Effects of Thymol and Thymol α-D-Glucopyranoside on Intestinal Function and Microbiota of Weaned Pigs

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Experimental Design and Measurements
2.3. Sample Collection
2.4. Laboratory Analysis
2.4.1. Concentrations of Active Compounds in Digesta
2.4.2. Small Intestinal Barrier Function and Histo-Morphology
2.4.3. Bacteriological Analyses
2.4.4. Glycosidase Activity in Mucosa and Digesta
2.4.5. Statistical Analysis
3. Results
3.1. Animal Performance
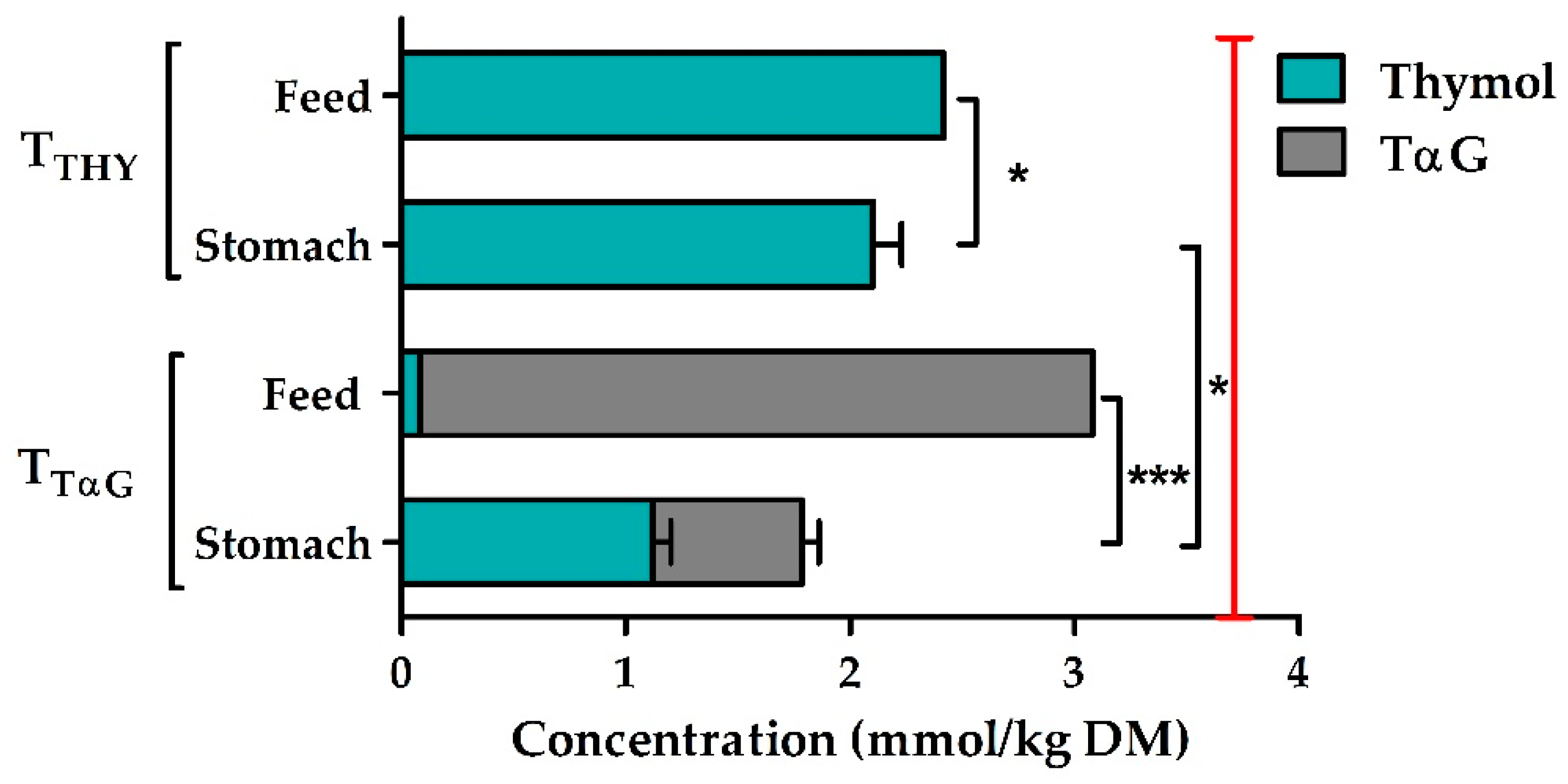
3.2. Concentrations of Active Compounds
3.3. Small Intestinal Barrier Function and Histo-Morphology
3.4. Bacteriological Analyses
3.4.1. Bacterial Counts
3.4.2. Bacterial Metabolites
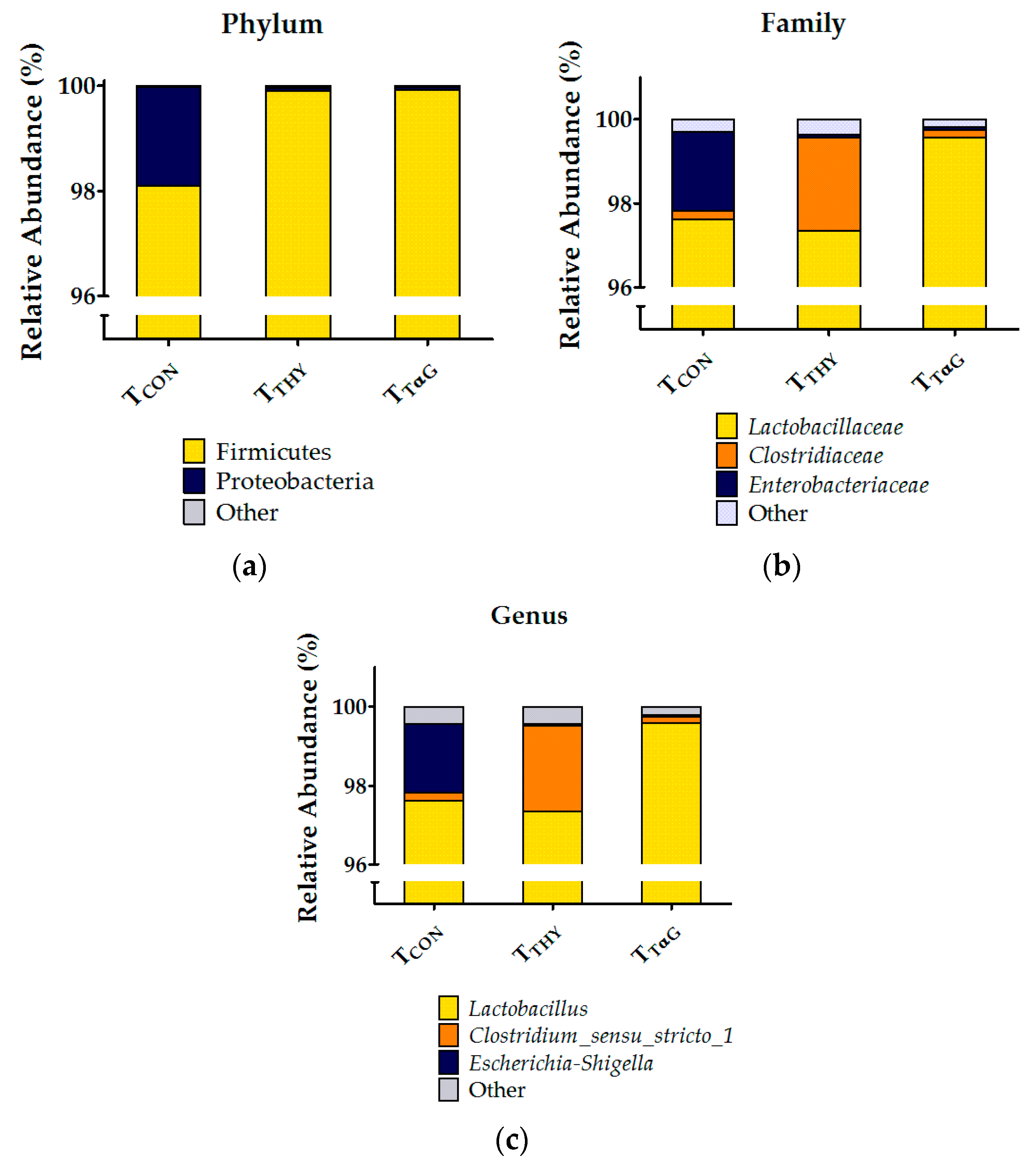
3.4.3. Microbial Composition by 16S rRNA Profiling
3.5. Glycosidase Activity in Mucosa and Digesta
4. Discussion
4.1. Effect of Thymol and TαG Supplementation on Pig Performance
4.2. Effects of TαG Supplementation on Thymol Concentrations in the Proximal GI Tract
4.3. Effects of Thymol and TαG Supplementation on the Intestinal Barrier Function
4.4. Effects of Thymol and TαG Supplementation on Gut Microbiota
4.5. Effects of Thymol and TαG Supplementation on the Glycosidase Activity in Mucosa
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lalles, J.-P.; Bosi, P.; Smidt, H.; Stokes, C.R. Nutritional management of gut health in pigs around weaning. Proc. Nutr. Soc. 2007, 66, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission. Regulation (EC) No 1831/2003 of the European Parliament and of the Council of 22 September 2003 on additives for use in animal nutrition. Off. J. Eur. Union 2003, 268, 29–43. [Google Scholar]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency. Questions and Answers on Veterinary Medicinal Products Containing Zinc Oxide to Be Administered Orally to Food-Producing Species. Outcome of a Referral Procedure under Article 35 of Directive 2001/82/EC (EMEA/V/A/118). Available online: https://www.ema.europa.eu/en/medicines/veterinary/referrals/zinc-oxide (accessed on 15 January 2020).
- Omonijo, F.A.; Ni, L.; Gong, J.; Wang, Q.; Lahaye, L.; Yang, C. Essential oils as alternatives to antibiotics in swine production. Anim. Nutr. 2018, 4, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Amasheh, S.; Aschenbach, J.R. Modulation of gastrointestinal barrier and nutrient transport function in farm animals by natural plant bioactive compounds—A comprehensive review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3237–3266. [Google Scholar] [CrossRef]
- Meeran, N.; Fizur, M.; Javed, H.; Al Taee, H.; Azimullah, S.; Ojha, S.K. Pharmacological properties and molecular mechanisms of thymol: Prospects for its therapeutic potential and pharmaceutical development. Front. Pharmacol. 2017, 8, 380. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Chowdhury, M.A.K.; Huo, Y.; Gong, J. Phytogenic compounds as alternatives to in-feed antibiotics: Potentials and challenges in application. Pathogens 2015, 4, 137–156. [Google Scholar] [CrossRef] [Green Version]
- Michiels, J.; Missotten, J.; Fremaut, D.; De Smet, S.; Dierick, N. In vitro characterisation of the antimicrobial activity of selected essential oil components and binary combinations against the pig gut flora. Anim. Feed Sci. Technol. 2009, 151, 111–127. [Google Scholar] [CrossRef]
- Si, W.; Gong, J.; Tsao, R.; Zhou, T.; Yu, H.; Poppe, C.; Johnson, R.; Du, Z. Antimicrobial activity of essential oils and structurally related synthetic food additives towards selected pathogenic and beneficial gut bacteria. J. Appl. Microbiol. 2006, 100, 296–305. [Google Scholar] [CrossRef]
- Michiels, J.; Missotten, J.; Dierick, N.; Fremaut, D.; Maene, P.; De Smet, S. In vitro degradation and in vivo passage kinetics of carvacrol, thymol, eugenol and trans-cinnamaldehyde along the gastrointestinal tract of piglets. J. Sci. Food Agric. 2008, 88, 2371–2381. [Google Scholar] [CrossRef]
- Jugl-Chizzola, M.; Ungerhofer, E.; Gabler, C.; Hagmüller, W.; Chizzola, R.; Zitterl-Eglseer, K.; Franz, C. Testing of the palatability of Thymus vulgaris L. and Origanum vulgare L. as flavouring feed additive for weaner pigs on the basis of a choice experiment. Berl. Munch. Tierarztl. Wochenschr. 2006, 119, 238–243. [Google Scholar] [PubMed]
- Dušan, F.; Marián, S.; Katarína, D.; Dobroslava, B. Essential oils—Their antimicrobial activity against Escherichia coli and effect on intestinal cell viability. Toxicol. Vitr. 2006, 20, 1435–1445. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.M.; Opapeju, F.O.; Pluske, J.R.; Kim, J.C.; Hampson, D.J.; Nyachoti, C.M. Gastrointestinal health and function in weaned pigs: A review of feeding strategies to control post-weaning diarrhoea without using in-feed antimicrobial compounds. J. Anim. Physiol. Anim. Nutr. 2013, 97, 207–237. [Google Scholar] [CrossRef]
- Jones, P.; Vogt, T. Glycosyltransferases in secondary plant metabolism: Tranquilizers and stimulant controllers. Planta 2001, 213, 164–174. [Google Scholar] [CrossRef]
- Friend, D.R. Colon-specific drug delivery. Adv. Drug Deliv. Rev. 1991, 7, 149–199. [Google Scholar] [CrossRef]
- Petrujkić, B.T.; Sedej, I.; Beier, R.C.; Anderson, R.C.; Harvey, R.B.; Epps, S.V.; Stipanovic, R.D.; Krueger, N.A.; Nisbet, D.J. Ex vivo absorption of thymol and thymol-β-D-glucopyranoside in piglet everted jejunal segments. J. Agric. Food Chem. 2013, 61, 3757–3762. [Google Scholar] [CrossRef]
- Van Noten, N.; Van Liefferinge, E.; Degroote, J.; De Smet, S.; Desmet, T.; Michiels, J. Fate of thymol and its monoglucosides in the gastrointestinal tract of piglets. Omega Under Rev. Under review.
- Nieddu, M.; Rassu, G.; Boatto, G.; Bosi, P.; Trevisi, P.; Giunchedi, P.; Carta, A.; Gavini, E. Improvement of thymol properties by complexation with cyclodextrins: In vitro and in vivo studies. Carbohydr. Polym. 2014, 102, 393–399. [Google Scholar] [CrossRef]
- Rassu, G.; Nieddu, M.; Bosi, P.; Trevisi, P.; Colombo, M.; Priori, D.; Manconi, P.; Giunchedi, P.; Gavini, E.; Boatto, G. Encapsulation and modified-release of thymol from oral microparticles as adjuvant or substitute to current medications. Phytomedicine 2014, 21, 1627–1632. [Google Scholar] [CrossRef]
- Hjelmeland, A.K.; Ebeler, S.E. Glycosidically Bound Volatile Aroma Compounds in Grapes and Wine: A Review. Am. J. Enol. Vitic. 2015, 66, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Uchida, K. Enzymatic glycosylation of aglycones of pharmacological significance. In Carbohydrate Biotechnology Protocols, 1st ed.; Bucke, C., Ed.; Humana Press: Totowa, NJ, USA, 1999; pp. 297–312. ISBN 978-1-59259-261-6. [Google Scholar]
- CVB. Feed Recommandations for Pigs; Centraal Veevoederbureau: Wageningen, The Netherlands, 2016. [Google Scholar]
- Spranghers, T.; Michiels, J.; Vrancx, J.; Ovyn, A.; Eeckhout, M.; De Clercq, P.; De Smet, S. Gut antimicrobial effects and nutritional value of black soldier fly (Hermetia illucens L.) prepupae for weaned piglets. Anim. Feed Sci. Technol. 2018, 235, 33–42. [Google Scholar] [CrossRef]
- De Winter, K.; Dewitte, G.; Dirks-Hofmeister, M.E.; De Laet, S.; Pelantová, H.; Křen, V.; Desmet, T. Enzymatic glycosylation of phenolic antioxidants: Phosphorylase-mediated synthesis and characterization. J. Agric. Food Chem. 2015, 63, 10131–10139. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Degroote, J.; Van Ginneken, C.; Van Poucke, M.; Vergauwen, H.; Dam, T.M.T.; Vanrompay, D.; Peelman, L.J.; De Smet, S.; Michiels, J. Intrauterine growth restriction in neonatal piglets affects small intestinal mucosal permeability and mRNA expression of redox-sensitive genes. FASEB J. 2016, 30, 863–873. [Google Scholar] [CrossRef] [Green Version]
- McKie, A.; Zammit, P.; Naftalin, R. Comparison of cattle and sheep colonic permeabilities to horseradish peroxidase and hamster scrapie prion protein in vitro. Gut 1999, 45, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Van Nevel, C.; Decuypere, J.; Dierick, N.; Molly, K. The influence of Lentinus edodes (Shiitake mushroom) preparations on bacteriological and morphological aspects of the small intestine in piglets. Arch. Anim. Nutr. 2003, 57, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Vanderheyde, H.; Henderickx, H. Zur Vereinfachung der quantitativen und qualitativen Bestimmung der Bakterien unter Verwendung von Ringplatten. Zentralblatt Bakteriol. 1963, 189, 224–228. [Google Scholar]
- Missotten, J.A.M.; Goris, J.; Michiels, J.; Van Coillie, E.; Herman, L.; De Smet, S.; Dierick, N.A.; Heyndrickx, M. Screening of isolated lactic acid bacteria as potential beneficial strains for fermented liquid pig feed production. Anim. Feed Sci. Technol. 2009, 150, 122–138. [Google Scholar] [CrossRef]
- Castro-Montoya, J.; De Campeneere, S.; Van Ranst, G.; Fievez, V. Interactions between methane mitigation additives and basal substrates on in vitro methane and VFA production. Anim. Feed Sci. Technol. 2012, 176, 47–60. [Google Scholar] [CrossRef]
- Conway, E.J. Microdiffusion Analysis and Volumetric Error, 4th ed.; Crosby Lockwood and Son, Ltd.: London, UK, 1957; p. 465. ISBN 978-025-896-635-8. [Google Scholar]
- Cong, O.N.; Bernard, T.; Pham Kim, D.; Do Duc, L.; Nassim, M.; Nguyen Thi, H.; Nguyen Hoang, T.; Georges, D.; Jérôme, B.; Vu Dinh, T. Growth performance, carcass quality characteristics and colonic microbiota profiles in finishing pigs fed diets with different inclusion levels of rice distillers’ by-product. Anim. Sci. J. 2019, 90, 948–960. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Fastrès, A.; Taminiau, B.; Vangrinsven, E.; Tutunaru, A.-C.; Moyse, E.; Farnir, F.; Daube, G.; Clercx, C. Effect of an antimicrobial drug on lung microbiota in healthy dogs. Heliyon 2019, 5, e02802. [Google Scholar] [CrossRef] [PubMed]
- Manzanilla, E.G.; Perez, J.F.; Martin, M.; Kamel, C.; Baucells, F.; Gasa, J. Effect of plant extracts and formic acid on the intestinal equilibrium of early-weaned pigs. J. Anim. Sci. 2004, 82, 3210–3218. [Google Scholar] [CrossRef] [Green Version]
- Hagmüller, W.; Jugl-Chizzola, M.; Zitterl-Eglseer, K.; Gabler, C.; Spergser, J.; Chizzola, R.; Franz, C. The use of Thymi Herba as feed additive (0.1%, 0.5%, 1.0%) in weanling piglets with assessment of the shedding of haemolysing E. coli and the detection of thymol in the blood plasma. Berl. Munch. Tierarztl. Wochenschr. 2006, 119, 50–54. [Google Scholar]
- Wei, H.K.; Xue, H.X.; Zhou, Z.X.; Peng, J. A carvacrol—Thymol blend decreased intestinal oxidative stress and influenced selected microbes without changing the messenger RNA levels of tight junction proteins in jejunal mucosa of weaning piglets. Animal 2017, 11, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Trevisi, P.; Merialdi, G.; Mazzoni, M.; Casini, L.; Tittarelli, C.; De Filippi, S.; Minieri, L.; Lalatta-Costerbosa, G.; Bosi, P. Effect of dietary addition of thymol on growth, salivary and gastric function, immune response, and excretion of Salmonella enterica serovar Typhimurium, in weaning pigs challenged with this microbe strain. Ital. J. Anim. Sci. 2007, 6, 374–376. [Google Scholar] [CrossRef]
- Michiels, J.; Missotten, J.; Van Hoorick, A.; Ovyn, A.; Fremaut, D.; De Smet, S.; Dierick, N. Effects of dose and formulation of carvacrol and thymol on bacteria and some functional traits of the gut in piglets after weaning. Arch. Anim. Nutr. 2010, 64, 136–154. [Google Scholar] [CrossRef]
- Piva, A.; Grilli, E.; Messina, M.; Albonetti, S.; Pizzamiglio, V.; Cipollini, I.; Gatta, P.; Zaghini, G. Citric acid and thymol influence gastrointestinal microflora in pigs at weaning. Poult. Sci. 2007, 86, 309. [Google Scholar]
- Anderson, R.C.; Krueger, N.A.; Genovese, K.J.; Stanton, T.B.; MacKinnon, K.M.; Harvey, R.B.; Edrington, T.S.; Callaway, T.R.; Nisbet, A.J. Effect of thymol or diphenyliodonium chloride on performance, gut fermentation characteristics, and Campylobacter colonization in growing swine. J. Food Prot. 2012, 75, 758–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omonijo, F.A.; Liu, S.; Hui, Q.; Zhang, H.; Lahaye, L.; Bodin, J.-C.; Gong, J.; Nyachoti, M.; Yang, C. Thymol Improves Barrier Function and Attenuates Inflammatory Responses in Porcine Intestinal Epithelial Cells during Lipopolysaccharide (LPS)-Induced Inflammation. J. Agric. Food Chem. 2019, 67, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Stevanović, Z.D.; Bošnjak-Neumüller, J.; Pajić-Lijaković, I.; Raj, J.; Vasiljević, M. Essential Oils as Feed Additives-Future Perspectives. Molecules 2018, 23, 1717. [Google Scholar] [CrossRef] [Green Version]
- Si, W.; Gong, J.; Chanas, C.; Cui, S.; Yu, H.; Caballero, C.; Friendship, R. In vitro assessment of antimicrobial activity of carvacrol, thymol and cinnamaldehyde towards Salmonella serotype Typhimurium DT104: Effects of pig diets and emulsification in hydrocolloids. J. Appl. Microbiol. 2006, 101, 1282–1291. [Google Scholar] [CrossRef]
- Putaala, H.; Nurminen, P.; Tiihonen, K. Effects of cinnamaldehyde and thymol on cytotoxicity, tight junction barrier resistance, and cyclooxygenase-1 and -2 expression in Caco-2 cells. J. Anim. Feed Sci. 2017, 26, 274–284. [Google Scholar] [CrossRef] [Green Version]
- Crespy, V.; Morand, C.; Besson, C.; Manach, C.; Demigne, C.; Remesy, C. Quercetin, but not its glycosides, is absorbed from the rat stomach. J. Agric. Food Chem. 2002, 50, 618–621. [Google Scholar] [CrossRef]
- Piskula, M.K.; Yamakoshi, J.; Iwai, Y. Daidzein and genistein but not their glucosides are absorbed from the rat stomach. FEBS Lett. 1999, 447, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Talavéra, S.v.; Felgines, C.; Texier, O.; Besson, C.; Lamaison, J.-L.; Rémésy, C. Anthocyanins Are Efficiently Absorbed from the Stomach in Anesthetized Rats. J. Nutr. 2003, 133, 4178–4182. [Google Scholar] [CrossRef]
- Cermak, R.; Landgraf, S.; Wolffram, S. The bioavailability of quercetin in pigs depends on the glycoside moiety and on dietary factors. J. Nutr. 2003, 133, 2802–2807. [Google Scholar] [CrossRef] [Green Version]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Xiang, Q.; Wang, J.; Peng, J.; Wei, H. Oregano essential oil improves intestinal morphology and expression of tight junction proteins associated with modulation of selected intestinal bacteria and immune status in a pig model. Biomed. Res. Int. 2016, 2016, 5436738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janczyk, P.; Trevisi, P.; Souffrant, W.B.; Bosi, P. Effect of thymol on microbial diversity in the porcine jejunum. Int. J. Food Microbiol. 2008, 126, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Day, A.J.; Gee, J.M.; DuPont, M.S.; Johnson, I.T.; Williamson, G. Absorption of quercetin-3-glucoside and quercetin-4′-glucoside in the rat small intestine: The role of lactase phlorizin hydrolase and the sodium-dependent glucose transporter. Biochem. Pharmacol. 2003, 65, 1199–1206. [Google Scholar] [CrossRef]
- Epps, S.V.; Petrujkić, B.T.; Sedej, I.; Krueger, N.A.; Harvey, R.B.; Beier, R.C.; Stanton, T.B.; Phillips, T.D.; Anderson, R.C.; Nisbet, D.J. Comparison of anti-Campylobacter activity of free thymol and thymol-β-d-glucopyranoside in absence or presence of β-glycoside-hydrolysing gut bacteria. Food Chem. 2015, 173, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Levent, G.; Harvey, R.B.; Ciftcioglu, G.; Beier, R.C.; Genovese, K.J.; He, H.L.; Anderson, R.C.; Nisbet, D.J. In Vitro Effects of Thymol-beta-D-Glucopyranoside on Salmonella enterica Serovar Typhimurium and Escherichia coli K88. J. Food Prot. 2016, 79, 299–303. [Google Scholar] [CrossRef]
- Li, Y.; Fu, X.; Geng, S.; Huang, Q.; Ma, X.; Jiang, X.; Han, X. Intestinal Microbiome-Metabolome Responses to Essential Oils in Piglets. Front. Microbiol. 2018, 9, 1988. [Google Scholar] [CrossRef]
- Looft, T.; Allen, H.K.; Cantarel, B.L.; Levine, U.Y.; Bayles, D.O.; Alt, D.P.; Henrissat, B.; Stanton, T.B. Bacteria, phages and pigs: The effects of in-feed antibiotics on the microbiome at different gut locations. ISME J. 2014, 8, 1566–1576. [Google Scholar] [CrossRef] [Green Version]
- Argüello, H.; Estellé, J.; Zaldívar-López, S.; Jiménez-Marín, Á.; Carvajal, A.; López-Bascón, M.A.; Crispie, F.; O’Sullivan, O.; Cotter, P.D.; Priego-Capote, F. Early Salmonella typhimurium infection in pigs disrupts microbiome composition and functionality principally at the ileum mucosa. Sci. Rep. 2018, 8, 7788. [Google Scholar] [CrossRef] [Green Version]
- Isaacson, R.; Kim, H.B. The intestinal microbiome of the pig. Anim. Health Res. Rev. 2012, 13, 100–109. [Google Scholar] [CrossRef]
- Yang, H.; Huang, X.; Fang, S.; Xin, W.; Huang, L.; Chen, C. Uncovering the composition of microbial community structure and metagenomics among three gut locations in pigs with distinct fatness. Sci. Rep. 2016, 6, 27427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janczyk, P.; Pieper, R.; Smidt, H.; Souffrant, W.B. Changes in the diversity of pig ileal lactobacilli around weaning determined by means of 16S rRNA gene amplification and denaturing gradient gel electrophoresis. FEMS Microbiol. Ecol. 2007, 61, 132–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konstantinov, S.R.; Awati, A.; Smidt, H.; Williams, B.A.; Akkermans, A.D.; De Vos, W.M. Specific response of a novel and abundant Lactobacillus amylovorus-like phylotype to dietary prebiotics in the guts of weaning piglets. Appl. Environ. Microbiol. 2004, 70, 3821–3830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kant, R.; Paulin, L.; Alatalo, E.; de Vos, W.M.; Palva, A. Genome sequence of Lactobacillus amylovorus GRL1118, isolated from pig ileum. J. Bacteriol. 2011, 193, 3147–3148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maenner, K.; Vahjen, W.; Simon, O. Studies on the effects of essential-oil-based feed additives on performance, ileal nutrient digestibility, and selected bacterial groups in the gastrointestinal tract of piglets. J. Anim. Sci. 2011, 89, 2106–2112. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Piao, X.; Ru, Y.; Han, X.; Xue, L.; Zhang, H. Effects of adding essential oil to the diet of weaned pigs on performance, nutrient utilization, immune response and intestinal health. Asian Australas. J. Anim. Sci. 2012, 25, 1617–1626. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Hossain, M.; Kim, G.; Hwang, J.; Ji, H.; Yang, C. Effects of resveratrol and essential oils on growth performance, immunity, digestibility and fecal microbial shedding in challenged piglets. Asian Australas. J. Anim. Sci. 2013, 26, 683. [Google Scholar] [CrossRef] [Green Version]
- Jang, I.S.; Ko, Y.H.; Kang, S.Y.; Lee, C.Y. Effect of a commercial essential oil on growth performance, digestive enzyme activity and intestinal microflora population in broiler chickens. Anim. Feed Sci. Technol. 2007, 134, 304–315. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Zhang, Y.; Cheng, Y.; Wang, Y. Rapid screening and identification of α-glucosidase inhibitors from mulberry leaves using enzyme-immobilized magnetic beads coupled with HPLC/MS and NMR. Biomed. Chromatogr. 2013, 27, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Taslimi, P.; Caglayan, C.; Gulcin, İ. The impact of some natural phenolic compounds on carbonic anhydrase, acetylcholinesterase, butyrylcholinesterase, and α-glycosidase enzymes: An antidiabetic, anticholinergic, and antiepileptic study. J. Biochem. Mol. Toxicol. 2017, 31, e21995. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-H.; Wang, M.-S.; Zeng, X.-A.; Gong, D.-M.; Huang, Y.-B. An in vitro investigation of the inhibitory mechanism of β-galactosidase by cinnamaldehyde alone and in combination with carvacrol and thymol. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 3189–3198. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Compounds | Amount | Compounds | Amount |
|---|---|---|---|
| Ingredient Composition | Calculated Nutrient Composition | ||
| Wheat | 297 | NEv for pigs (MJ/kg) | 10.0 |
| Barley | 250 | Digestible 2 Lysine | 11.4 |
| Corn | 150 | Digestible 2 methionine + cysteine | 6.8 |
| Soybean meal | 76 | Digestible 2 threonine | 7.4 |
| Whey powder | 53 | Digestible 2 tryptophan | 2.4 |
| Toasted soybeans | 50 | Calcium | 6.6 |
| Sunflower meal | 31 | Digestible phosphorus | 3.1 |
| Potato protein | 30 | Sodium | 2.9 |
| Soybean oil | 23 | Potassium | 7.1 |
| Limestone (fine) | 11 | Chlorine | 4.0 |
| Sodium bicarbonate | 5.6 | Magnesium | 1.4 |
| Dicalcium phosphate | 5.3 | ||
| Premix minerals and vitamins 1 | 5 | ||
| Salt | 2.0 | Analyzed Nutrient Composition | |
| L-Lysine | 5.4 | Dry matter | 896 |
| L-Threonine | 2.3 | Crude ash | 47 |
| DL-Methionine | 1.9 | Crude protein | 174 |
| L-Valine | 1.1 | Crude fat | 55 |
| L-Tryptophan | 0.7 | Crude fibre | 36 |
| Phytase (10,000 FYT/g) | 0.1 | ||
| TCON 1 | TTHY | TTαG | SEM | p-Value | |
|---|---|---|---|---|---|
| Body Weight (kg) | |||||
| d0 | 6.52 | 6.52 | 6.52 | 0.23 | 0.999 2 |
| d5 | 6.24 | 6.56 | 6.59 | 0.10 | 0.294 |
| d13 | 8.11 | 8.41 | 8.56 | 0.17 | 0.550 |
| Average Daily Gain (g/d) | |||||
| d0–5 | −46 | 7 | 12 | 13.7 | 0.199 |
| d5–13 | 234 | 231 | 246 | 14.1 | 0.894 |
| d0–13 | 114 | 135 | 146 | 11.8 | 0.550 |
| Average Daily Feed Intake (g/d) | |||||
| d0–5 | 62 | 79 | 87 | 9.0 | 0.539 |
| d5–13 | 262 | 274 | 295 | 17.2 | 0.741 |
| d0–13 | 185 | 199 | 215 | 13.4 | 0.673 |
| Feed:Gain | |||||
| d0–5 | −0.88 | 5.06 | −4.67 | 1.62 | 0.082 |
| d5–13 | 1.14 | 1.19 | 1.20 | 0.04 | 0.811 |
| d0–13 | 1.54 | 1.48 | 1.50 | 0.04 | 0.823 |
| Fecal Consistency | |||||
| Average Score 3 | 1.81 a | 1.53 b | 1.60 ab | 0.20 | 0.038 |
| Diarrhea incidence (%) | 24.3 | 12.2 | 20.3 | 2.1 | 0.086 |
| TCON 1 | TTHY | TTαG | SEM | p-Value | |
|---|---|---|---|---|---|
| Apparent Permeability (*10−7 cm/s)2 | |||||
| FD4 3 | 6.54 a | 2.88 b | 6.32 ab | 0.84 | 0.028 4 |
| HRP 3 | 3.02 | 0.67 | 1.31 | 0.47 | 0.143 4 |
| Histo-Morphology 2 | |||||
| Villus Height (µm) | 513 | 584 | 548 | 15 | 0.160 |
| Crypt depth (µm) | 205 | 205 | 200 | 3 | 0.715 |
| Villus height:crypt depth | 2.57 | 2.88 | 2.77 | 0.06 | 0.071 |
| TCON 1 | TTHY | TTαG | SEM | p-Value | |
|---|---|---|---|---|---|
| SI1 2 | |||||
| Lactobacilli | 7.38 | 7.36 | 7.46 | 0.09 | 0.898 |
| Total anaerobes | 6.60 | 6.72 | 6.39 | 0.14 | 0.334 3 |
| E. coli | 3.44 ab | 3.35 b | 4.29 a | 0.16 | 0.041 |
| Total coliforms | 4.21 | 3.33 | 3.81 | 0.22 | 0.267 |
| Streptococci | 7.31 | 7.45 | 7.41 | 0.09 | 0.820 |
| SI3 | |||||
| Lactobacilli | 8.71 | 8.38 | 8.85 | 0.08 | 0.070 |
| Total anaerobes | 7.53 | 7.07 | 7.44 | 0.09 | 0.069 |
| E. coli | 5.51 | 5.26 | 6.22 | 0.17 | 0.078 |
| Total coliforms | 5.48 | 5.22 | 6.21 | 0.17 | 0.069 |
| Streptococci | 8.44 | 8.45 | 8.79 | 0.09 | 0.174 |
| TCON 1 | TTHY | TTαG | SEM | p-Value | |
|---|---|---|---|---|---|
| SI3 2 | |||||
| Total Volatile Fatty Acids | 6.44 | 7.47 | 5.65 | 0.63 | 0.504 |
| % Acetate | 96.6 | 98.2 | 98.3 | 0.6 | 0.330 3 |
| % Propionate | 3.2 | 1.3 | 1.5 | 0.6 | 0.132 3 |
| % Butyrate | 0.3 | 0.6 | 0.2 | 0.1 | 0.349 3 |
| Lactate | 53.45 | 42.42 | 61.30 | 5.04 | 0.320 |
| Mid-colon | |||||
| Total Volatile Fatty Acids | 164.9 | 176.0 | 162.1 | 3.55 | 0.251 |
| % Acetate | 53.7 | 55.4 | 55.0 | 0.7 | 0.505 4 |
| % Propionate | 27.0 | 26.2 | 27.3 | 0.6 | 0.699 |
| % Iso-butyrate | 1.0 | 1.1 | 0.7 | 0.1 | 0.182 |
| % Butyrate | 13.2 | 13.3 | 12.4 | 0.4 | 0.703 |
| % Iso-valerate | 1.4 | 1.5 | 1.2 | 0.1 | 0.254 4 |
| % Valerate | 3.8 | 2.7 | 3.3 | 0.2 | 0.088 |
| Lactate | 22.14 | 23.53 | 21.08 | 0.53 | 0.185 |
| TCON 1 | TTHY | TTαG | SEM | p-Value | |
|---|---|---|---|---|---|
| Mucosa (mmol/min/g) | |||||
| α-Glucosidase | |||||
| Duodenum | 0.84 | 0.87 | 0.87 | 0.03 | 0.863 |
| Proximal Jejunum | 1.17 | 1.06 | 1.25 | 0.06 | 0.399 3 |
| Distal Jejunum | 1.06 | 1.10 | 1.29 | 0.06 | 0.350 3 |
| β-Glucosidase | |||||
| Duodenum | 12.03 a | 7.95 b | 11.25 ab | 0.62 | 0.027 |
| Proximal Jejunum | 11.93 | 9.31 | 10.54 | 0.67 | 0.299 |
| Distal Jejunum | 4.88 | 4.66 | 4.41 | 0.39 | 0.882 |
| β-Galactosidase | |||||
| Duodenum | 18.93 a | 13.25 b | 17.68 ab | 0.85 | 0.026 |
| Proximal Jejunum | 21.23 | 17.59 | 18.40 | 0.95 | 0.276 |
| Distal Jejunum | 14.39 | 15.70 | 18.59 | 1.06 | 0.269 |
| Digesta (nmol/min/g) | |||||
| α-Glucosidase | |||||
| Stomach | 25.0 | 24.2 | 17.7 | 1.8 | 0.212 |
| SI12 | 113.1 | 67.4 | 113.2 | 11.5 | 0.190 |
| SI2 | 610.1 | 473.3 | 578.4 | 46.7 | 0.508 4 |
| SI3 | 1303.0 | 1097.5 | 1248.4 | 63.2 | 0.348 |
| Caecum | 479.7 | 357.0 | 433.0 | 53.9 | 0.596 |
| Mid-colon | 441.7 | 450.0 | 545.6 | 40.4 | 0.489 |
| β-Glucosidase | |||||
| Stomach | 39.1 | 36.6 | 27.3 | 3.2 | 0.271 3 |
| SI1 | 325.0 | 290.3 | 319.8 | 38.9 | 0.926 |
| SI2 | 1276.1 | 1033.9 | 1153.7 | 124.8 | 0.643 4 |
| SI3 | 1852.5 | 1684.1 | 1942.5 | 202.2 | 0.863 |
| Caecum | 674.8 | 564.8 | 663.2 | 98.8 | 0.650 4 |
| Mid-colon | 838.1 | 585.8 | 512.1 | 81.3 | 0.280 3 |
| β-Galactosidase | |||||
| Stomach | 22.6 | 22.2 | 19.8 | 2.0 | 0.829 |
| SI1 | 264.1 | 238.0 | 276.6 | 31.3 | 0.862 |
| SI2 | 1132.4 | 953.9 | 963.6 | 113.5 | 0.715 4 |
| SI3 | 1673.0 | 1396.6 | 1831.3 | 186.9 | 0.619 |
| Caecum | 1341.0 | 1583.2 | 1715.3 | 246.9 | 0.818 |
| Mid-colon | 1846.9 | 1815.6 | 2103.6 | 144.4 | 0.668 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Noten, N.; Degroote, J.; Van Liefferinge, E.; Taminiau, B.; De Smet, S.; Desmet, T.; Michiels, J. Effects of Thymol and Thymol α-D-Glucopyranoside on Intestinal Function and Microbiota of Weaned Pigs. Animals 2020, 10, 329. https://doi.org/10.3390/ani10020329
Van Noten N, Degroote J, Van Liefferinge E, Taminiau B, De Smet S, Desmet T, Michiels J. Effects of Thymol and Thymol α-D-Glucopyranoside on Intestinal Function and Microbiota of Weaned Pigs. Animals. 2020; 10(2):329. https://doi.org/10.3390/ani10020329
Chicago/Turabian StyleVan Noten, Noémie, Jeroen Degroote, Elout Van Liefferinge, Bernard Taminiau, Stefaan De Smet, Tom Desmet, and Joris Michiels. 2020. "Effects of Thymol and Thymol α-D-Glucopyranoside on Intestinal Function and Microbiota of Weaned Pigs" Animals 10, no. 2: 329. https://doi.org/10.3390/ani10020329