 18:1n-9t | <0.1 | 0.0 |
| 18:1n-12 | <0.1 | 0.0 |
| 18:1n-7c | 2.8 | 41.0 |
| 18:1n-7t | <0.1 | 0.0 |
| 19:0 | <0.1 | 0.2 |
| 18:2n-6 (LNA) 2 | 0.3 | 4.4 |
| 20:0 | 0.1 | 0.8 |
| 18:3n-6 | 0.1 | 1.6 |
| 20:1n-15 | <0.1 | 0.0 |
| 20:1 | <0.1 | 0.0 |
| 20:1n-9 | <0.1 | 0.3 |
| 18:3n-3 (ALA) 3 | 0.1 | 1.2 |
| 18:2n-6t | 0.1 | 0.8 |
| 18:4n-3 | 0.2 | 2.6 |
| 20:2n-6 | 0.0 | 0.0 |
| 22:3n-3 | 0.1 | 0.8 |
| 22:0 | <0.1 | 0.3 |
| 20:3n-6 | 0.1 | 1.1 |
| 22:1n-9 | <0.1 | 0.5 |
| 20:3n-3 | <0.1 | 0.0 |
| 20:4n-6 (ARA) 4 | 0.1 | 1.8 |
| 22:2n-6 | <0.1 | 0.0 |
| 24:0 | 0.1 | 1.6 |
| 20:5n-3 (EPA) 5 | 0.7 | 10.5 |
| 24:1n-9 | <0.1 | 0.0 |
| 22:4n-6 | <0.1 | 0.2 |
| 22:5n-3 | 0.1 | 1.8 |
| 22:6n-3 (DHA) 6 | 43.4 | 640.7 |
| ΣSFA 7 | 46.7 | 689.1 |
| ΣMUFA 8 | 8.2 | 120.6 |
| ΣPUFA 9 | 45.2 | 666.7 |
| ΣMUFA ≥ 18C | 3.5 | 51.6 |
| ΣMUFA > 18C | 0.1 | 1.6 |
| ΣC18 PUFA | 0.7 | 10.5 |
| ΣC20 PUFA | 0.9 | 13.4 |
| ΣC22 PUFA | 43.6 | 642.8 |
| ΣEPA & DHA | 0.2 | 651.2 |
| Σn-6 | 0.7 | 9.9 |
| Σn-3 | 44.5 | 656.8 |
| ΣOdd chain | 1.9 | 28.0 |
| n-3/n-6 | 67.0 | 67.0 |
| Ingredient 1 (g/kg) | FO | MO/CO | MO |
|---|---|---|---|
| Fish meal | 150 | 150 | 150 |
| Fish (herring) oil | 100 | 0 | 0 |
| Microbial oil (MO) 2 | 0 | 75 | 100 |
| Camelina oil 3 | 50 | 75 | 50 |
| Ground wheat | 170 | 170 | 170 |
| Empyreal (corn protein concentrate) | 120 | 120 | 120 |
| Poultry by-product meal | 210 | 210 | 210 |
| Blood meal | 160 | 160 | 160 |
| Vitamin and mineral mix 4 | 2 | 2 | 2 |
| Dicalcium phosphate | 20 | 20 | 20 |
| Pigment mix 5 | 2.5 | 2.5 | 2.5 |
| Lysine HCl | 5 | 5 | 5 |
| Choline chloride | 10.5 | 10.5 | 10.5 |
| Chemical composition (as fed, g/kg) | |||
| Dry matter | 94.7 | 95.1 | 94.9 |
| Crude protein | 44.2 | 44.5 | 44.6 |
| Total lipid | 23.9 | 24.1 | 24.2 |
| FA | FO | MO/CO | MO | F-Value | p-Value |
|---|---|---|---|---|---|
| 14:0 | 10.8 ± 0.5 b (4.3) | 12.3 ± 0.9 b (5.0) | 18.3 ± 1.6 a (6.8) | 40.25 | 0.000 |
| 16:0 | 45.7 ± 3.0 b (18.1) | 49.3 ± 5.0 b (20.1) | 61.3 ± 4.2 a (22.8) | 11.77 | 0.008 |
| 16:1n-7 | 13.2 ± 0.5 a (5.2) | 7.8 ± 0.7 c (3.2) | 9.5 ± 0.8 b (3.5) | 49.08 | 0.000 |
| 18:0 | 9.8 ± 0.5 a (3.9) | 6.8 ± 0.6 b (2.8) | 6.6 ± 0.5 b (2.5) | 30.76 | 0.001 |
| 18:1n-9 | 41.3 ± 1.4 a (16.4) | 36.0 ± 3.5 ab (14.7) | 31.5 ± 2.6 b (11.7) | 10.28 | 0.012 |
| 18:2n-6 (LNA) 2 | 27.3 ± 0.9 (10.8) | 31.9 ± 2.8 (13.1) | 29.0 ± 2.4 (10.9) | 3.33 | 0.107 |
| 18:3n-3 (ALA) 3 | 25.0 ± 0.8 b (9.9) | 31.1 ± 1.5 a (12.8) | 25.1 ± 2.5 b (9.4) | 11.74 | 0.008 |
| 20:1n-9 | 13.0 ± 0.3 b (5.2) | 15.6 ± 1.1 a (6.4) | 12.6 ± 1.1 b (4.7) | 9.61 | 0.013 |
| 20:4n-6 (ARA) 4 | 1.3 ± 0.0 a (0.5) | 0.6 ± 0.0 b (0.2) | 0.7 ± 0.1 b (0.3) | 190.94 | 0.000 |
| 20:5n-3 (EPA) 5 | 19.0 ± 0.8 a (7.5) | 1.8 ± 0.1 b (0.8) | 2.1 ± 0.2 b (0.8) | 1229.24 | 0.000 |
| 22:1n-9 | 2.2 ± 0.0 b (0.9) | 2.8 ± 0.2 a (1.1) | 2.3 ± 0.2 b (0.8) | 9.83 | 0.013 |
| 22:5n-3 | 2.2 ± 0.1 a (0.9) | 0.3 ± 0.0 b (0.1) | 0.4 ± 0.0 b (0.1) | 1950.49 | 0.000 |
| 22:6n-3 (DHA) 6 | 13.2 ± 0.2 c (5.2) | 28.6 ± 0.8 b (11.7) | 48.9 ± 4.9 a (18.2) | 118.78 | 0.000 |
| 24:1n-9 | 1.1 ± 0.0 a (0.4) | 0.8 ± 0.1 b (0.3) | 0.7 ± 0.1 c (0.2) | 34.65 | 0.001 |
| ∑SFA 7 | 70.1 ± 4.0 b (27.8) | 72.3 ± 6.7 b (29.5) | 90.8 ± 6.7 a (33.8) | 10.95 | 0.010 |
| ∑MUFA 8 | 86.3 ± 2.4 a (34.3) | 73.6 ± 6.5 ab (30.1) | 68.0 ± 5.9 b (25.3) | 9.72 | 0.013 |
| ∑PUFA 9 | 95.3 ± 2.9 (37.9) | 98.3 ± 5.4 (40.3) | 109.9 ± 10.2 (40.9) | 3.80 | 0.086 |
| ∑n-3 | 63.6 ± 1.9 b (25.3) | 63.4 ± 2.5 b (26.0) | 78.0 ± 7.8 a (29.0) | 8.97 | 0.016 |
| ∑n-6 | 31.6 ± 1.0 (12.6) | 34.9 ± 3.0 (14.3) | 31.9 ± 2.6 (11.9) | 1.74 | 0.254 |
| n-3/n-6 | 2.0 ± 0.0 b (2.0) | 1.8 ± 0.1 b (1.8) | 2.4 ± 0.1 a (2.4) | 42.82 | 0.000 |
| Total | 251.7 ± 9.1 | 244.2 ±18.5 | 268.7 ± 22.5 | 1.51 | 0.295 |
| Gene Name (Symbol) (GenBank Acc. No.) a,b,c | Nucleotide sequence (5′–3′) d | Amplicon Size (bp) | Efficiency (%) e | |
|---|---|---|---|---|
| Liver | Muscle | |||
| Acyl-coenzyme A oxidase 1, peroxisomal (acox1) (XM_021568072) | F: ACATACCACTGCCAGGTGTG | 104 | 91.3 | 101.5 |
| R: GCGAGGAATTCGTACGTTCT | ||||
| Arachidonate 5-lipoxygenase a (alox5a) (XM_021580837) | F: GCTGGTGAAGATAGAGAAGCAG | 110 | 81.0 e | 80.4 e |
| R: AGTGGAAGCAGGGGAACTCTA | ||||
| Carnitine O-palmitoyl transferase 1 alpha 1a (cpt1a1a) (AJ619768) | F: CATCCCAGCTGAGTGTCAGA | 100 | 98.8 | 99.0 |
| R: GAAGCAATTGAAGGGGATGA | ||||
| Carnitine O-palmitoyl transferase 1 alpha 1b (cpt1a1b) (GU592679) | F: CCTACTTCAGAAGCGGCAAG | 107 | 90.6 | 99.6 |
| R: CGGGTTGTCGATCTCGTATT | ||||
| Carnitine O-palmitoyl transferase 1 alpha 2a (cpt1a2a) (XM_021624450) | F: CCGCCCATAAAAGACACACT | 123 | ND | 123.9 e |
| R: CAACCTGTTCCCCAGACTGT | ||||
| Carnitine O-palmitoyl transferase 1 alpha 2b (cpt1a2b) (XM_021565275) | F: ACCCCTGATGAGTTTGAACG | 100 | 97.4 | 114.1 e |
| R: CCCAGAGAGCCTTGAGTTTG | ||||
| Carnitine O-palmitoyl transferase 1 beta 1a (cpt1b1a) (AJ606076) | F: TCGCTGTGATAGCCATCATG | 99 | 109.2 e | 105.0 |
| R: TGTAGTCACTGACAGGCAGGG | ||||
| Carnitine O-palmitoyl transferase 1 beta 1b (cpt1b1b) (AF327058) | F: GAATGGTAAACTGGGGGTTAATG | 108 | 92.6 | 94.8 |
| R: GTGTAGCCCAAAAGGAAGCA | ||||
| Cyclooxygenase-1a (cox1a) (AJ299018) | F: TGGGTCTGGGCATGTATCC | 136 | 84.3 | 99.6 |
| R: CAATGCCAAACCTGACACAC | ||||
| Delta 5 fatty acyl desaturase (fadsd5) (XM_021598601) | F: AAATCCGGCTGGAACCACAA | 114 | 92.0 | 105.2 |
| R: AAAAATGTTGGGCTTAGCGTG | ||||
| Delta 6 fatty acyl desaturase a (fadsd6a) (NM_001124287) | F: AGCCATCATTGATGTTGTCG | 155 | 91.7 | 97.2 |
| R: CACAAACGTCTGGGGAAACT d | ||||
| Delta 6 fatty acyl desaturase b (fadsd6b) (XM_021598609) | F: CTACTTATTCCAGTGTATTTCCAC | 94 | 85.0 | 100.0 |
| R: GGTAGAAACTCATCGACCATGC | ||||
| ELOVL fatty acid elongase 2 (elovl2) (KM244737) | F: GATGCCTGCTCTTCCAGTTC | 113 | 91.3 | ND |
| R: GCGACTGGACTTGATGGATT | ||||
| ELOVL fatty acid elongase 5a (elovl5a) (AY605100) | F: CTATGTCATCACACTTATTGCCC | 123 | 92.0 | 91.5 |
| R: ACATGGCCATTCAATGAAGC | ||||
| ELOVL fatty acid elongase 5b (elovl5b) (XM_021576494) | F: ACAAGGCCAGCTGATTCAATT | 123 | 89.1 | ND |
| R: GCAATAAGCGAGGCCATATAG | ||||
| 60S ribosomal protein L32 (rpl32) (CF752566) a | F: AGACCAAGCACATGCTACCC | 149 | 83.1 | 89.0 |
| R: CCTCTCCACAATCAGCTTCC | ||||
| ATP-binding cassette sub-family F member 2 (abcf2) (CA383423) a | F: CGTGTGGTGGATGACAAGAC | 150 | 101.6 | 98.2 |
| R: GTCCAGGTCAATGCCAAACT | ||||
| Beta-actin (actb) (AF157514) a,b,c | F: AGAGCTACGAGCTGCCTGAC | 104 | 87.1 | 96.0 |
| R: GCAAGACTCCATACCGAGGA | ||||
| Elongation factor 1-alpha, oocyte form (ef1a) (CF752140) a,b | F: CTTTGTGCCCATCTCTGGAT | 122 | 82.4 | 94.0 |
| R: CCAGCAGAGTCACACCATTG | ||||
| Eukaryotic translation initiation factor 3 subunit D (etif3d) (CU070663) a | F: CACCGAGCTGAAGAACAACA | 108 | 103.5 | 98.1 |
| R: TTCGCGTGATAACGAGACAC | ||||
| Polyadenylate-binding protein cytoplasmic 1 (pabpc1) (CA355003) a,c | F: AACCGCGCTGCCTACTACT | 102 | 94.3 | 88.6 |
| R: GGGCATGTTCTGGAAGTGTT | ||||
| Growth Parameters | FO | MO/CO | MO | p-Value |
|---|---|---|---|---|
| Initial weight 2 | 19.2 ± 2.9 | 18.7 ± 3.1 | 18.7 ± 2.8 | 0.419 |
| Final weight 3 | 82.1 ± 14.4 | 75.6 ± 17.3 | 80.3 ± 15.8 | 0.064 |
| Weight gain 4 | 62.7 ± 4.6 | 56.9 ± 2.0 | 61.6 ± 2.2 | 0.135 |
| Initial length 2 | 11.7 ± 0.7 | 11.8 ± 0.8 | 11.7 ± 0.6 | 0.356 |
| Final length 3 | 18.5 ± 1.0 a | 17.9 ± 1.2 b | 18.3 ± 1.2 a,b | 0.009 |
| Initial CF 5 | 1.18 ± 0.1 a | 1.14 ± 0.1 b | 1.17 ± 0.1 a,b | 0.011 |
| Final CF 5 | 1.3 ± 0.1 | 1.3 ± 0.1 | 1.3 ± 0.1 | 0.623 |
| Initial VSI 6 | 12.3 ± 1.6 | 13.0 ± 1.1 | 12.9 ± 1.7 | 0.552 |
| Final VSI 6 | 11.6 ± 1.3 | 11.2 ± 1.1 | 10.7 ± 1.4 | 0.169 |
| SGR 7 | 2.6 ± 0.4 | 2.9 ± 0.3 | 2.5 ± 0.2 | 0.225 |
| AFI 8 | 48.1 ± 2.5 | 43.5 ± 3.5 | 50.1 ± 3.6 | 0.107 |
| FCR 9 | 0.8 ± 0.0 | 0.8 ± 0.0 | 0.8 ± 0.0 | 0.467 |
| Whole Body Analysis | ||||
| Crude protein (%) | 52.5 ± 2.1 | 51.7 ± 2.2 | 51.9 ± 2.0 | 0.897 |
| Total fat (%) | 32.5 ± 2.6 | 33.4 ± 3.2 | 31.9 ± 1.7 | 0.773 |
| Ash (%) | 7.2 ± 0.7 | 6.9 ± 0.4 | 6.9 ± 0.5 | 0.700 |
| Calcium (%) | 1.43 ± 0.18 | 1.37 ± 0.11 | 1.43 ± 0.16 | 0.848 |
| Potassium (%) | 1.10 ± 0.04 | 1.07 ± 0.05 | 1.05 ± 0.05 | 0.483 |
| Magnesium (%) | 0.09 ± 0.01 | 0.09 ± 0.01 | 0.09 ± 0.01 | 0.688 |
| Phosphorus (%) | 1.38 ± 0.11 | 1.30 ± 0.08 | 1.34 ± 0.07 | 0.598 |
| Sodium (%) | 0.27 ± 0.01 | 0.25 ± 0.03 | 0.22 ± 0.05 | 0.351 |
| Zinc (ppm) | 67.3 ± 7.1 | 61.8 ± 3.2 | 62.5 ± 5.3 | 0.462 |
| Fatty Acid | Initial | FO | MO/CO | MO | F-Value | p-Value | |||
|---|---|---|---|---|---|---|---|---|---|
| 14:0 | 2.6 ± 0.7 | 1.3 ± 0.2 b | 1.8 ± 0.3 a | 1.8 ± 0.5 a | 8.87 | 0.001 | |||
| 14:1n-5 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.38 | 0.683 | |||
| 15:0 | 0.3 ± 0.1 | 0.2 ± 0.0 b | 0.4 ± 0.1 a | 0.3 ± 0.1 a | 39.99 | 0.000 | |||
| 16:0 | 31.0 ± 5.6 | 20.3 ± 3.4 b | 26.0 ± 4.3 a | 21.9 ± 3.2 b | 9.94 | 0.000 | |||
| 16:1n-7c | 4.4 ± 2.4 | 1.7 ± 0.4 | 1.6 ± 0.6 | 1.7 ± 0.5 | 0.26 | 0.774 | |||
| 16:1n-7t | 0.1 ± 0.0 | 0.1 ± 0.0 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 9.56 | 0.000 | |||
| 17:0 | 0.4 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.0 | 0.2 ± 0.0 | 3.13 | 0.054 | |||
| 17:1n-7 | 0.3 ± 0.1 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | 2.24 | 0.119 | |||
| 18:0 | 10.0 ± 1.7 | 8.0 ± 1.3 a,b | 8.8 ± 1.3 a | 7.3 ± 1.9 b | 3.52 | 0.039 | |||
| 18:1n-9c | 18.8 ± 9.0 | 10.7 ± 2.4 ab | 11.8 ± 1.9 a | 9.6 ± 1.7 b | 4.12 | 0.023 | |||
| 18:1n-9t | 0.1 ± 0.0 | 0.1 ± 0.0 a | 0.1 ± 0.0 b | 0.1 ± 0.0 b | 17.71 | 0.000 | |||
| 18:1n-12 | 0.3 ± 0.1 | 3.0 ± 0.1 a | 0.3 ± 0.1 a | 0.2 ± 0.0 b | 10.48 | 0.000 | |||
| 18:1n-7c | 4.0 ± 1.0 | 1.8 ± 0.4 | 1.9 ± 0.4 | 2.0 ± 0.3 | 1.14 | 0.329 | |||
| 19:0 | 0.2 ± 0.0 | 0.1 ± 0.0 a | 0.1 ± 0.0 b | 0.1 ± 0.0 b | 37.67 | 0.000 | |||
| 19:1n-12 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.02 | 0.982 | |||
| 18:2n-6 (LNA) 2 | 3.4± 1.0 | 3.4 ± 0.5 b | 4.5 ± 0.6a | 3.0 ± 0.6 b | 27.33 | 0.000 | |||
| 20:0 | 0.2 ± 0.1 | 0.3 ± 0.0 b | 0.3 ± 0.0 a | 0.2 ± 0.1c | 23.95 | 0.000 | |||
| 18:3n-6 | 0.1 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 1.45 | 0.245 | |||
| 20:1n-15 | 0.1 ± 0.0 | 0.0 ± 0.0 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 78.17 | 0.000 | |||
| 20:1 | 0.8 ± 0.3 | 0.3 ± 0.0 a | 0.1 ± 0.0 b | 0.1 ± 0.0 b | 124.72 | 0.000 | |||
| 20:1n-9 | 2.2 ± 0.7 | 3.1 ± 0.6 b | 3.8 ± 0.7 a | 2.6 ± 0.9 b | 9.88 | 0.000 | |||
| 18:3n-3 (ALA) 3 | 0.5 ± 0.2 | 1.6 ± 0.3 b | 2.6 ± 0.6 a | 1.5 ± 0.3 b | 34.95 | 0.000 | |||
| 18:4n-3 | 0.2 ± 0.1 | 0.1 ± 0.0 a | 0.1 ± 0.0 a | 0.1 ± 0.0 b | 13.12 | 0.000 | |||
| 20:2n-6 | 1.0 ± 0.3 | 1.7 ± 0.3 b | 2.1 ± 0.4 a | 1.3 ± 0.4 c | 17.72 | 0.000 | |||
| 22:0 | 0.1 ± 0.1 | 0.1 ± 0.0 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 9.90 | 0.000 | |||
| 20:3n-6 | 0.9 ± 0.2 | 0.8 ± 0.1 a | 0.8 ± 0.2 a | 0.5 ± 0.2 b | 14.69 | 0.000 | |||
| 22:1n-11 | 0.3 ± 0.2 | 0.2 ± 0.1 | 0.1 ± 0.0 | 0.1 ± 0.1 | 1.02 | 0.371 | |||
| 22:1n-9 | 0.1 ± 0.1 | 0.2 ± 0.1 a,b | 0.2 ± 0.0 a | 0.1 ± 0.0 b | 8.36 | 0.001 | |||
| 20:3n-3 | 0.1 ± 0.1 | 0.7 ± 0.1 b | 1.1 ± 0.3 a | 0.6 ± 0.2 b | 23.08 | 0.000 | |||
| 20:4n-6 (ARA) 4 | 6.2 ± 1.1 | 4.2 ± 0.7 b | 5.7 ± 0.9 a | 4.8 ± 0.9 b | 11.31 | 0.000 | |||
| 20:5n-3 (EPA) 5 | 9.3 ± 1.5 | 6.2 ± 0.9 a | 2.4 ± 0.5 b | 2.0 ± 0.3 b | 191.76 | 0.000 | |||
| 24:1n-9 | 1.7 ± 0.3 | 1.1 ± 0.3 | 1.1 ± 0.4 | 1.0 ± 0.3 | 0.41 | 0.665 | |||
| 22:4n-6 | 0.2 ± 0.1 | 0.1 ± 0.1 | 0.2 ± 0.2 | 0.3 ± 0.3 | 1.00 | 0.375 | |||
| 22:5n-3 | 2.2 ± 0.4 | 1.5 ± 0.2 | 0.5 ± 0.2 | 0.5 ± 0.1 | 203.28 | 0.000 | |||
| 22:6n-3 (DHA) 6 | 66.4 ± 9.1 | 40.7 ± 5.8 b | 53.6 ± 6.9 a | 44.7 ± 8.6 b | 12.71 | 0.000 | |||
| ΣSFA 7 | 44.7 ± 7.5 | 30.5 ± 4.8 b | 37.6 ± 5.1 a | 31.8 ± 4.9 b | 8.92 | 0.001 | |||
| ΣMUFA 8 | 33.3 ± 13.8 | 19.6 ± 3.9 ab | 21.2 ± 3.1 a | 17.8 ± 3.3 b | 3.67 | 0.034 | |||
| ΣMUFA ≥ 18 | 28.5 ± 11.3 | 17.7 ± 3.5 ab | 19.5 ± 2.7 a | 15.9 ± 3.0 b | 4.83 | 0.013 | |||
| ΣMUFA < 18 | 5.3 ± 1.3 | 4.8 ± 0.8 ab | 5.4 ± 1.0 a | 4.0 ± 1.1 b | 7.80 | 0.001 | |||
| ΣPUFA 9 | 90.7 ± 12.2 | 61.2 ± 8.2 b | 73.6 ± 9.6 a | 59.2 ± 10.8 b | 9.85 | 0.000 | |||
| ΣC18 PUFA | 4.2 ± 1.3 | 5.1 ± 0.8 b | 7.2 ± 1.1 a | 4.6 ± 0.8 b | 33.90 | 0.000 | |||
| ΣC20 PUFA | 17.5 ± 2.6 | 13.7 ± 1.8 | 12.1 ± 2.0 | 9.1 ± 1.8 | 22.47 | 0.000 | |||
| ΣC22 PUFA | 68.9 ± 9.4 | 42.4 ± 6.0 b | 54.4 ± 7.2 a | 45.4 ± 8.9 b | 10.59 | 0.000 | |||
| ΣEPA & DHA | 75.8 ± 10.3 | 47.0 ± 6.6 b | 50.6 ± 7.3 a | 46.7 ± 8.8 b | 7.20 | 0.002 | |||
| Σn-3 | 78.8 ± 10.6 | 50.9 ± 7.0 b | 60.3 ± 8.0 a | 49.2 ± 9.0 b | 8.17 | 0.001 | |||
| Σn-6 | 11.9 ± 1.9 | 10.4 ± 1.4 b | 13.4 ± 1.9 a | 9.9 ± 2.0 b | 16.73 | 0.000 | |||
| Σodd chain | 1.2 ± 03 | 0.7 ± 0.1 b | 0.8 ± 0.1 a | 0.7 ± 0.1 ab | 6.95 | 0.003 | |||
| n-3/n-6 | 6.7 ± 0.7 | 4.9 ± 0.3 a | 4.5 ± 0.4 b | 5.0 ± 0.4 a | 6.69 | 0.003 | |||
| EPA/ARA | 1.5 ± 0.3 | 1.5 ± 0.1a | 0.4 ± 0.0 b | 0.4 ± 0.1 b | 860.23 | 0.000 | |||
| Total Lipid | |||||||||
| Lipid % ww | 4.7 ± 0.6 | 4.2 ± 1.0 | 3.9 ± 0.5 | 4.2 ± 1.0 | 0.59 | 0.560 | |||
| Lipid % dw | 20.1 ± 2.7 | 17.4 ± 4.0 | 16.2 ± 2.3 | 17.5 ± 3.7 | 0.62 | 0.543 | |||
| Lipid (µg/mg) ww | 39.7 ± 6.9 | 26.9 ± 3.7 b | 32.0 ± 3.7 a | 26.2 ± 4.7 b | 8.93 | 0.001 | |||
| Lipid (µg/mg) dw | 168.7 ± 29.8 | 111.2 ±15.1 b | 132.5 ± 16.9 a | 108.7 ± 18.5b | 8.85 | 0.001 | |||
| Fatty Acid | Initial | FO | MO/CO | MO | F-Value | p-Value |
|---|---|---|---|---|---|---|
| 14:0 | 5.1 ± 1.5 | 7.1 ± 2.3 b | 8.9 ± 3.3 ab | 10.9 ± 2.5 a | 7.35 | 0.002 |
| 14:1n-5 | 0.1 ± 0.0 | 0.1 ± 0.1 b | 0.1 ± 0.0 ab | 0.1 ± 0.0 a | 3.83 | 0.030 |
| 15:0 | 0.4 ± 0.1 | 0.5 ± 0.2 b | 1.0 ± 0.4 a | 1.2 ± 0.3 a | 25.99 | 0.000 |
| 16:0 | 26.6 ± 7.3 | 39.6 ± 12.1 b | 47.7 ± 15.2 ab | 51.8 ± 10.7 a | 3.56 | 0.037 |
| 16:1n-7c | 8.7 ± 2.7 | 11.3 ± 4.0 | 9.7 ± 3.9 | 10.8 ± 2.6 | 0.73 | 0.490 |
| 16:1n-7t | 0.1 ± 0.0 | 0.2 ± 0.1 a | 0.1 ± 0.0 b | 0.0 ± 0.0 b | 33.25 | 0.000 |
| 17:0 | 0.3 ± 0.1 | 0.5 ± 0.2 | 0.4 ± 0.1 | 0.4 ± 0.1 | 1.05 | 0.361 |
| 17:1n-7 | 0.2 ± 0.1 | 0.3 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.0 | 2.10 | 0.135 |
| 18:0 | 5.6 ± 1.6 | 8.4 ± 2.7 | 8.7 ± 2.7 | 8.3 ± 1.6 | 0.11 | 0.894 |
| 18:1n-9c | 31.0 ± 9.7 | 40.1 ± 15.0 | 44.5 ±17.4 | 39.1 ± 9.6 | 0.59 | 0.557 |
| 18:1n-9t | 0.2 ± 0.1 | 0.2 ± 0.1 a | 0.1 ± 0.0 b | 0.1 ± 0.0 b | 9.90 | 0.000 |
| 18:1n-12 | 0.9 ± 0.3 | 0.7 ± 0.2 | 0.8 ± 0.3 | 0.7 ± 0.1 | 0.38 | 0.684 |
| 18:1n-7c | 4.7 ± 1.4 | 5.1 ± 1.8 | 5.8 ± 2.2 | 6.0 ± 1.3 | 1.08 | 0.350 |
| 18:1n-7t | 0.3 ± 0.2 | 0.6 ± 0.3 a | 0.1 ± 0.1 b | 0.0 ± 0.0 b | 71.09 | 0.000 |
| 19:1n-12 | 0.3 ± 0.1 | 0.7 ± 0.3 a | 0.1 ± 0.0 b | 0.0 ± 0.0 b | 83.41 | 0.000 |
| 18:2n-6 (LNA) 2 | 11.6 ± 3.4 | 18.9 ± 6.5 b | 26.4 ± 10.3 a | 21.7 ± 5.3 ab | 3.59 | 0.036 |
| 20:0 | 0.2 ± 0.1 | 0.5 ± 0.2 b | 0.7 ± 0.3 a | 0.5 ± 0.1 b | 4.72 | 0.014 |
| 18:3n-6 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.57 | 0.571 |
| 20:1n-15 | 0.0 ± 0.0 | 0.0 ± 0.0 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 23.89 | 0.000 |
| 20:1 | 2.6 ± 0.8 | 1.2 ± 0.5 a | 1.1 ± 0.4 ab | 0.8 ± 0.2 b | 3.69 | 0.033 |
| 20:1n-9 | 2.2 ± 0.7 | 7.7 ± 2.7 b | 11.7 ± 4.4 a | 8.8 ± 2.2 b | 6.01 | 0.005 |
| 18:3n-3 (ALA) 3 | 2.4 ± 0.7 | 13.8 ± 4.5 b | 23.1 ± 8.7 a | 16.1 ± 4.0 b | 9.31 | 0.000 |
| 18:4n-3 | 1.4 ± 0.4 | 2.2 ± 0.7 a | 1.4 ± 0.5 b | 1.1 ± 0.3 b | 19.14 | 0.000 |
| 20:2n-6 | 0.9 ± 0.3 | 2.2 ± 0.6 ab | 2.7 ± 1.0 a | 2.0 ± 0.4 b | 4.35 | 0.019 |
| 22:3n-3 | 0.0 ± 0.0 | 0.1 ± 0.1 b | 0.2 ± 0.1 a | 0.2 ± 0.1 ab | 7.45 | 0.002 |
| 22:0 | 0.1 ± 0.0 | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.0 | 1.13 | 0.332 |
| 20:3n-6 | 0.5 ± 0.2 | 0.6 ± 0.2 | 0.7 ± 0.2 | 0.6 ± 0.1 | 1.09 | 0.347 |
| 22:1n-11 | 2.3 ± 0.7 | 2.4 ± 0.9 | 2.8 ± 1.1 | 2.6 ± 0.6 | 0.88 | 0.422 |
| 22:1n-9 | 0.4 ± 0.1 | 1.2 ± 0.4 b | 1.8 ± 0.7 a | 1.4 ± 0.3 ab | 5.84 | 0.006 |
| 20:3n-3 | 0.2 ± 0.1 | 1.1 ± 0.3 b | 1.9 ± 0.7 a | 1.3 ± 0.3 a | 11.82 | 0.000 |
| 20:4n-6 (ARA) 4 | 1.3 ± 0.3 | 1.4 ± 0.3 | 1.4 ± 0.3 | 1.5 ± 0.2 | 0.64 | 0.534 |
| 22:2n-6 | 0.1 ± 0.0 | 0.2 ± 0.1 a | 0.2 ± 0.1 ab | 0.1 ± 0.0 b | 4.34 | 0.019 |
| 24:0 | 0.1 ± 0.0 | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.1 ± 0.1 | 1.61 | 0.213 |
| 20:5n-3 (EPA) 5 | 7.4 ± 1.8 | 9.5 ± 2.2 a | 3.4 ± 0.8 b | 3.0 ± 0.5 b | 105.04 | 0.000 |
| 24:1n-9 | 0.5 ± 0.1 | 0.8 ± 0.3 | 1.0 ± 0.3 | 0.9 ± 0.2 | 1.59 | 0.215 |
| 22:4n-6 | 0.2 ± 0.0 | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.78 | 0.467 |
| 22:5n-3 | 2.0 ± 0.5 | 2.6 ± 0.8 a | 1.0 ± 0.3 b | 0.9 ± 0.2 b | 60.54 | 0.000 |
| 22:6n-3 (DHA) 6 | 20.6 ± 4.6 | 23.3 ± 3.5 c | 38.7 ± 8.8 b | 47.1 ± 7.7 a | 43.89 | 0.000 |
| ΣSFA 7 | 38.5 ± 10.6 | 56.9 ± 17.7 a | 67.8 ± 22.1 ab | 73.5 ± 15.3 b | 3.10 | 0.056 |
| ΣMUFA 8 | 54.6 ± 16.6 | 72.7 ± 26.5 | 80.1 ± 30.8 | 72.0 ± 17.1 | 0.46 | 0.632 |
| ΣMUFA ≥ C18 | 45.4 ± 13.8 | 61.0 ± 22.3 | 69.9 ± 26.9 | 60.7 ± 14.4 | 0.87 | 0.426 |
| ΣMUFA < C18 | 8.3 ± 2.5 | 14.2 ± 5.1 | 18.7 ± 7.1 | 14.7 ± 3.5 | 3.06 | 0.057 |
| ΣPUFA 9 | 48.9 ± 12.0 | 76.5 ± 19.3 b | 101.3 ± 31.6 a | 95.9 ± 18.8 ab | 4.44 | 0.018 |
| ΣC18 PUFA | 15.7 ± 4.6 | 35.3 ± 11.7 b | 51.2 ± 19.6 a | 39.2 ± 9.6 ab | 5.00 | 0.011 |
| ΣC20 PUFA | 10.3 ± 2.6 | 14.8 ± 3.4 a | 10.0 ± 2.9 b | 8.3 ± 1.4 b | 22.94 | 0.000 |
| ΣC22 PUFA | 22.9 ± 5.1 | 26.3 ± 4.3 c | 40.1 ± 9.2 b | 48.3 ± 7.9 a | 33.51 | 0.000 |
| ΣEPA & DHA | 28.0 ± 6.3 | 32.8 ± 5.5 c | 42.1 ± 9.5 b | 50.1 ± 8.1 a | 17.97 | 0.000 |
| Σn-3 | 34.0 ± 7.9 | 52.6 ± 11.6 b | 69.5 ± 19.6 a | 69.5 ± 12.7 a | 6.30 | 0.004 |
| Σn-6 | 14.9 ± 4.2 | 23.9 ± 7.7 | 31.8 ± 12.0 | 26.4 ± 6.2 | 3.05 | 0.058 |
| ΣOdd chain | 1.3 ± 0.4 | 2.1 ± 0.7 | 1.8 ± 0.7 | 2.0 ± 0.4 | 0.92 | 0.408 |
| n-3/n-6 | 2.3 ± 0.3 | 2.3 ± 0.3 b | 2.3 ± 0.2 b | 2.7 ± 0.2 a | 13.36 | 0.000 |
| EPA/ARA | 5.9 ± 0.4 | 6.9 ± 0.5 a | 2.1 ± 0.2 c | 2.4 ± 0.2 b | 1039.80 | 0.000 |
| EPA + DHA/100g | 607.0 ± 141 | 799.4 ± 151.6 c | 1007.8 ± 241.2 b | 1252.3 ± 231.0 a | 17.9 | 0.000 |
| Total lipid | ||||||
| Lipid % ww | 3.6 ± 0.7 | 4.6 ± 1.2 | 4.8 ± 1.1 | 5.3 ± 1.2 | 1.34 | 0.272 |
| Lipid % dw | 16.6 ± 3.2 | 19.0 ± 4.5 | 20.2 ± 4.3 | 21.2 ± 4.4 | 0.92 | 0.406 |
| Lipid (µg/mg) ww | 30.8 ± 8.8 | 50.4 ± 16.6 | 59.8 ± 20.9 | 60.4 ± 14.3 | 1.55 | 0.223 |
| Lipid (µg/mg) dw | 141.9 ± 38.8 | 206.1 ± 63.2 | 249.2 ± 84.1 | 241.3 ± 50.8 | 1.73 | 0.189 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osmond, A.T.Y.; Arts, M.T.; Hall, J.R.; Rise, M.L.; Bazinet, R.P.; Armenta, R.E.; Colombo, S.M. Schizochytrium sp. (T18) Oil as a Fish Oil Replacement in Diets for Juvenile Rainbow Trout (Oncorhynchus mykiss): Effects on Growth Performance, Tissue Fatty Acid Content, and Lipid-Related Transcript Expression. Animals 2021, 11, 1185. https://doi.org/10.3390/ani11041185
Osmond ATY, Arts MT, Hall JR, Rise ML, Bazinet RP, Armenta RE, Colombo SM. Schizochytrium sp. (T18) Oil as a Fish Oil Replacement in Diets for Juvenile Rainbow Trout (Oncorhynchus mykiss): Effects on Growth Performance, Tissue Fatty Acid Content, and Lipid-Related Transcript Expression. Animals. 2021; 11(4):1185. https://doi.org/10.3390/ani11041185
Chicago/Turabian StyleOsmond, Angelisa T. Y., Michael T. Arts, Jennifer R. Hall, Matthew L. Rise, Richard P. Bazinet, Roberto E. Armenta, and Stefanie M. Colombo. 2021. "Schizochytrium sp. (T18) Oil as a Fish Oil Replacement in Diets for Juvenile Rainbow Trout (Oncorhynchus mykiss): Effects on Growth Performance, Tissue Fatty Acid Content, and Lipid-Related Transcript Expression" Animals 11, no. 4: 1185. https://doi.org/10.3390/ani11041185