Genotyping of Infectious Laryngotracheitis Virus (ILTV) Isolates from Western Canadian Provinces of Alberta and British Columbia Based on Partial Open Reading Frame (ORF) a and b

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. ILTV Isolates
2.2. ILTV Propagation
2.3. ILTV Propagation in Chicken Embryo Liver Cell
2.4. CAM Inoculation
2.5. DNA Extraction
2.6. PCR and Amplicon Purification
2.7. Phylogenetic Analysis
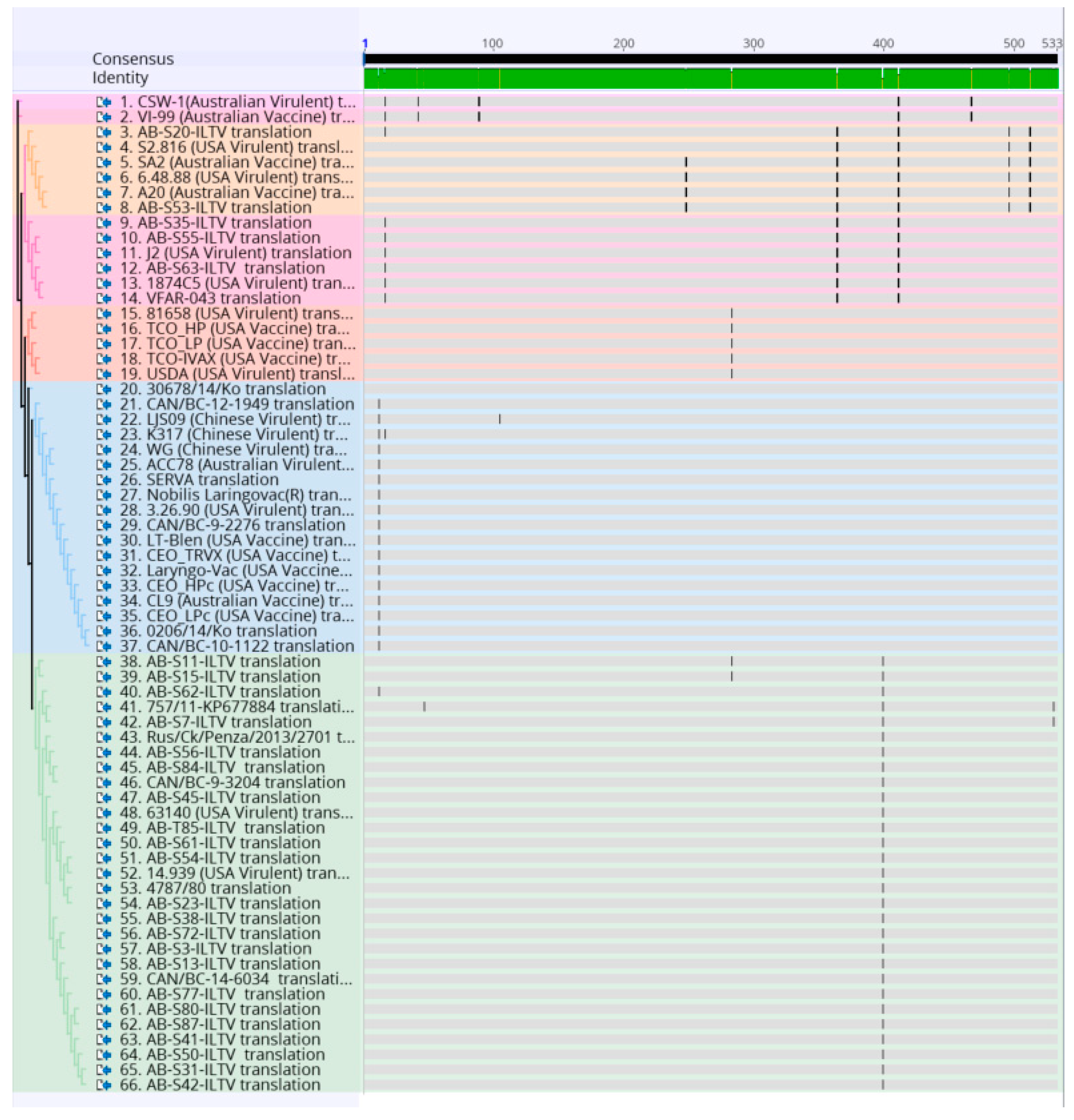
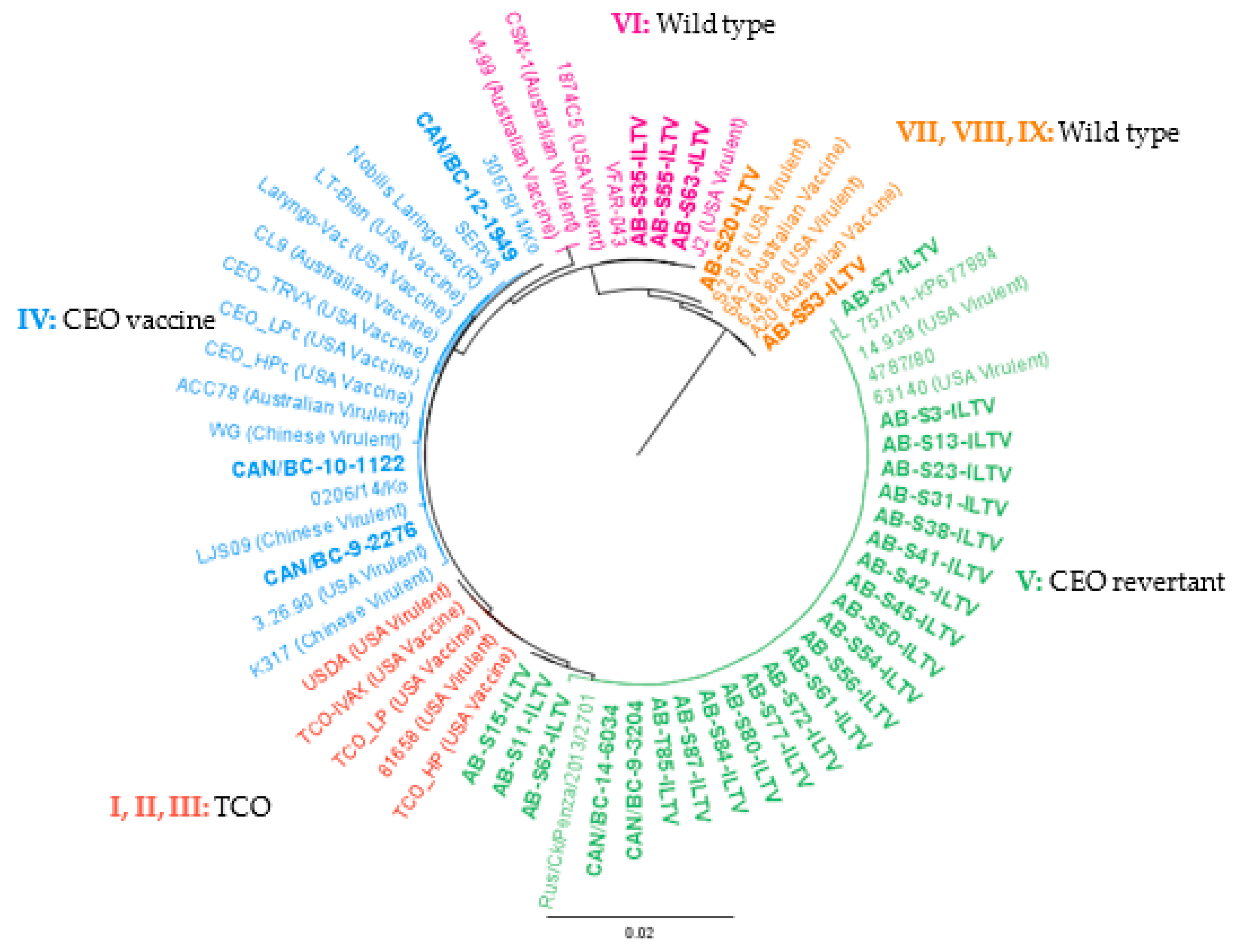
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Garcia, M.; Volkening, J.; Riblet, S.; Spatz, S. Genomic sequence analysis of the United States infectious laryngotracheitis vaccine strains chicken embryo origin (CEO) and tissue culture origin (TCO). Virology 2013, 440, 64–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, H. Infectious laryngotracheitis: A review. Braz. J. Poult. Sci. 2003, 5, 157–168. [Google Scholar] [CrossRef]
- Coppo, M.J.; Hartley, C.A.; Devlin, J.M. Immune responses to infectious laryngotracheitis virus. Dev. Comp. Immunol. 2013, 41, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Kernohan, G. Infectious laryngotracheitis in fowls. J. Am. Vet. Med. Assoc. 1931, 78, 196–202. [Google Scholar]
- Seddon, H.; Hart, L. The ocurrence of infectious laryngotracheitis in fowla in New South Wales. Aust. Vet. J. 1935, 11, 11–221. [Google Scholar] [CrossRef]
- Davison, S.; Smith, G.; Eckroade, R.J. Laryngotracheitis in chickens: The length of the preinfectious and infectious periods. Avian Dis. 1989, 33, 18–23. [Google Scholar] [CrossRef]
- Bagust, T.J. Laryngotracheitis (Gallid-1) herpesvirus infection in the chicken 4. latency establishment by wild and vaccine strains of ILT virus. Avian Pathol. 1986, 15, 581–595. [Google Scholar] [CrossRef]
- Thilakarathne, D.S.; Hartley, C.A.; Diaz-Méndez, A.; Coppo, M.J.C.; Devlin, J.M. Development and application of a combined molecular and tissue culture-based approach to detect latent infectious laryngotracheitis virus (ILTV) in chickens. J. Virol. Methods 2020, 277, 113797. [Google Scholar] [CrossRef]
- Chacon, J.L.; Nunez, L.F.; Vejarano, M.P.; Parra, S.H.; Astolfi-Ferreira, C.S.; Ferreira, A.J. Persistence and spreading of field and vaccine strains of infectious laryngotracheitis virus (ILTV) in vaccinated and unvaccinated geographic regions, in Brazil. Trop Anim. Health Prod. 2015, 47, 1101–1108. [Google Scholar] [CrossRef]
- Han, M.G.; Kim, S.J. Efficacy of live virus vaccines against infectious laryngotracheitis assessed by polymerase chain reaction-restriction fragment length polymorphism. Avian Dis. 2003, 47, 261–271. [Google Scholar] [CrossRef]
- Hughes, C.S.; Williams, R.A.; Gaskell, R.M.; Jordan, F.T.; Bradbury, J.M.; Bennett, M.; Jones, R.C. Latency and reactivation of infectious laryngotracheitis vaccine virus. Arch. Virol. 1991, 121, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.A.; Bennett, M.; Bradbury, J.M.; Gaskell, R.M.; Jones, R.C.; Jordan, F.T.W. Demonstration of sites of latency of infectious laryngotracheitis virus using the polymerase chain reaction. J. Gen. Virol. 1992, 73, 2415–2420. [Google Scholar] [CrossRef]
- Ojkic, D.; Swinton, J.; Vallieres, M.; Martin, E.; Shapiro, J.; Sanei, B.; Binnington, B. Characterization of field isolates of infectious laryngotracheitis virus from Ontario. Avian Pathol. 2006, 35, 286–292. [Google Scholar] [CrossRef] [PubMed]
- MacLachlan, N.J.; Dubovi, E.J. Chapter 9—Herpesvirales. In Fenner’s Veterinary Virology, 4th ed.; MacLachlan, N.J., Dubovi, E.J., Eds.; Academic Press: Boston, MA, USA, 2013; pp. 189–216. [Google Scholar] [CrossRef]
- Dufour-Zavala, L. Epizootiology of infectious laryngotracheitis and presentation of an industry control program. Avian Dis. 2008, 52, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sary, K.; Chénier, S.; Gagnon, C.A.; Shivaprasad, H.L.; Sylvestre, D.; Boulianne, M. Esophagitis and Pharyngitis Associated with Avian Infectious Laryngotracheitis in Backyard Chickens: Two Cases. Avian Dis. 2017, 61, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Gough, A.W.; Pettit, J.R.; Gagnon, A.; Weber, L.J. An outbreak of infectious laryngotracheitis in commercial poultry flocks in Ontario. Can. J. Comp. Med. 1977, 41, 146–151. [Google Scholar]
- Vanderkop, M. Alberta. Infectious laryngotracheitis in a flock of layers. Can. Vet. J. 1990, 31, 780. [Google Scholar]
- Vanderkop, M. Alberta. Infectious laryngotracheitis in commercial broiler chickens. Can. Vet. J. 1993, 34, 185. [Google Scholar]
- UPDATE: Infectious Laryngotracheitis—ILT—Canada. Available online: https://www.canadianpoultrymag.com/disease-watch-infectious-laryngotracheitis-ilt-canada-30562/ (accessed on 21 April 2020).
- ILT—Quebec. Available online: https://www.canadianpoultrymag.com/ilt-quebec-30886/ (accessed on 21 April 2020).
- Garcia, M.; Spatz, S.; Guy, J.S. Infectious Laryngotracheitis. In Disease of Poultry, 13th ed.; Swayne, D.E., Ed.; Wiley-Blackwell: West Sussex, UK, 2013; pp. 161–174. [Google Scholar]
- Shehata, A.A.; Halami, M.Y.; Sultan, H.H.; Abd El-Razik, A.G.; Vahlenkamp, T.W. Chicken embryo origin-like strains are responsible for Infectious laryngotracheitis virus outbreaks in Egyptian cross-bred broiler chickens. Virus Genes 2013, 46, 423–430. [Google Scholar] [CrossRef]
- Spatz, S.J.; Volkening, J.D.; Keeler, C.L.; Kutish, G.F.; Riblet, S.M.; Boettger, C.M.; Clark, K.F.; Zsak, L.; Afonso, C.L.; Mundt, E.S.; et al. Comparative full genome analysis of four infectious laryngotracheitis virus (Gallid herpesvirus-1) virulent isolates from the United States. Virus Genes 2012, 44, 273–285. [Google Scholar] [CrossRef]
- Garcia, M.; Zavala, G. Commercial Vaccines and Vaccination Strategies Against Infectious Laryngotracheitis: What We Have Learned and Knowledge Gaps That Remain. Avian Dis. 2019, 63, 325–334. [Google Scholar] [CrossRef]
- Johnson, D.I.; Vagnozzi, A.; Dorea, F.; Riblet, S.M.; Mundt, A.; Zavala, G.; Garcia, M. Protection Against Infectious Laryngotracheitis by In Ovo Vaccination with Commercially Available Viral Vector Recombinant Vaccines. Avian Dis. 2010, 54, 1251–1259. [Google Scholar] [CrossRef] [PubMed]
- Vagnozzi, A.; Zavala, G.; Riblet, S.M.; Mundt, A.; García, M. Protection induced by commercially available live-attenuated and recombinant viral vector vaccines against infectious laryngotracheitis virus in broiler chickens. Avian Pathol. 2012, 41, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Davison, S.; Eric, N.G.; Susan, C.; Eckroade, R.J. Evaluation of the Efficacy of a Live Fowlpox-Vectored Infectious Laryngotracheitis/Avian Encephalomyelitis Vaccine against ILT Viral Challenge. Avian Dis. 2006, 50, 50–54. [Google Scholar] [CrossRef]
- Esaki, M.; Godoy, A.; Rosenberger, J.K.; Rosenberger, S.C.; Gardin, Y.; Yasuda, A.; Dorsey, K.M. Protection and Antibody Response Caused by Turkey Herpesvirus Vector Newcastle Disease Vaccine. Avian Dis. 2013, 57, 750–755. [Google Scholar] [CrossRef]
- Oldoni, I.; Rodriguez-Avila, A.; Riblet, S.; Garcia, M. Characterization of Infectious Laryngotracheitis Virus (ILTV) Isolates from Commercial Poultry by Polymerase Chain Reaction and Restriction Fragment Length Polymorphism (PCR-RFLP). Avian Dis. 2008, 52, 59–63. [Google Scholar] [CrossRef]
- Andreasen, J.R.; Glisson, J.R.; Villegas, P. Differentiation of Vaccine Strains and Georgia Field Isolates of Infectious Laryngotracheitis Virus by Their Restriction Endonuclease Fragment Patterns. Avian Dis. 1990, 34, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Guy, J.S.; Barnes, H.J.; Munger, L.L.; Rose, L. Restriction Endonuclease Analysis of Infectious Laryngotracheitis Viruses: Comparison of Modified-Live Vaccine Viruses and North Carolina Field Isolates. Avian Dis. 1989, 33, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Keeler, C.L.; Hazel, J.W.; Hastings, J.E.; Rosenberger, J.K. Restriction Endonuclease Analysis of Delmarva Field Isolates of Infectious Laryngotracheitis Virus. Avian Dis. 1993, 37, 418–426. [Google Scholar] [CrossRef]
- Spatz, S.J.; Garcia, M.; Riblet, S.; Ross, T.A.; Volkening, J.D.; Taylor, T.L.; Kim, T.; Afonso, C.L. MinION sequencing to genotype US strains of infectious laryngotracheitis virus. Avian Pathol. 2019, 48, 255–269. [Google Scholar] [CrossRef]
- Fuchs, W.; Veits, J.; Helferich, D.; Granzow, H.; Teifke, J.P.; Mettenleiter, T.C. Molecular biology of avian infectious laryngotracheitis virus. Vet. Res. 2007, 38, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkpatrick, N.C.; Mahmoudian, A.; Colson, C.A.; Devlin, J.M.; Noormohammadi, A.H. Relationship between mortality, clinical signs and tracheal pathology in infectious laryngotracheitis. Avian Pathol. 2006, 35, 449–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.J.; La, T.M.; Choi, I.S.; Song, C.S.; Park, S.Y.; Lee, J.B.; Lee, S.W. Genotyping of infectious laryngotracheitis virus using allelic variations from multiple genomic regions. Avian Pathol. 2016, 45, 443–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, S.M.; Dufour-Zavala, L.; Jackwood, M.W.; Lee, M.D.; Lupiani, B.; Reed, W.M.; Spackman, E.; Woolcock, P.R. A Laboratory Manual for the Isolation, Identification and Charaacterization of Avian Pathognes, 5th ed.; American Association of Avian Pathologists: Athens, GA, USA, 2008. [Google Scholar]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Oldoni, I.; García, M. Characterization of infectious laryngotracheitis virus isolates from the US by polymerase chain reaction and restriction fragment length polymorphism of multiple genome regions. Avian Pathol. 2007, 36, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Bayoumi, M.; El-Saied, M.; Amer, H.; Bastami, M.; Sakr, E.E.; El-Mahdy, M. Molecular characterization and genetic diversity of the infectious laryngotracheitis virus strains circulating in Egypt during the outbreaks of 2018 and 2019. Arch. Virol. 2020, 165, 661–670. [Google Scholar] [CrossRef]
- Craig, M.I.; Rojas, M.F.; van der Ploeg, C.A.; Olivera, V.; Vagnozzi, A.E.; Perez, A.M.; Konig, G.A. Molecular Characterization and Cluster Analysis of Field Isolates of Avian Infectious Laryngotracheitis Virus from Argentina. Front. Vet. Sci. 2017, 4, 212. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Avila, A.; Oldoni, I.; Riblet, S.; Garcia, M. Evaluation of the protection elicited by direct and indirect exposure to live attenuated infectious laryngotracheitis virus vaccines against a recent challenge strain from the United States. Avian Pathol. 2008, 37, 287–292. [Google Scholar] [CrossRef]
- Guy, J.S.; Barnes, H.J.; Smith, L. Increased Virulence of Modified-Live Infectious Laryngotracheitis Vaccine Virus Following Bird-to-Bird Passage. Avian Dis. 1991, 35, 348–355. [Google Scholar] [CrossRef]
- Agnew-Crumpton, R.; Vaz, P.K.; Devlin, J.M.; O’Rourke, D.; Blacker-Smith, H.P.; Konsak-Ilievski, B.; Hartley, C.A.; Noormohammadi, A.H. Spread of the newly emerging infectious laryngotracheitis viruses in Australia. Infect. Genet. Evol. 2016, 43, 67–73. [Google Scholar] [CrossRef]
- La, T.-M.; Choi, E.-J.; Lee, J.-B.; Park, S.-Y.; Song, C.-S.; Choi, I.-S.; Lee, S.-W. Comparative genome analysis of Korean field strains of infectious laryngotracheitis virus. PLoS ONE 2019, 14, e0211158. [Google Scholar] [CrossRef]
- Lee, S.W.; Devlin, J.M.; Markham, J.F.; Noormohammadi, A.H.; Browning, G.F.; Ficorilli, N.P.; Hartley, C.A.; Markham, P.F. Phylogenetic and molecular epidemiological studies reveal evidence of multiple past recombination events between infectious laryngotracheitis viruses. PLoS ONE 2013, 8, e55121. [Google Scholar] [CrossRef] [Green Version]
- Palomino-Tapia, V.A.; Zavala, G.; Cheng, S.; Garcia, M. Long-term protection against a virulent field isolate of infectious laryngotracheitis virus induced by inactivated, recombinant, and modified live virus vaccines in commercial layers. Avian Pathol. 2019, 48, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Chacon, J.L.; Ferreira, A.J. Differentiation of field isolates and vaccine strains of infectious laryngotracheitis virus by DNA sequencing. Vaccine 2009, 27, 6731–6738. [Google Scholar] [CrossRef] [PubMed]
- Veits, J.; Köllner, B.; Teifke, J.P.; Granzow, H.; Mettenleiter, T.C.; Fuchs, W. Isolation and Characterization of Monoclonal Antibodies against Structural Proteins of Infectious Laryngotracheitis Virus. Avian Dis. 2003, 47, 330–342. [Google Scholar] [CrossRef]
- Ziemann, K.; Mettenleiter, T.C.; Fuchs, W. Gene arrangement within the unique long genome region of infectious laryngotracheitis virus is distinct from that of other alphaherpesviruses. J. Virol. 1998, 72, 847–852. [Google Scholar] [CrossRef] [Green Version]
- Veits, J.; Mettenleiter, T.C.; Fuchs, W. Five unique open reading frames of infectious laryngotracheitis virus are expressed during infection but are dispensable for virus replication in cell culture. J. Gen. Virol. 2003, 84, 1415–1425. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Origin | Year | ORF a and b Gene Genotyping | Province | GenBank Accession Number | Vaccinated |
|---|---|---|---|---|---|---|
| CAN/BC-9-2276 | Broiler | 2009 | IV | British Columbia | MT639625 | No |
| CAN/BC-9-3204 | Broiler | 2009 | V | British Columbia | MT639621 | Unknown |
| CAN/BC-10-1122 | Layer | 2010 | IV | British Columbia | MT639626 | Yes |
| CAN/BC-12-1949 | Broiler | 2012 | IV | British Columbia | MT639627 | No |
| AB-S3-ILTV | Backyard chicken | 2013 | V | Alberta | MT639620 | No |
| CAN/BC-14-6034 | Layer | 2014 | V | British Columbia | MT639619 | Unknown |
| AB-S7-ILTV | Backyard chicken | 2014 | V | Alberta | MT639603 | No |
| AB-S11-ILTV | Backyard chicken | 2014 | V | Alberta | MT639601 | No |
| AB-S13-ILTV | Backyard chicken | 2014 | V | Alberta | MT639605 | No |
| AB-S15-ILTV | Backyard chicken | 2014 | V | Alberta | MT639602 | Yes |
| AB-S20-ILTV | Backyard chicken | 2015 | VII, VIII, IX | Alberta | MT639631 | No |
| AB-S23-ILTV | Backyard chicken | 2015 | V | Alberta | MT639606 | No |
| AB-S31-ILTV | Backyard chicken | 2015 | V | Alberta | MT639607 | No |
| AB-S35-ILTV | Backyard chicken | 2015 | VI | Alberta | MT639628 | No |
| AB-S38-ILTV | Backyard chicken | 2015 | V | Alberta | MT639608 | No |
| AB-S41-ILTV | Backyard chicken | 2016 | V | Alberta | MT639609 | No |
| AB-S42-ILTV | Backyard chicken | 2016 | V | Alberta | MT639610 | No |
| AB-S45-ILTV | Backyard chicken | 2016 | V | Alberta | MT639611 | No |
| AB-S50-ILTV | Backyard chicken | 2016 | V | Alberta | MT639613 | No |
| AB-S53-ILTV | Backyard chicken | 2016 | VII, VIII, IX | Alberta | MT639632 | No |
| AB-S54-ILTV | Backyard chicken | 2016 | V | Alberta | MT639622 | No |
| AB-S55-ILTV | Backyard chicken | 2016 | VI | Alberta | MT639629 | No |
| AB-S56-ILTV | Backyard chicken | 2017 | V | Alberta | MT639604 | No |
| AB-S61-ILTV | Backyard chicken | 2017 | V | Alberta | MT639623 | No |
| AB-S62-ILTV | Backyard chicken | 2017 | V | Alberta | MT639624 | No |
| AB-S63-ILTV | Backyard chicken | 2017 | VI | Alberta | MT639630 | No |
| AB-S72-ILTV | Backyard chicken | 2017 | V | Alberta | MT639615 | No |
| AB-S77-ILTV | Backyard chicken | 2017 | V | Alberta | MT639616 | No |
| AB-S80-ILTV | Backyard chicken | 2017 | V | Alberta | MT639617 | No |
| AB-S84-ILTV | Backyard chicken | 2017 | V | Alberta | MT639618 | No |
| AB-S85-ILTV | Backyard chicken | 2018 | V | Alberta | MT639612 | No |
| AB-S87-ILTV | Backyard chicken | 2018 | V | Alberta | MT639614 | No |
| SNP Position | Reference Nucleotide | Nucleotide Change | Type of Nucleotide Change | AA Change | Reference AA | AA Change | Genotype and/or Isolates with Same Change |
|---|---|---|---|---|---|---|---|
| 34 | A | C | Tn ** | 12 | Lys | Gln | All except IV |
| 49 | G | A | Tr *** | 17 | Ala | Thr | K317 |
| 50 | C * | T | Tr | 17 | Ala | Val | VI |
| 126 | T | A | Tn | 42 | Cys | * | VI-99 and CSW-1 |
| 139 | C | T | Tr | 47 | Pro | Ser | 757/11 |
| 265 | G | A | Tr | 89 | Val | Met | VI-99 and CSW-1 |
| 314 | T | C | Tr | 105 | Leu | Ser | LJS09 |
| 742 | A | G | Tr | 248 | Thr | Ala | VII, VIII, IX except AB-S20-ILTV and S2.816 |
| 848 | T | C | Tr | 283 | Val | Ala | I, II, III, and AB-S11-ILTV and AB-S15-ILTV |
| 1091 | T | C | Tr | 364 | Val | Ala | VI, VII, VIII, IX |
| 1196 | A | C | Tn | 399 | Gln | Pro | V |
| 1231 | G | T | Tn | 411 | Gly | Cys | VI, VII, VIII, IX |
| 1399 | G | A | Tr | 467 | Glu | Lys | VI-99 and CSW-1 |
| 1487 | A | G | Tr | 496 | Lys | Arg | VII, VIII, IX |
| 1533 | AG | GA | Tr | 512 | Ala | Thr | VII, VIII, IX |
| 1588 | T | C | Tr | 530 | Trp | Arg | AB-S7-ILTV and 757/11 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barboza-Solis, C.; Contreras, A.P.; Palomino-Tapia, V.A.; Joseph, T.; King, R.; Ravi, M.; Peters, D.; Fonseca, K.; Gagnon, C.A.; van der Meer, F.; et al. Genotyping of Infectious Laryngotracheitis Virus (ILTV) Isolates from Western Canadian Provinces of Alberta and British Columbia Based on Partial Open Reading Frame (ORF) a and b. Animals 2020, 10, 1634. https://doi.org/10.3390/ani10091634
Barboza-Solis C, Contreras AP, Palomino-Tapia VA, Joseph T, King R, Ravi M, Peters D, Fonseca K, Gagnon CA, van der Meer F, et al. Genotyping of Infectious Laryngotracheitis Virus (ILTV) Isolates from Western Canadian Provinces of Alberta and British Columbia Based on Partial Open Reading Frame (ORF) a and b. Animals. 2020; 10(9):1634. https://doi.org/10.3390/ani10091634
Chicago/Turabian StyleBarboza-Solis, Catalina, Ana Perez Contreras, Victor A. Palomino-Tapia, Tomy Joseph, Robin King, Madhu Ravi, Delores Peters, Kevin Fonseca, Carl A. Gagnon, Frank van der Meer, and et al. 2020. "Genotyping of Infectious Laryngotracheitis Virus (ILTV) Isolates from Western Canadian Provinces of Alberta and British Columbia Based on Partial Open Reading Frame (ORF) a and b" Animals 10, no. 9: 1634. https://doi.org/10.3390/ani10091634