Secondary Functions of Arginine and Sulfur Amino Acids in Poultry Health: Review


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Arginine
2.1. Arginine Metabolism
2.2. Arginine: Role in Fat, Bone, and Muscle Metabolism
3. Total Sulfur Amino Acid
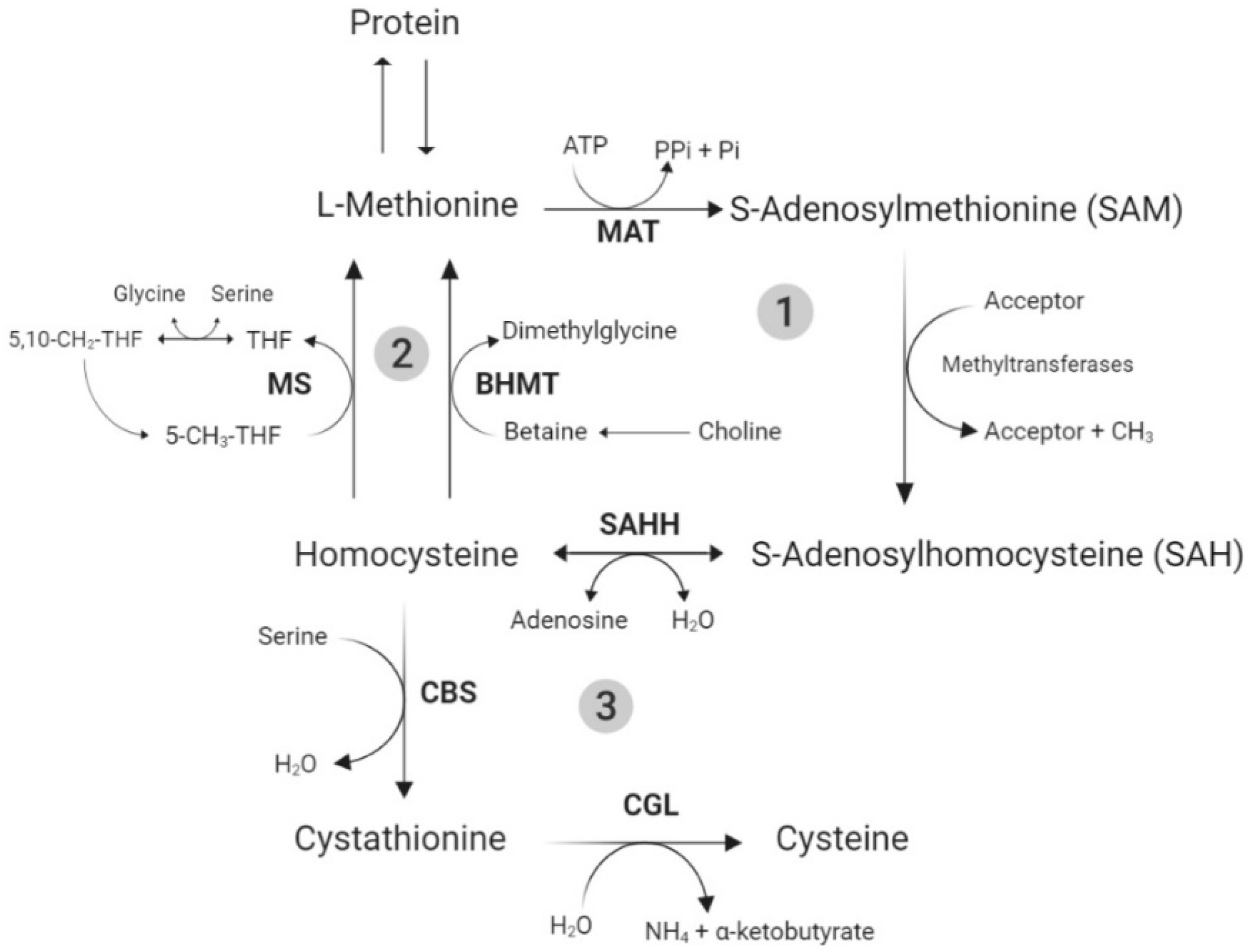
3.1. Total Sulfur Amino Acid Metabolism
3.2. Total Sulfur Amino Acids: Role in Heat Stress and Bone Metabolism
4. Arginine and Total Sulfur Amino Acids Utilization under an Eimeria spp. Infection
4.1. Intestinal Digestion and Absorption
4.2. Antioxidant Capacity and Cellular Repair
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Dietary requirements of synthesizable amino acids by animals: A paradigm shift in protein nutrition. J. Anim. Sci. Biotechnol. 2014, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Marc Rhoads, J.; Carey Satterfield, M.; Smith, S.B.; Spencer, T.E.; Yin, Y. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009, 37, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Wu, G. Functional amino acids in growth, reproduction, and health. Adv. Nutr. 2010, 1, 31–37. [Google Scholar] [CrossRef]
- Ball, R.O.; Urschel, K.L.; Pencharz, P.B. Nutritional consequences of interspecies differences in arginine and lysine metabolism. J. Nutr. 2007, 137, 1626S–1641S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khajali, F.; Wideman, R.F. Dietary arginine: Metabolic, environmental, immunological and physiological interrelationships. World’s Poult. Sci. J. 2010, 66, 751–766. [Google Scholar] [CrossRef]
- Wu, L.Y.; Fang, Y.J.; Guo, X.Y. Dietary L-arginine supplementation beneficially regulates body fat deposition of meat-type ducks. Br. Poult. Sci. 2011, 52, 221–226. [Google Scholar] [CrossRef]
- Fouad, A.M.; El-Senousey, H.K.; Yang, X.J.; Yao, J.H. Dietary L-arginine supplementation reduces abdominal fat content by modulating lipid metabolism in broiler chickens. Animal 2013, 7, 1239–1245. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.I.; Murakami, A.E.; Martins, E.N.; Sakamoto, M.I.; Garcia, E.R. Effect of arginine on the development of the pectoralis muscle and the diameter and the protein:deoxyribonucleic acid rate of its skeletal myofibers in broilers. Poult. Sci. 2009, 88, 1399–1406. [Google Scholar] [CrossRef]
- Castro, F.L.S.; Su, S.; Choi, H.; Koo, E.; Kim, W.K. L-Arginine supplementation enhances growth performance, lean muscle, and bone density but not fat in broiler chickens. Poult. Sci. 2019, 98, 1716–1722. [Google Scholar] [CrossRef]
- Jankowski, J.; Mikulski, D.; Mikulska, M.; Ognik, K.; Całyniuk, Z.; Mróz, E.; Zduńczyk, Z. The effect of different dietary ratios of arginine, methionine, and lysine on the performance, carcass traits, and immune status of turkeys. Poult. Sci. 2020, 99, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Atakisi, O.; Atakisi, E.; Kart, A. Effects of dietary zinc and l-Arginine supplementation on total antioxidants capacity, lipid peroxidation, nitric oxide, egg weight, and blood biochemical values in japanase quails. Biol. Trace Elem. Res. 2009, 132, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Tayade, C.; Jaiswal, T.N.; Mishra, S.C.; Koti, M. l-Arginine stimulates immune response in chickens immunized with intermediate plus strain of infectious bursal disease vaccine. Vaccine 2006, 24, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Munir, K.; Muneer, M.A.; Masaoud, E.; Tiwari, A.; Mahmud, A.; Chaudhry, R.M.; Rashid, A. Dietary arginine stimulates humoral and cell-mediated immunity in chickens vaccinated and challenged against hydropericardium syndrome virus. Poult. Sci. 2009, 88, 1629–1638. [Google Scholar] [CrossRef] [PubMed]
- Corzo, A.; Moran Jr, E.T.; Hoehler, D. Arginine need of heavy broiler males: Applying the ideal protein concept. Poult. Sci. 2003, 82, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Dai, Z.; Li, D.; Wang, J.; Wu, Z. Amino acid nutrition in animals: Protein synthesis and beyond. Annu. Rev. Anim. Biosci. 2014, 2, 387–417. [Google Scholar] [CrossRef]
- Bunchasak, C. Role of dietary methionine in poultry production. J. Poult. Sci. 2009, 46, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Stipanuk, M.H. Sulfur amino acid metabolism: Pathways for production and removal of homocysteine and cysteine. Annu. Rev. Nutr. 2004, 24, 539–577. [Google Scholar] [CrossRef]
- Brake, J. Optimum dietary arginine: Lysine ratio for broiler chickens is altered during heat stress in association with changes in intestinal uptake and dietary sodium chloride. Br. Poult. Sci. 1998, 39, 639–647. [Google Scholar] [CrossRef]
- Lai, A.; Dong, G.; Song, D.; Yang, T.; Zhang, X. Responses to dietary levels of methionine in broilers medicated or vaccinated against coccidia under Eimeria tenella-challenged condition. BMC Vet. Res. 2018, 14, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M. Arginine metabolism revisited. J. Nutr. 2016, 146, 2579S–2586S. [Google Scholar] [CrossRef] [PubMed]
- Tamir, H.; Ratner, S. Enzymes of arginine metabolism in chicks. Arch. Biochem. Biophys. 1963, 102, 249–258. [Google Scholar] [CrossRef]
- Su, C.; Austic, R. The recycling of L-citrulline to L-arginine in a chicken macrophage cell line. Poult. Sci. 1999, 78, 353–355. [Google Scholar] [CrossRef] [PubMed]
- Tamir, H.; Ratner, S. A study of ornithine, citrulline and arginine synthesis in growing chicks. Arch. Biochem. Biophys. 1963, 102, 259–269. [Google Scholar] [CrossRef]
- Djeraba, A.; Musset, E.; van Rooijen, N.; Quéré, P. Resistance and susceptibility to Marek’s disease: Nitric oxide synthase/arginase activity balance. Vet. Microbiol. 2002, 86, 229–244. [Google Scholar] [CrossRef]
- Gonzalez-Esquerra, R.; Leeson, S. Concentrations of putrescine, spermidine, and spermine in duodenum and pancreas as affected by the ratio of arginine to lysine and source of methionine in broilers under heat stress. Poult. Sci. 2006, 85, 1398–1408. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Knabe, D.A.; Li, P.; Li, X.; McKnight, J.R.; Satterfield, M.C.; et al. Proline and hydroxyproline metabolism: Implications for animal and human nutrition. Amino Acids 2011, 40, 1053–1063. [Google Scholar] [CrossRef] [Green Version]
- Rath, M.; Müller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via arginase or nitric oxide synthase: Two competing arginine pathways in macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef] [Green Version]
- Jobgen, W.S.; Fried, S.K.; Fu, W.J.; Meininger, C.J.; Wu, G. Regulatory role for the arginine-nitric oxide pathway in metabolism of energy substrates. J. Nutr. Biochem. 2006, 17, 571–588. [Google Scholar] [CrossRef]
- Fouad, A.M.; El-Senousey, H.K. Nutritional factors affecting abdominal fat deposition in poultry: A review. Asian Australas J. Anim. Sci. 2014, 27, 1057–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolea, S.; Pertusa, J.A.G.; Martín, F.; Sanchez-Andrés, J.V.; Soria, B. Regulation of pancreatic β-cell electrical activity and insulin release by physiological amino acid concentrations. Pflügers Arch. 1997, 433, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Collier, S.R.; Casey, D.P.; Kanaley, J.A. Growth hormone responses to varying doses of oral arginine. Growth Horm. Igf Res. 2005, 15, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Houston, B.; O’Neill, I. Insulin and growth hormone act synergistically to stimulate insulin-like growth factor-I production by cultured chicken hepatocytes. J. Endocrinol. 1991, 128, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Duclos, M.J. Insulin-loke growth factor-I (IGF-I) mRNA levels and chicken muscle growth. J. Physiol. Pharmacol. 2005, 56, 25–35. [Google Scholar] [PubMed]
- Florini, J.R.; Ewton, D.Z.; Coolican, S.A. Growth hormone and the insulin-like growth factor system in myogenesis. Endocr. Rev. 1996, 17, 481–517. [Google Scholar]
- Tomas, F.M.; Pym, R.A.; McMurtry, J.P.; Francis, G.L. Insulin-like growth factor (IGF)-I but not IGF-II promotes lean growth and feed efficiency in broiler chickens. Gen. Comp. Endocrinol. 1998, 110, 262–275. [Google Scholar] [CrossRef]
- Conlon, M.A.; Kita, K. Muscle protein synthesis rate is altered in response to a single injection of insulin-like growth factor-I in seven-day-old Leghorn chicks. Poult. Sci. 2002, 81, 1543–1547. [Google Scholar] [CrossRef]
- Chamruspollert, M.; Pesti, G.M.; Bakalli, R.I. Dietary interrelationships among arginine, methionine, and lysine in young broiler chicks. Br. J. Nutr. 2002, 88, 655–660. [Google Scholar] [CrossRef]
- Chen, J.; Wang, M.; Kong, Y.; Ma, H.; Zou, S. Comparison of the novel compounds creatine and pyruvateon lipid and protein metabolism in broiler chickens. Animal 2011, 5, 1082–1089. [Google Scholar] [CrossRef] [Green Version]
- Subramaniyan, S.A.; Kang, D.R.; Park, J.R.; Siddiqui, S.H.; Ravichandiran, P.; Yoo, D.J.; Na, C.S.; Shim, K.S. Effect of in ovo injection of l-arginine in different chicken embryonic development stages on post-hatchability, immune response, and Myo-D and myogenin proteins. Animals 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevalley, T.; Rizzoli, R.; Manen, D.; Caverzasio, J.; Bonjour, J.-P. Arginine increases insulin-like growth factor-I production and collagen synthesis in osteoblast-like cells. Bone 1998, 23, 103–109. [Google Scholar] [CrossRef]
- Hayden, J.M.; Mohan, S.; Baylink, D.J. The insulin-like growth factor system and the coupling of formation to resorption. Bone 1995, 17, S93–S98. [Google Scholar] [CrossRef]
- Spagnoli, A.; Rosenfeld, R.G. The mechanisms by which growth hormone brings about growth: The relative contributions of growth hormone and insulin-like growth factors. Endocrinol. Metab. Clin. N. Am. 1996, 25, 615–631. [Google Scholar] [CrossRef]
- Olney, R.C. Regulation of bone mass by growth hormone. Med. Pediatric. Oncol. 2003, 41, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Fini, M.; Torricelli, P.; Giavaresi, G.; Carpi, A.; Nicolini, A.; Giardino, R. Effect of L-lysine and L-arginine on primary osteoblast cultures from normal and osteopenic rats. Biomed. Pharmacother. 2001, 55, 213–220. [Google Scholar] [CrossRef]
- Riancho, J.A.; Salas, E.; Zarrabeitia, M.T.; Olmos, J.M.; Amado, J.A.; Fernández-Luna, J.L.; González-Macías, J. Expression and functional role of nitric oxide synthase in osteoblast-like cells. J. Bone Miner. Res. 1995, 10, 439–446. [Google Scholar] [CrossRef]
- Baldik, Y.; Talu, U.; Altinel, L.; Bilge, H.; Demiryont, M.; Aykac-Toker, G. Bone healing regulated by nitric oxide: An experimental study in rats. Clin. Orthop. Relat. Res. 2002, 404, 343–352. [Google Scholar] [CrossRef]
- Mancini, L.; Moradi-Bidhendi, N.; Becherini, L.; Martineti, V.; MacIntyre, I. The biphasic effects of nitric oxide in primary rat osteoblasts are cGMP dependent. Biochem. Biophys. Res. Commun. 2000, 274, 477–481. [Google Scholar] [CrossRef]
- Damoulis, P.D.; Hauschka, P.V. Nitric oxide acts in conjunction with proinflammatory cytokines to promote cell death in osteoblasts. J. Bone Miner. Res. 1997, 12, 412–422. [Google Scholar] [CrossRef]
- Minne, H.W.; Pfeilschifter, J.; Scharla, S.; Mutschelknauss, S.; Schwarz, A.; Krempien, B.; Ziegler, R. Inflammation-mediated osteopenia in the rat: A new animal model for pathological loss of bone mass*. Endocrinology 1984, 115, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Croucher, P.I.; Vedi, S.; Motley, R.J.; Garrahan, N.J.; Stanton, M.R.; Compston, J.E. Reduced bone formation in patients with osteoporosis associated with inflammatory bowel disease. Osteoporos. Int. 1993, 3, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Armour, K.E.; Van ’T Hof, R.J.; Grabowski, P.S.; Reid, D.M.; Ralston, S.H. Evidence for a pathogenic role of nitric oxide in inflammation-induced osteoporosis. J. Bone Miner. Res. 1999, 14, 2137–2142. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T.; Brosnan, M.E. The sulfur-containing amino acids: An overview. J. Nutr. 2006, 136, 1636S–1640S. [Google Scholar] [CrossRef] [Green Version]
- Métayer, S.; Seiliez, I.; Collin, A.; Duchêne, S.; Mercier, Y.; Geraert, P.-A.; Tesseraud, S. Mechanisms through which sulfur amino acids control protein metabolism and oxidative status. J. Nutr. Biochem. 2008, 19, 207–215. [Google Scholar] [CrossRef]
- Martin-Venegas, R.; Geraert, P.A.; Ferrer, R. Conversion of the methionine hydroxy analogue DL-2-Hydroxy-(4-Methylthio) butanoic acid to sulfur-containing amino acids in the chicken small intestine. Poult. Sci. 2006, 85, 1932–1938. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Kidd, M.T. Taurine in poultry nutrition. Anim. Feed Sci. Technol. 2020, 260, 114339. [Google Scholar] [CrossRef]
- Grimble, R.F.; Jackson, A.A.; Persaud, C.; Wride, M.J.; Delers, F.; Engler, R. Cysteine and glycine supplementation modulate the metabolic response to tumor necrosis factor α in rats fed a low protein diet. J. Nutr. 1992, 122, 2066–2073. [Google Scholar] [CrossRef]
- Wu, G.; Fang, Y.-Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione Metabolism and Its Implications for Health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef] [Green Version]
- Huseby, N.; Sundkvist, E.; Svineng, G. Glutathione and sulfur containing amino acids: Antioxidant and conjugation activities. In Glutathione and Sulfur Amino Acids in Human Health and Disease; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2008; pp. 91–120. ISBN 978-0-470-47597-3. [Google Scholar]
- Zhao, F.Q.; Zhang, Z.W.; Qu, J.P.; Yao, H.D.; Li, M.; Li, S.; Xu, S.W. Cold stress induces antioxidants and Hsps in chicken immune organs. Cell Stress Chaperones 2014, 19, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Akbarian, A.; Michiels, J.; Degroote, J.; Majdeddin, M.; Golian, A.; De Smet, S. Association between heat stress and oxidative stress in poultry; Mitochondrial dysfunction and dietary interventions with phytochemicals. J. Anim. Sci. Biotechnol. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simitzis, P.E.; Kalogeraki, E.; Goliomytis, M.; Charismiadou, M.A.; Triantaphyllopoulos, K.; Ayoutanti, A.; Niforou, K.; Hager-Theodorides, A.L.; Deligeorgis, S.G. Impact of stocking density on broiler growth performance, meat characteristics, behavioural components and indicators of physiological and oxidative stress. Br. Poult. Sci. 2012, 53, 721–730. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. Economic losses from heat stress by US livestock industries. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef] [Green Version]
- Mashaly, M.M.; Hendricks, G.L.; Kalama, M.A.; Gehad, A.E.; Abbas, A.O.; Patterson, P.H. Effect of heat stress on production parameters and immune responses of commercial laying hens. Poult. Sci. 2004, 83, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Wallis, I.R.; Balnave, D. The influence of environmental temperature, age and sex on the digestibility of amino acids in growing broiler chickens. Br. Poult. Sci. 1984, 25, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Zuprizal; Larbier, M.; Chagneau, A.M.; Geraert, P.A. Influence of ambient temperature on true digestibility of protein and amino acids of rapeseed and soybean meals in broilers. Poult. Sci. 1993, 72, 289–295. [Google Scholar] [CrossRef]
- Attia, Y.A.; Hassan, R.A.; Qota, E.M.A. Recovery from adverse effects of heat stress on slow-growing chicks in the tropics 1: Effect of ascorbic acid and different levels of betaine. Trop. Anim. Health Prod. 2009, 41, 807–818. [Google Scholar] [CrossRef]
- Attia, Y.A.; Abd El-Hamid, A.E.-H.E.; Abedalla, A.A.; Berika, M.A.; Al-Harthi, M.A.; Kucuk, O.; Sahin, K.; Abou-Shehema, B.M. Laying performance, digestibility and plasma hormones in laying hens exposed to chronic heat stress as affected by betaine, vitamin C, and/or vitamin E supplementation. SpringerPlus 2016, 5, 1619–1631. [Google Scholar] [CrossRef] [Green Version]
- Wolfenson, D.; Frei, Y.F.; Snapir, N.; Berman, A. Effect of diurnal or nocturnal heat stress on egg formation. Br. Poult. Sci. 1979, 20, 167–174. [Google Scholar] [CrossRef]
- Mitchell, M.A.; Carlisle, A.J. The effects of chronic exposure to elevated environmental temperature on intestinal morphology and nutrient absorption in the domestic fowl (Gallus domesticus). Comp. Biochem. Physiol. Part A Physiol. 1992, 101, 137–142. [Google Scholar] [CrossRef]
- Ma, X.; Lin, Y.; Zhang, H.; Chen, W.; Wang, S.; Ruan, D.; Jiang, Z. Heat stress impairs the nutritional metabolism and reduces the productivity of egg-laying ducks. Anim. Reprod. Sci. 2014, 145, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Dibner, J.J.; Atwell, C.A.; Ivey, F.J. Effect of heat stress on 2-Hydroxy-4-(Methylthio)butanoic acid and dl-methionine absorption measured in vitro. Poult. Sci. 1992, 71, 1900–1910. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.D.; Wuelling, C.W.; Atwell, C.A.; Dibner, J.J. Effect of intermittent periods of high environmental temperature on broiler performance responses to sources of methionine activity. Poult. Sci. 1994, 73, 627–639. [Google Scholar] [CrossRef] [PubMed]
- del Vesco, A.P.; Gasparino, E.; de Grieser, D.O.; Zancanela, V.; Soares, M.A.M.; de Oliveira Neto, A.R. Effects of methionine supplementation on the expression of oxidative stress-related genes in acute heat stress-exposed broilers. Br. J. Nutr. 2015, 113, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Elwan, H.A.M.; Elnesr, S.S.; Xu, Q.; Xie, C.; Dong, X.; Zou, X. Effects of in ovo methionine-cysteine injection on embryonic development, antioxidant status, IGF-I and TLR4 gene expression, and jejunum histomorphometry in newly hatched broiler chicks exposed to heat stress during incubation. Animals 2019, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Gregory, N.G.; Wilkins, L.J. Broken bones in domestic fowl: Handling and processing damage in end-of-lay battery hens. Br. Poult. Sci. 1989, 30, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Casey-Trott, T.M.; Widowski, T.M. Behavioral differences of laying hens with fractured keel bones within furnished cages. Front. Vet. Sci. 2016, 3, 42. [Google Scholar] [CrossRef] [Green Version]
- Knowles, T.G.; Kestin, S.C.; Haslam, S.M.; Brown, S.N.; Green, L.E.; Butterworth, A.; Pope, S.J.; Pfeiffer, D.; Nicol, C.J. Leg disorders in broiler chickens: Prevalence, risk factors and prevention. PLoS ONE 2008, 3, e1545. [Google Scholar] [CrossRef] [Green Version]
- Fleming, R.H. Nutritional factors affecting poultry bone health: Symposium on ‘Diet and bone health’. Proc. Nutr. Soc. 2008, 67, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Heaney, R.P.; Layman, D.K. Amount and type of protein influences bone health. Am. J. Clin. Nutr. 2008, 87, 1567S–1570S. [Google Scholar] [CrossRef] [Green Version]
- del Vesco, A.; Gasparino, E.; Oliveira Neto, A.; Guimarães, S.E.F.; Marcato, S.M.M.; Voltolini, D. Dietary methionine effects on IGF-I and GHR mRNA expression in broilers. Genet. Mol. Res. Gmr. 2013, 12, 6414–6423. [Google Scholar] [CrossRef] [PubMed]
- Castro, F.L.S.; Kim, H.Y.; Hong, Y.G.; Kim, W.K. The effect of total sulfur amino acid levels on growth performance, egg quality, and bone metabolism in laying hens subjected to high environmental temperature. Poult. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Castro, F.L.S.; Kim, Y.; Xu, H.; Kim, W.K. The effect of total sulfur amino acid levels on growth performance and bone metabolism in pullets under heat stress. Poult. Sci. 2020, 99, 5783–5791. [Google Scholar] [CrossRef] [PubMed]
- Morgan, S.; Poundarik, A.A.; Vashishth, D. Do non-collagenous proteins affect skeletal mechanical properties? Calcif. Tissue Int. 2015, 97, 281–291. [Google Scholar] [CrossRef]
- Clarke, B. Normal bone anatomy and physiology. Clin. J. Am. Soc. Nephrol. 2008, 3 (Suppl. 3), S131–S139. [Google Scholar] [CrossRef] [Green Version]
- Heinegård, D.; Oldberg, A. Structure and biology of cartilage and bone matrix noncollagenous macromolecules. Faseb J. 1989, 3, 2042–2051. [Google Scholar] [CrossRef] [Green Version]
- Al-Qtaitat, A.I. A review of non-collagenous proteins; Their Role in bone. AJLS 2014, 2, 351. [Google Scholar] [CrossRef]
- Windmueller, H.G.; Spaeth, A.E. Metabolism of absorbed aspartate, asparagine, and arginine by rat small intestine in vivo. Arch. Biochem. Biophys. 1976, 175, 670–676. [Google Scholar] [CrossRef]
- Castillo, L.; Chapman, T.E.; Yu, Y.M.; Ajami, A.; Burke, J.F.; Young, V.R. Dietary arginine uptake by the splanchnic region in adult humans. Am. J. Physiol. Endocrinol. Metab. 1993, 265, E532–E539. [Google Scholar] [CrossRef]
- Stoll, B.; Henry, J.; Reeds, P.J.; Yu, H.; Jahoor, F.; Burrin, D.G. Catabolism dominates the first-pass intestinal metabolism of dietary essential amino acids in milk protein-fed piglets. J. Nutr. 1998, 128, 606–614. [Google Scholar] [CrossRef]
- Shoveller, A.K.; Brunton, J.A.; Pencharz, P.B.; Ball, R.O. The methionine requirement is lower in neonatal piglets fed parenterally than in those fed enterally. J. Nutr. 2003, 133, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Shoveller, A.K.; Stoll, B.; Ball, R.O.; Burrin, D.G. Nutritional and functional importance of intestinal sulfur amino acid metabolism. J. Nutr. 2005, 135, 1609–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.B. Intercurrent coccidiosis and necrotic enteritis of chickens: Rational, integrated disease management by maintenance of gut integrity. Avian Pathol. 2005, 34, 159–180. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.B. A compartmentalised model for the estimation of the cost of coccidiosis to the world’s chicken production industry. Int. J. Parasitol. 1999, 29, 1209–1229. [Google Scholar] [CrossRef]
- Koinarski, V.; Georgieva, N.; Gadjeva, V.; Petkov, P. Antioxidant status of broiler chickens, infected with Eimeria acervulina. Rev. Méd. Vét. 2005, 5, 498. [Google Scholar]
- Hasegawa, H.; Shinohara, Y.; Akahane, K.; Hashimoto, T. Direct detection and evaluation of conversion of D-methionine into L-methionine in rats by stable isotope methodology. J. Nutr. 2005, 135, 2001–2005. [Google Scholar] [CrossRef]
- Yegani, M.; Korver, D.R. Factors Affecting Intestinal Health in Poultry. Poult. Sci. 2008, 87, 2052–2063. [Google Scholar] [CrossRef]
- McDougald, L.R. Protozoal infections. In Diseases of Poultry; Swayne, D.E., Ed.; John wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 1147–1167. [Google Scholar]
- Kipper, M.; Andretta, I.; Lehnen, C.R.; Lovatto, P.A.; Monteiro, S.G. Meta-analysis of the performance variation in broilers experimentally challenged by Eimeria spp. Vet. Parasitol. 2013, 196, 77–84. [Google Scholar] [CrossRef]
- Fernando, M.A.; McCraw, B.M. Mucosal morphology and cellular renewal in the intestine of chickens following a single infection of eimeria acervulina. J. Parasitol. 1973, 59, 493–501. [Google Scholar] [CrossRef]
- Witlock, D.R.; Ruff, M.D. Comparison of the intestinal surface damage caused by eimeria mivati, E. necatrix, E. maxima, E. brunetti, and E. acervulina by scanning electron microscopy. J. Parasitol. 1977, 63, 193–199. [Google Scholar] [CrossRef]
- Russell, J.; Ruff, M.D. Eimeria Spp.: Influence of coccidia on digestion (amylolytic activity) in broiler chickens. Exp. Parasitol. 1978, 45, 234–240. [Google Scholar] [CrossRef]
- Allen, P.C. Physiological responses of chicken gut tissue to coccidial infection: Comparative effects of eimeria acervulina and eimeria mitis on mucosal mass, carotenoid content, and brush border enzyme activity. Poult. Sci. 1987, 66, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.; Vahl, H.A.; Veldman, A. Interaction between nutrition and Eimeria acervulina infection in broiler chickens: Development of an experimental infection model. Br. J. Nutr. 1996, 75, 867–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruff, M.D. Reduced transport of methionine in intestines of chickens infected with eimeria necatrix. J. Parasitol. 1974, 60, 838. [Google Scholar] [CrossRef] [PubMed]
- Ruff, M.D.; Witlock, D.R.; Smith, R.R. Eimeria acervulina and E. tenella: Effect on methionine absorption by the avian intestine. Exp. Parasitol. 1976, 39, 244–251. [Google Scholar] [CrossRef]
- Ruff, M.D.; Wilkins, G.C. Total intestinal absorption of glucose and l-methionine in broilers infected with Eimeria acervulina, E. mivati, E. maxima or E. brunetti. Parasitology 1980, 80, 555. [Google Scholar] [CrossRef]
- Persia, M.E.; Young, E.L.; Utterback, P.L.; Parsons, C.M. Effects of dietary ingredients and eimeria acervulina infection on chick performance, apparent metabolizable energy, and amino acid digestibility. Poult. Sci. 2006, 85, 48–55. [Google Scholar] [CrossRef]
- Rochell, S.J.; Parsons, C.M.; Dilger, R.N. Effects of Eimeria acervulina infection severity on growth performance, apparent ileal amino acid digestibility, and plasma concentrations of amino acids, carotenoids, and α1-acid glycoprotein in broilers. Poult. Sci. 2016, 95, 1573–1581. [Google Scholar] [CrossRef]
- Paris, N.E.; Wong, E.A. Expression of digestive enzymes and nutrient transporters in the intestine of Eimeria maxima-infected chickens. Poult. Sci. 2013, 92, 1331–1335. [Google Scholar] [CrossRef]
- Torras-Llort, M.; Torrents, D.; Soriano-García, J.F.; Gelpí, J.L.; Estévez, R.; Ferrer, R.; Palacín, M.; Moretó, M. Sequential Amino Acid Exchange across b0,+-like System in Chicken Brush Border Jejunum. J. Membr. Biol. 2001, 180, 213–220. [Google Scholar] [CrossRef]
- Verrey, F. System L: Heteromeric exchangers of large, neutral amino acids involved in directional transport. Pflügers Arch. Eur. J. Physiol. 2003, 445, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Estévez, M. Oxidative damage to poultry: From farm to fork. Poult. Sci. 2015, 94, 1368–1378. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.-Z.; Yang, S.; Wu, G. Free radicals, antioxidants, and nutrition. Nutrition 2002, 18, 872–879. [Google Scholar] [CrossRef]
- Georgieva, N.V.; Koinarski, V.; Gadjeva, V. Antioxidant status during the course of Eimeria tenella infection in broiler chickens. Vet. J. 2006, 172, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, N.V.; Gabrashanska, M.; Koinarski, V.; Yaneva, Z. Zinc Supplementation against eimeria acervulina-induced oxidative damage in broiler chickens. Vet. Med. Int. 2011, 2011, 647124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Yin, Y.-L.; Li, D.; Woo Kim, S.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237. [Google Scholar] [CrossRef] [Green Version]
- Mishra, B.; Jha, R. Oxidative stress in the poultry gut: Potential challenges and interventions. Front. Vet. Sci. 2019, 6, 60. [Google Scholar] [CrossRef]
- Liang, M.; Wang, Z.; Li, H.; Cai, L.; Pan, J.; He, H.; Wu, Q.; Tang, Y.; Ma, J.; Yang, L. l-Arginine induces antioxidant response to prevent oxidative stress via stimulation of glutathione synthesis and activation of Nrf2 pathway. Food Chem. Toxicol. 2018, 115, 315–328. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione synthesis. Biochim. Et Biophys. Acta Gen. Subj. 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Li, F.; Mou, S.; Feng, J.; Liu, P.; Xu, L. Effects of dietary L-arginine on laying performance and anti-oxidant capacity of broiler breeder hens, eggs, and offspring during the late laying period. Poult. Sci. 2015, 94, 2938–2943. [Google Scholar] [CrossRef]
- Chen, Y.P.; Chen, X.; Zhang, H.; Zhou, Y.M. Effects of dietary concentrations of methionine on growth performance and oxidative status of broiler chickens with different hatching weight. Br. Poult. Sci. 2013, 54, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, J.; Kubińska, M.; Juśkiewicz, J.; Czech, A.; Ognik, K.; Zduńczyk, Z. Effect of different dietary methionine levels on the growth performance and tissue redox parameters of turkeys. Poult. Sci. 2017, 96, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Castro, F.L.S.; Tompkins, Y.H.; Pazdro, R.; Kim, W.K. The effects of total sulfur amino acids on the intestinal health status of broilers challenged with Eimeria spp. Poult. Sci. 2020, 99, 5027–5036. [Google Scholar] [CrossRef] [PubMed]
- Moskovitz, J.; Berlett, B.S.; Poston, J.M.; Stadtman, E.R. Methionine sulfoxide reductase in antioxidant defense. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; Volume 300, pp. 239–244. ISBN 0076-6879. [Google Scholar]
- Levine, R.; Moskovitz, J.; Stadtman, E. Oxidation of methionine in proteins: Roles in antioxidant defense and cellular regulation. Iubmb Life 2001, 50, 301–307. [Google Scholar] [CrossRef]
- Levine, R.L.; Mosoni, L.; Berlett, B.S.; Stadtman, E.R. Methionine residues as endogenous antioxidants in proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 15036–15040. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Jang, H.-J.; Cho, W.-Y.; Yeon, S.-J.; Lee, C.-H. In vitro antioxidant actions of sulfur-containing amino acids. Arab. J. Chem. 2020, 13, 1678–1684. [Google Scholar] [CrossRef]
- Lenis, Y.; Elmetwally, M.; Maldonado-Estrada, J.; Bazer, F. Physiological importance of polyamines. Zygote 2017, 25. [Google Scholar] [CrossRef]
- Rochell, S.J.; Helmbrecht, A.; Parsons, C.M.; Dilger, R.N. Interactive effects of dietary arginine and Eimeria acervulina infection on broiler growth performance and metabolism. Poult. Sci. 2017, 96, 659–666. [Google Scholar] [CrossRef]
- Allen, P.C.; Fetterer, R.H. Effect of Eimeria acervulina infections on plasma L-arginine. Poult. Sci. 2000, 79, 1414–1417. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Li, G. Nitric Oxide Production by macrophages stimulated with coccidia sporozoites, lipopolysaccharide, or interferon-γ, and its dynamic changes in sc and tk strains of chickens infected with eimeria tenella. Avian Dis. 2004, 48, 244–253. [Google Scholar] [CrossRef]
- Moinard, C.; Cynober, L.; de Bandt, J.-P. Polyamines: Metabolism and implications in human diseases. Clin. Nutr. 2005, 24, 184–197. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, F.L.d.S.; Kim, W.K. Secondary Functions of Arginine and Sulfur Amino Acids in Poultry Health: Review. Animals 2020, 10, 2106. https://doi.org/10.3390/ani10112106
Castro FLdS, Kim WK. Secondary Functions of Arginine and Sulfur Amino Acids in Poultry Health: Review. Animals. 2020; 10(11):2106. https://doi.org/10.3390/ani10112106
Chicago/Turabian StyleCastro, Fernanda Lima de Souza, and Woo K. Kim. 2020. "Secondary Functions of Arginine and Sulfur Amino Acids in Poultry Health: Review" Animals 10, no. 11: 2106. https://doi.org/10.3390/ani10112106