Understanding of Colistin Usage in Food Animals and Available Detection Techniques: A Review

 , ,
, , 
 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
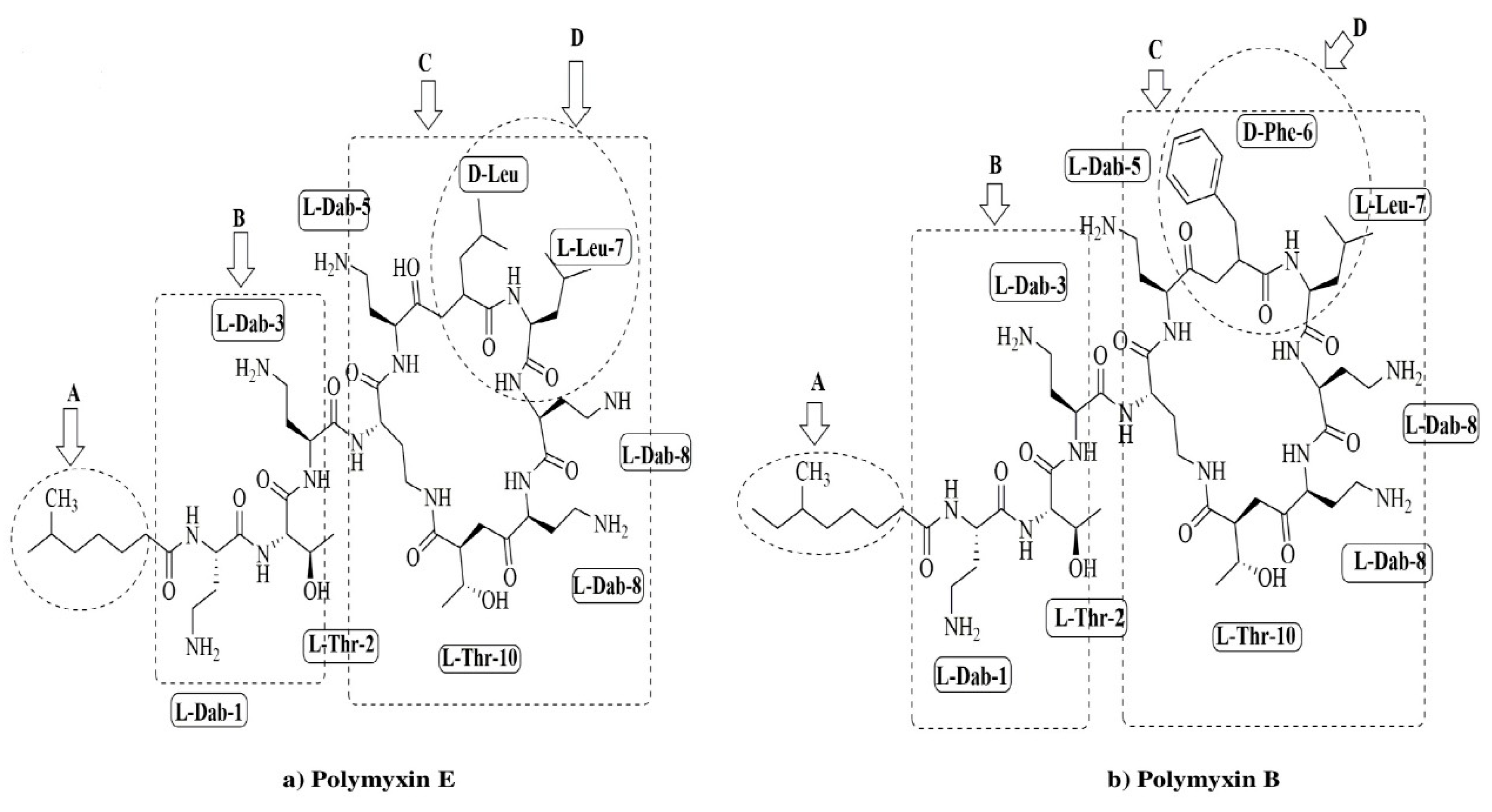
2. Colistin Use in Veterinary Medicine
3. Dosing Regimen of Colistin in Animals
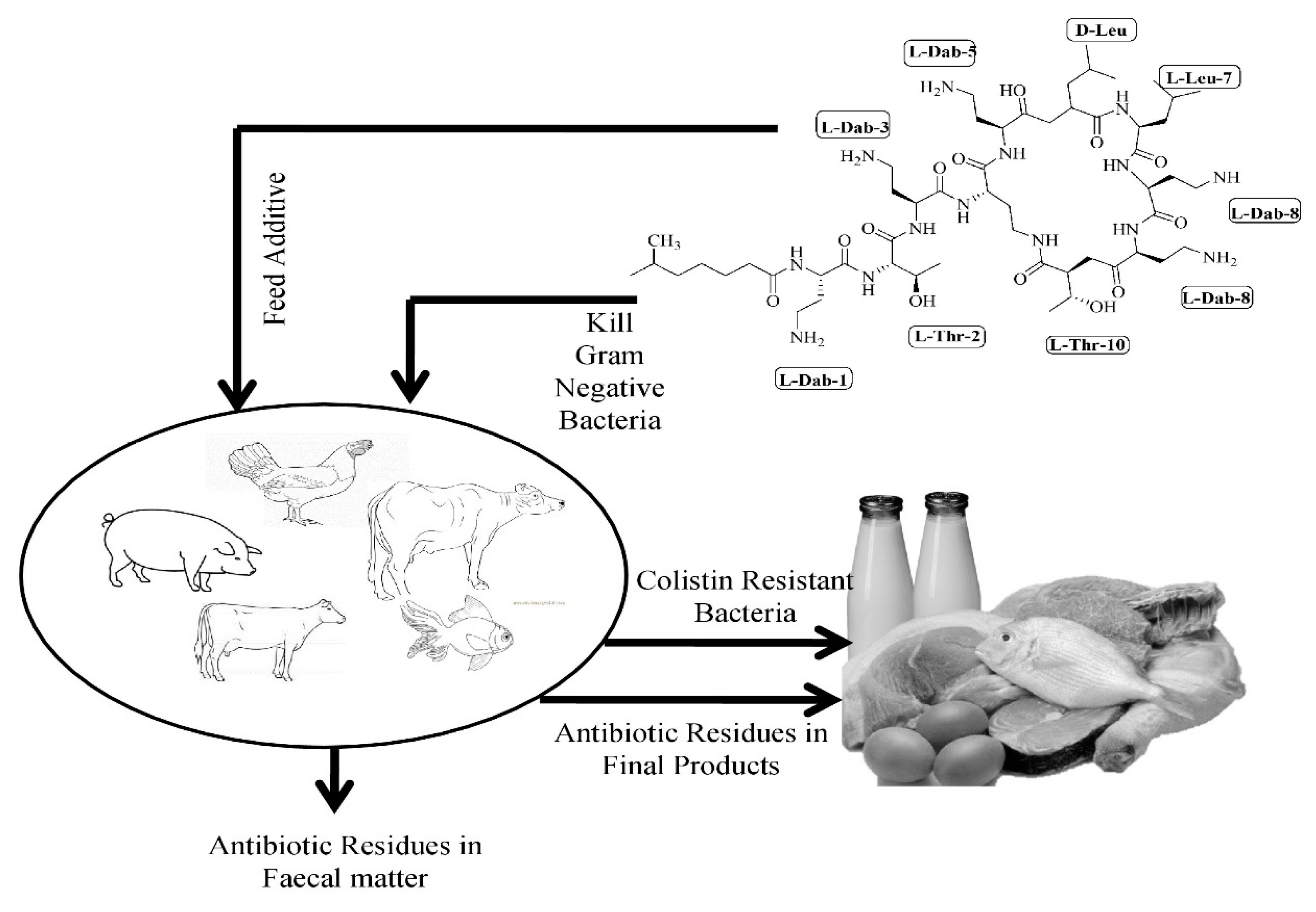
4. Negative Consequences of Colistin Consumption
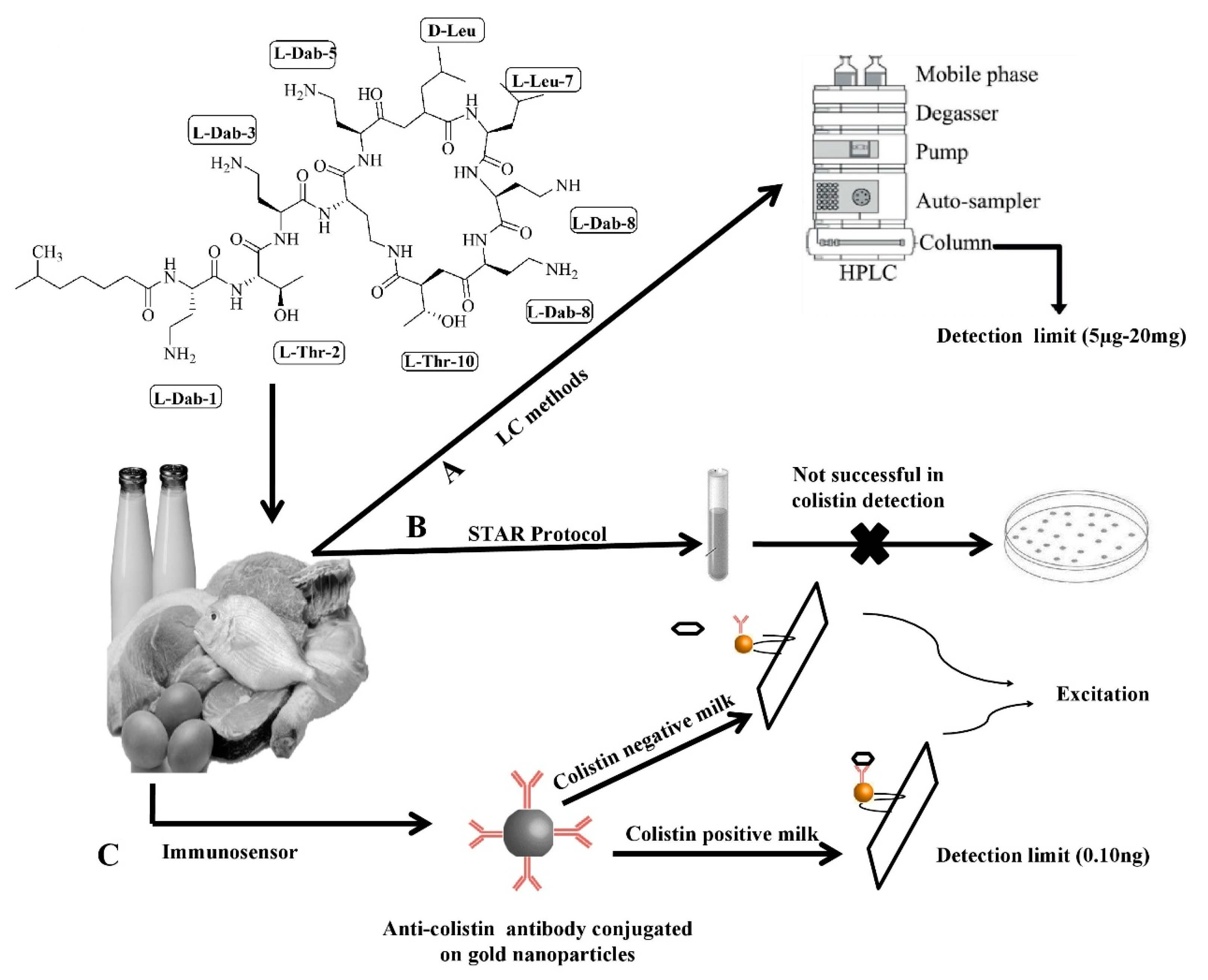
5. Routine Methods for Colistin Detection in Animals and Its Associated Challenges
6. Conclusions and Future Outlook
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Komura, S.; Kurahashi, K. Partial-purification and properties of L-2,4-diaminobutyric acid activating enzyme from a polymyxin-E producing organism. J. Biochem. 1979, 86, 1013–1021. [Google Scholar] [CrossRef]
- Falagas, M.E.; Kasiakou, S.K. Colistin: The revival of polymyxins for the management of multidrug-resistant gram-negative bacterial infections. Clin. Infect. Dis. 2005, 40, 1333–1341. [Google Scholar] [CrossRef] [Green Version]
- Newton, B.A. Properties and mode of action of the polymyxins. Bacteriol. Rev. 1956, 20, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Schindler, M.; Osborn, M.J. Interaction of divalent cations and polymyxin B with lipopolysaccharide. Biochemistry 1979, 18, 4425–4430. [Google Scholar] [CrossRef]
- Koch-Weser, J.; Sidel, V.W.; Federman, E.B.; Kanarek, P.; Finer, D.C.; Eaton, A.E. Adverse effects of sodium colistimethate. Manifestations and specific reaction rates during 317 courses of therapy. Ann. Intern. Med. 1970, 72, 857–868. [Google Scholar] [CrossRef]
- Spapen, H.; Jacobs, R.; Van-Gorp, V.; Troubleyn, J.; Honoré, P.M. Renal and neurological side effects of colistin in critically ill patients. Ann. IntensiveCare. 2011, 1, 14. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [Green Version]
- EFSA European Food Safety Authority; ECDC (European Centre for Disease Prevention and Control). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA J. 2017, 15, 5077. [Google Scholar]
- Aviv, G.; Tsyba, K.; Steck, N.; Salmon-Divon, M.; Cornelius, A.; Rahav, G.; Grassl, A.G.; Gal-Mor, O. A unique megaplasmid contributes to stress tolerance and pathogenicity of an emergent Salmonella enterica serovar Infantis strain. Environ. Microbiol. 2014, 16, 977–994. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.; Leekitcharoenphon, P.; Feltrin, F.; Alba, P.; Cordaro, G.; Iurescia, M.; Tolli, R.; D’Incau, M.; Staffolani, M.; Giannatale, E.D.; et al. Emergence of a clonal lineage of multidrug-resistant ESBL-producing Salmonella infantis transmitted from broilers and broiler meat to humans in Italy between 2011 and 2014. PLoS ONE 2015, 10, e0144802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hindermann, D.; Gopinath, G.; Chase, H.; Negrete, F.; Althaus, D.; Zurfluh, K.; Tall, B.D.; Stephan, R.; Nüesch-Inderbinen, M. Salmonella enterica serovar infantis from food and human infections, Switzerland, 2010–2015: Poultry-related multidrug resistant clones and an emerging ESBL producing clonal lineage. Front. Microbiol. 2017, 8, 1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, H.; Folster, J.P.; Hsu, C.H.; Chen, J.; Hoffmann, M.; Li, C.; Morales, C.; Tyson, G.H.; Mukherjee, S.; Brown, A.C.; et al. Comparative analysis of extended spectrum beta-lactamase CTX-M-65-producing Salmonella infantis isolates from humans, food animals, and retail chickens in the United States. Antimicrob. Agents Chemother. 2017, 61, e00488–e00517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano-Leon, A.; Garcia-Omil, C.; Dalama, J.; Rodriguez-Souto, R.; Martinez-Urtaza, J.; Gonzalez-Escalona, N. Detection of colistin resistance mcr-1 gene in Salmonella enterica serovar Rissen isolated from mussels, Spain, 2012–2016. Eurosurveillance 2019, 24, 1900200. [Google Scholar] [CrossRef] [PubMed]
- Mazutti, K.; Costa, L.B.; Nascimento, L.V.; Filho, T.F.; Beirão, B.C.B.; Júnior, P.C.M.; Maiorka, A. Effect of colistin and tylosin used as feed additives on the performance, diarrhea incidence, and immune response of nursery pigs. Semin. Cienc Agrar. 2016, 37, 1947–1962. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Tang, S.; Zhang, W.; Cui, X.; Zhang, Y.; Jin, Y.; Zhang, X.; Chen, Y. A surface-enhanced Raman scattering-based lateral flow immunosensor for colistin in raw milk. Sens. Actuat. B Chem. 2019, 282, 703–711. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Matsoso, P.; Pant, S.; Brower, C.; Røttingen, J.A.; Klugman, K.; Davies, S. Access to effective antimicrobials: A worldwide challenge. Lancet 2016, 387, 168–175. [Google Scholar] [CrossRef]
- Shen, Z.; Wang, Y.; Shen, Y.; Shen, J.; Wu, C. Early emergence of mcr-1 in Escherichia coli from food-producing animals. Lancet Infect. Dis. 2016, 16, 293. [Google Scholar] [CrossRef] [Green Version]
- Maron, D.F.; Smith, T.J.; Nachman, K.E. Restrictions on antimicrobial use in food animal production: An international regulatory and economic survey. Global Health. 2013, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- EMA/AMEG. Updated Advice on the Use of Colistin Products in Animals within the European Union: Development of Resistance and Possible Impact on Human and Animal Health. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/updated-advice-use-colistin-products-animals-within-european-union-development-resistance-possible_en-0.pdf (accessed on 9 August 2019).
- ECDC/EFSA/EMA. ECDC/EFSA/EMA Second Joint Report on the Integrated Analysis of the Consumption of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Bacteria from Humans and Food-Producing Animals. Available online: https://www.ema.europa.eu/en/documents/report/ecdc/efsa/ema-second-joint-report-integrated-analysis-consumption-antimicrobial-agents-occurrence_en.pdf (accessed on 9 August 2019).
- Friedlander, L.G.; Arnold, D. Colistin. Available online: http://www.fao.org/fileadmin/user_upload/vetdrug/docs/2-2006-colistin.pdf (accessed on 13 August 2019).
- Kempf, I.; Fleury, M.A.; Drider, D.; Bruneau, M.; Sanders, P.; Chauvin, C.; Madec, J.Y.; Jouy, E. What do we know about resistance to colistin in Enterobacteriaceae in avian and pig production in Europe? Int. J. Antimicrob. Agents 2013, 42, 379–383. [Google Scholar] [CrossRef]
- Catry, B.; Cavaleri, M.; Baptiste, K.; Grave, K.; Grien, K.; Holm, A.; Jukes, H.; Liebana, E.; Navas, A.L.; Mackay, D.; et al. Use of colistin-containing products within the European union and European economic area(EU/EEA): Development of resistance in animals and possible impact on human and animal health. Int. J. Antimicrob. Agents 2015, 46, 297–306. [Google Scholar] [CrossRef]
- Barbieri, N.; Nielsen, D.W.; Wannemuehler, Y.; Cavender, T.; Hussein, A.; Yan, S.G.; Nolan, L.K.; Logue, C.M. mcr-1 identified in avian pathogenic Escherichia coli (APEC). PLoS ONE 2017, 12, e0172997. [Google Scholar] [CrossRef] [Green Version]
- Botsoglou, N.A.; Fletouris, D.J. Antibacterial drugs. In Drug Residues in Foods; Fletouris, D., Botsoglou, N., Eds.; CRC Press: London, UK, 2000; pp. 27–103. [Google Scholar]
- Goetting, V.; Lee, K.A.; Tell, L.A. Pharmacokinetics of veterinary drugs in laying hens and residues in eggs: A review of the literature. J. Vet. Pharmacol. Ther. 2011, 34, 521–556. [Google Scholar] [CrossRef]
- Nolan, L.K.; Barnes, H.J.; Vaillancourt, J.P.; Abdul-Aziz, T.; Louge, C.M. Colibacillosis. In Diseases of Poultry; Swayne, D., Ed.; JohnWiley & Sons, Ltd.: Chichester, UK, 2007; pp. 751–805. [Google Scholar]
- Löhren, U.; Ricci, A.; Cummings, T.S. Guidelines for antimicrobial use in poultry. In Guide to Antimicrobial Use in Animals; Luca, G., Jensen, L.B., Kruse, H., Eds.; BlackwellPublishing, Ltd.: Oxford, UK, 2009; pp. 126–142. [Google Scholar]
- EU. On Pharmacologically Active Substances and Their Classification Regarding Maximum Residue Limits in Food Stuffs of Animal Origin. Available online: https://ec.europa.eu/health/sites/health/files/files/eudralex/vol-5/reg_2010_37/reg_2010_37_en.pdf (accessed on 7 August 2019).
- FSSAI. Food Safety and Standards (Contaminants, ToxinsandResidues). Available online: https://archive.fssai.gov.in/home/fss-legislation/fss-regulations.html (accessed on 7 August 2019).
- MAC. National Standard of the People’s Republic of China. Available online: http://www.agrichina.org/admin/kindeditor-4.1.2/attached/file/20160617/20160617183721_1681.pdf (accessed on 7 August 2019).
- PHR. Available online: https://www.elegislation.gov.hk/hk/cap139N (accessed on 7 August 2019).
- JFCRF. Maximum Residue Limitsin Animalsand Fishery Products. Available online: http://db.ffcr.or.jp/front/food_group_comp (accessed on 7 August 2019).
- Ziv, G.; Nouws, F.M.; Van Ginnekin, C.A.M. The pharmacokinetics and tissue levels of polymyxin B, colistin and gentamicin in calves. J. Vet. Pharmacol. Ther. 1982, 5, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Blood, D.C.; Radostits, O.M. Practical antimicrobial therapeutics: Polymyxin B and colistin. In Veterinary Medicine; Tindall, B., Ed.; London, UK, 1989; p. 151. Available online: http://www.fao.org/fileadmin/user_upload/vetdrug/docs/2-2006-colistin.pdf (accessed on 17 July 2019).
- Archimbault, P.; Boutier, C.; Fellous, R.; Muscat, G. Etude pharmacocinétique de la colistine chez les bovins. Rec Méd. Vét. 1980, 156, 621–626. [Google Scholar]
- Escoula, L.; Coste, M.; Larrieu, G. Biodisponibilité de l’association érythromycinecolistine chez les veaux. Ann. Rech. Vet. 1981, 12, 321–326. [Google Scholar] [PubMed]
- Ziv, G.; Sulman, F.G. Passage of polymyxin from serum into milk in Ewes. Am. J. Vet. Res. 1973, 34, 317–322. [Google Scholar]
- Al-Khayyat, A.A.; Aronson, A.L. Pharmacologic and toxicologic studies with the polymyxins.II.Comparative pharmacologic studies of the sulfate and methanesulfonate salts of polymyxin B and colistin in dogs. Chemotherapy 1973, 19, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Ouchi, M.; Koumi, J. Studies on the distribution of colistin sulfate in the body. Distribution and change with time in chickens and pigs by oral administration. Jpn. J. Antibiot. 1972, 25, 239–245. [Google Scholar]
- Terakado, S.; Azechi, H.; Omae, K.; Koyama, T.; Ninomiya, K.; Kashiwazaki, M. Distribution of colistin sulfate and changes with time in intestinal E.coli counts in pigs following oral administration. Seventy-Third Congr. Jpn. Soc. Vet. Med. 1972, 5–22. [Google Scholar]
- Lin, B.; Zhang, C.; Xiao, X. Toxicity, bioavailability and pharmacokinetics of a newly formulated colistin sulfate solution. J. Vet. Pharm. Ther. 2005, 28, 349–354. [Google Scholar] [CrossRef]
- Guyonnet, J.; Manco, B.; Baduel, L.; Kaltsatos, V.; Aliabadi, M.H.F.S.; Lees, P. Determination of a dosage regimen of colistin by pharmacokinetic/pharmacodynamic integration and modeling for treatment of G.I.T.disease in pigs. Res. Vet. Sci. 2010, 88, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Beaudry, F.; Thériault, W.; Bergeron, N.; Laurent-Lewandowski, S.; Fairbrother, J.M.; Letellier, A. Gastric stability and oral bioavailability of colistin sulfate in pigs challenged or not with Escherichia coli O149:F4(K88). Res. Vet. Sci. 2015, 102, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Beaudry, F.; Thériault, W.; Bergeron, N.; Beauchamp, G.; Laurent-Lewandowski, S.; Fairbrother, J.M.; Letellier, A. In vivo therapeutic efficacy and pharmacokinetics of colistin sulfate in an experimental model of enterotoxigenic Escherichia coli infection in weaned pigs. Vet. Res. 2016, 47, 58. [Google Scholar] [CrossRef] [Green Version]
- Rhouma, M.; Beaudry, F.; Letellier, A. Resistance to colistin: What is the fate for this antibiotic in pig production? Int. J. Antimicrob. Agents 2016, 48, 119–126. [Google Scholar] [CrossRef]
- Olaitan, A.O.; Morand, S.; Rolain, J.M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delannoy, S.; Le, D.L.; Jouy, E.; Fach, P.; Drider, D.; Kempf, I. Characterization of colistin-resistant Escherichia coli isolated from diseased pigs in France. Front. Microbiol. 2017, 8, 2278. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism mcr-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Biswas, S.; Brunel, J.M.; Dubus, J.C.; Reynaud-Gaubert, M.; Rolain, J.M. Colistin: An update on the antibiotic of the 21st century. Expert Rev. Anti-Infect Ther. 2012, 10, 917–934. [Google Scholar] [CrossRef]
- Xavier, B.B.; Lammens, C.; Ruhal, R.; Kumar-Sing, S.; Butaye, P.; Goossens, H.; Malhotra-Kumar, S. Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. Eurosurveillance 2016, 21, 30280. [Google Scholar] [CrossRef]
- AbuOun, M.; Stubberfield, E.J.; Duggett, N.A.; Kirchner, M.; Dormer, L.; Nunez-Garcia, J.; Randall, L.P.; Lemma, F.; Crook, D.W.; Teale, C.; et al. mcr-1 and mcr-2 variant genes identified in Moraxella species isolated from pigs in Great Britain from 2014 to 2015. J. Antimicrob. Chemother. 2017, 72, 2745–2749. [Google Scholar] [CrossRef] [Green Version]
- Borowiak, M.; Fischer, J.; Hammerl, J.A.; Hendriksen, R.S.; Szabo, I.; Malorny, B. Identification of a novel transposon-associated phosphoethanolamine transferase gene, mcr-5,conferring colistin resistance in d-tartrate fermenting Salmonella enterica subsp. Enterica serovar Paratyphi B. J. Antimicrob. Chemother. 2017, 72, 3317–3324. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.; Li, H.; Shen, Y.; Liu, Z.; Wang, S.; Shen, Z.; Zhang, R.; Walsh, T.R.; Shen, J.; Wang, Y. Novel plasmid-mediated colistin resistance gene mcr-3 in Escherichia coli. MBio 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teo, J.W.P.; Kalisvar, M.; Venkatachalam, I.; Ng, O.T.; Lin, R.T.P.; Octavia, S. mcr-3 and mcr-4 variants in carbapenemase-producing clinical Enterobacteriaceae do not confer phenotypic polymyxin resistance. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, Y.; Zhou, Y.; Li, J.; Yin, W.; Wang, S.; Zhang, S.; Shen, J.; Wang, Y. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg. Microbes Infect. 2018, 7, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.Q.; Li, Y.X.; Lei, C.W.; Zhang, A.Y.; Wang, H.N. Nove lplasmid-mediated colistin resistance gene mcr-7.1 in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2018, 73, 1791–1795. [Google Scholar] [CrossRef] [Green Version]
- Joshi, P.R.; Thummeepak, R.; Paudel, S.; Acharya, M.; Pradhan, S.; Banjara, M.R.; Leungtongkam, U.; Sitthisak, S. Molecular characterization of colistin-resistant Escherichia coli isolated from chickens: First report fromNepal. Microb. Drug Resist. 2019, 25, 846–854. [Google Scholar] [CrossRef]
- Yassin, A.K.; Zhang, J.; Wang, J.; Chen, L.; Kelly, P.; Butaye, P.; Lu, G.; Gong, J.; Li, M.; Wang, Y.; et al. Identification and characterization of mcr mediated colistin resistance in extra intestinal Escherichia coli from poultry and live stock in China. FEMS Microbiol. Lett. 2017, 364, fnx242. [Google Scholar] [CrossRef]
- Pishnian, Z.; Haeili, M.; Feizi, A. Prevalence and molecular determinants of colistin resistance among commensal Enterobacteriaceae isolated from poultry in northwest of Iran. Gut Pathog. 2019, 11, 2. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.T.; Nguyen, H.M.; Nguyen, C.V.; Nguyen, T.V.; Nguyenm, M.T.; Thai, H.Q.; Ho, M.H.; Thwaites, G.; Ngo, H.T.; Baker, S.; et al. Use of colistin and other critical antimicrobials on pig and chicken farms in Southern Vietnam and its association with resistance in commensal Escherichia col i bacteria. Appl. Environ. Microbiol. 2016, 82, 3227–3235. [Google Scholar] [CrossRef] [Green Version]
- Monte, D.F.; Mem, A.; Fernandes, M.R.; Cerdeira, L.; Esposito, F.; Galvão, J.A.; Franco, B.D.G.M.; Lincopan, N.; Landgraf, M. Chicken meat as a reservoir of colistin-resistant Escherichia coli strains carrying mcr-1 genes in South America. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Hasman, H.; Hammerum, A.M.; Hansen, F.; Hendriksen, R.S.; Olesen, B.; Agersø, Y.; Zankari, E.; Leekitcharoenphon, P.; Stegger, M.; Kass, R.S.; et al. Detection of mcr-1 encoding plasmid-mediated colistin resistant Escherichiacoli isolates from human bloodstream infection and imported chicken meat, Denmark 2015. Eurosurveillance 2015, 20, 30085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quesada, A.; Ugarte-Ruiz, M.; Iglesias, M.R.; Porrero, M.C.; Martínez, R.; Florez-Cuadrado, D.; Campos, M.J.; García, M.; Píriz, S.; Sáez, J.L.; et al. Detection of plasmid mediated colistin resistance(MCR-1) in Escherichia coli and Salmonella enterica isolated from poultry and swine in Spain. Res. Vet. Sci. 2016, 105, 134–135. [Google Scholar] [CrossRef] [PubMed]
- Donà, V.; Bernasconi, O.J.; Pires, J.; Collaud, A.; Overesch, G.; Ramette, A.; Perreten, V.; Endimiani, A. Heterogens genetic location of mcr-1 in colistin-resistant Escherichia coli isolates from humans and retail chicken meat in Switzerland: Emergence of mcr-1-carrying Inc K2 plasmids. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwaiger, K.; Huther, S.; Hölzel, C.; Kämpf, P.; Bauer, J. Prevalence of antibiotic-resistant Enterobacteriaceae isolated from chicken and pork meat purchased at the slaughter house and at retail in Bavaria, Germany. Int. J. Food Microbiol. 2012, 154, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Kusumoto, M.; Ogura, Y.; Gotoh, Y.; Iwata, T.; Hayashi, T.; Akiba, M. Colistin-resistant mcr-1–positive pathogenic Escherichia coli in swine, Japan,2007–2014. Emerg. Infect. Dis. 2016, 22, 1315–1317. [Google Scholar] [CrossRef] [Green Version]
- Benameura, Q.; Guemourb, D.; Hammoudic, A.; Aoudiad, H.; Aggad, H.; Humblet, M.F.; Saegerman, C. Antimicrobial resistance of Escherichia coli isolated from chickens in West of Algeria. Int. J. Sci. Basic Appl. Res. 2014, 13, 366–370. [Google Scholar]
- Chiou, C.S.; Chen, Y.T.; Wang, Y.W.; Liu, Y.Y.; Kuo, H.C.; Tu, Y.H.; Lin, A.C.; Liao, Y.S.; Hong, Y.P. Dissemination of mcr-1-carrying plasmids among colistin-resistant Salmonella strains from humans and food-producing animals in Taiwan. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Anjum, M.F.; Duggett, N.A.; AbuOun, M.; Randall, L.; Nunez-Garcia, J.; Ellis, R.J.; Rogers, J.; Horton, R.; Brena, C.; Williamson, S.; et al. Colistin resistance in Salmonella and Escherichia coli isolates from a pig farm in Great Britain. J. Antimicrob. Chemother. 2016, 71, 2306–2313. [Google Scholar] [CrossRef] [Green Version]
- Enne, V.I.; Cassar, C.; Sprigings, K.; Woodward, M.J.; Bennett, P.M. A high prevalence of antimicrobial resistant Escherichia coli isolated from pigs and a low prevalence of antimicrobial resistant E.coli from cattle and sheep in Great Britain at slaughter. FEMS Microbiol. Lett. 2008, 278, 193–1999. [Google Scholar] [CrossRef] [Green Version]
- Curcio, L.; Luppi, A.; Bonilauri, P.; Gherpelli, Y.; Pezzotti, G.; Pesciaroli, M.; Magistrali, C.F. Detection of the colistin resistance gene mcr-1 in pathogenic Escherichia coli from pigs affected by post-weaning diarrhoea in Italy. J. Glob. Antimicrob. Resist. 2017, 10, 80–83. [Google Scholar] [CrossRef]
- Magwira, C.A.; Gashe, B.A.; Collison, E.K. Prevalence and antibiotic resistance profiles of Escherichia coli O157:H7 in beef products from retail outlets in Gaborone, Botswana. J. FoodProt. 2005, 68, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Waghamare, R.N.; Paturkar, A.M.; Vaidya, V.M.; Zende, R.J.; Dubal, Z.N.; Dwivedi, A.; Gaikwad, R.V. Phenotypic and genotypic drug resistance profile of Salmonella serovars isolated from poultry farm and processing units located in and around Mumbai city, India. Vet. World. 2018, 11, 1682–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mir, I.A.; Kashyap, S.K.; Maherchandani, S. Isolation, serotype diversity and antibiogram of Salmonella enterica isolated from different species of poultry in India. Asian Pac. J. Trop. Biomed. 2015, 5, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Giani, T.; Arena, F.; Vaggelli, G.; Conte, V.; Chiarelli, A.; Angelis, L.H.D.; Fornaini, R.; Grazzini, M.; Niccolini, F.; Pecile, P.; et al. Large nosocomial outbreak of colistin-resistant, carbapenemase-producing Klebsiella pneumoniae traced to clonal expansion of an mgrB deletion mutant. J. Clin. Microbiol. 2015, 53, 3341–3344. [Google Scholar] [CrossRef] [Green Version]
- Landman, D.; Georgescu, C.; Martin, D.A.; Quale, J. Polymyxins revisited. Clin. Microbiol. Rev. 2008, 21, 449–465. [Google Scholar] [CrossRef] [Green Version]
- Lim, L.M.; Ly, N.; Anderson, D.; Yang, J.C.; Macander, L.; Jarkowski, A.; Forrest, A.; Bulitta, J.B.; Tsuji, B.T. Resurgence of colistin: A review of resistance, toxicity, pharmacodynamics, and dosing. Pharmacotherapy 2010, 30, 1279–1291. [Google Scholar] [CrossRef]
- Yahav, D.; Farbman, L.; Leibovici, L.; Paul, M. Colistin: New lessons on an old antibiotic. Clin. Microbiol. Infect. 2012, 18, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.; Walsh, T.R. A colistin crisis in India. Lancet Infect. Dis. 2018, 18, 256–257. [Google Scholar] [CrossRef]
- Livemint. Govt May Ban Antibiotic Colistin Used to Fatten Chicken. Available online: https://www.livemint.com/Industry/yt5eE5hqMLYP1px2d63Q1K/Govt-may-ban-antibiotic-colistin-used-to-fatten-chicken.html (accessed on 20 June 2019).
- TOI. Tolerance Limits’ to Be Fixed by Food Regulator for Presence of Antibiotics in Animal, Foods. Available online: https://www.fssai.gov.in/upload/media/FSSAI_News_AntiBiotics_TOI_01_08_2018.pdf (accessed on 12 June 2019).
- FSSAI. Direction under Section 16(5)Read with 18(2)(d)of Food Safety and Standard Act, 2006 Regarding Operationalisation of Draft Food Safety and Standard (Contaminants, Toxins and Residues) Amendment Regulations. 2019. Available online: https://www.fssai.gov.in/upload/advisories/2019/08/5d4c042779d77Direction_Colistin_Ban_FSSAI_08_08_2019.pdf (accessed on 8 September 2019).
- Kunin, C.M. Binding of antibiotics to tissue homogenates. J. Infect. Dis. 1970, 121, 55–64. [Google Scholar] [CrossRef]
- Sin, D.W.; Ho, C.; Wong, Y.C.; Ho, S.K.; Ip, A.C.B. Analysis of major components of residual bacitracin and colistin in food samples byliquid chromatography tandem mass spectrometry. Anal. Chim. Acta. 2005, 535, 23–31. [Google Scholar] [CrossRef]
- Xu, I.; Tian, X.; Ren, C.; Huang, H.; Zhang, X.; Gong, X.; Liu, H.; Yu, Z.; Zhang, L. Analysis of colistin A and B in fishery products by ultra performance liquid chromatography with positive electro spray ionization tandem mass spectrometry. J. Chromatogr. B 2012, 899, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Morovján, G.; CsoKán, P.P.; Németh-Konda, L. HPLC determination of colistin and aminoglycoside antibiotics in feeds by post-column derivatization and fluorescence detection. Chromatographia 1998, 48, 32–36. [Google Scholar] [CrossRef]
- Cancho-Grande, B.; Rodríguez-Comesaña, M.; Simal-Gándara, J. Simple HPLC determination of colistin in medicated feeds by pre-column derivatization and fluorescence detection. Chromatographia 2001, 54, 481–484. [Google Scholar] [CrossRef]
- Decolin, D.; Leroy, P.; Nicolas, A.; Archimbault, P. Hyphenated liquid chromatographic method for the determination of colistin residues in bovine tissues. J. Chromatogr. Sci. 1997, 35, 557–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Li, X.; Zheng, K.; Ke, Y.; Wang, Y.; Wang, L.; Yu, F.; Xia, X. Determination of colistin in animal tissues, egg, milk, and feed by ultra-highperformance liquid chromatography-tandem mass spectrometry. Food Chem. 2018, 248, 166–172. [Google Scholar] [CrossRef]
- Meersche, T.V.D.; Pamel, E.V.; Poucke, C.V.; Herman, L.; Heyndrickx, M.; Rasschaert, G.; Daeseleire, E. Development, validation and application of an ultra high performance liquid chromatographic-tandem mass spectrometric method for the simultaneous detection and quantification of five different classes of veterinary antibiotics in swine manure. J. Chromatogr. A. 2016, 1429, 248–257. [Google Scholar] [CrossRef]
- Tao, Y.; Xie, S.; Zhu, Y.; Chen, D.; Pan, Y.; Wang, X.; Liu, Z.; Huang, L.; Peng, D.; Yuan, Z. Analysis of major components of bacitracin, colistin and virginiamycin in feed using matrix solid-phase dispersion extraction by liquid chromatography-electrospray ionization tandem mass spectrometry. J. Chromatogr. Sci. 2017, 56, 281–291. [Google Scholar] [CrossRef]
- Song, X.; Huang, Q.; Zhang, Y.; Zhang, M.; Xie, J.; He, L. Rapid multi residue analysis of authorized/banned cyclo poly peptide antibiotics in feed by liquid chromatography-tandem mass spectrometry based on dispersives olid-phase extraction. J. Pharm. Biomed. Anal. 2019, 170, 234–242. [Google Scholar] [CrossRef]
- Boison, J.O.; Lee, S.; Matus, J. A multi-residue method for the determination of seven polypeptide drug residues in chicken muscle tissues byLC-MS/MS. Anal. Bioanal. Chem. 2015, 406, 4065–4078. [Google Scholar] [CrossRef]
- Saluti, G.; Diamanti, I.; Giusepponi, D.; Pucciarini, L.; Rossi, R.; Moretti, S.; Sardella, R.; Galarini, R. Simultaneous determination of aminoglycosides and colistins in food. Food Chem. 2018, 266, 9–16. [Google Scholar] [CrossRef]
- Wan, E.C.; Ho, C.; Sin, D.W.; Wong, Y.C. Detection of residual bacitracin A, colistin A, and colist in B in milk and animal tissues by liquid chromatography tandem mass spectrometry. Anal. Bioanal. Chem. 2006, 385, 181–188. [Google Scholar] [CrossRef]
- Kaufmann, A.; Widmer, M. Quantitative analysis of polypeptide antibiotic residues in a variety of food matrices by liquid chromatography coupled to tandem mass spectrometry. Anal. Chim. Acta 2013, 797, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Gaudin, V.; Maris, P.; Fuselier, R.; Ribouchon, J.L.; Cadieu, N.; Rault, A. Validation of a microbiological method: The STAR protocol, a five-plate test, for the screening of antibiotic residues in milk. Food Addit. Contam. 2004, 21, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Gaudin, V.; Hedou, C.; Rault, A.; Verdon, E. Validation of a five plate test, the STAR protocol, for the screening of antibiotic residues in muscle from different animal species according to European decision 2002/657/EC. Food Addit. Contam. 2010, 27, 935–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aarestrup, F.M.; Wegener, H.C.; Collignon, P. Resistance in bacteria of the foodc hain: Epidemiology and control strategies. Expert Rev. Anti-Infect Ther. 2008, 6, 733–750. [Google Scholar] [CrossRef]
- Rhouma, M.; Beaudry, F.; Thériault, W.; Letellier, A. Colistin in pig production: Chemistry, mechanism of antibacterial action, microbial resistance emergence, and one health perspectives. Front. Microbiol. 2016, 7, 1789. [Google Scholar] [CrossRef]
- Gupta, S.; Kaushal, A.; Kumar, A.; Kumar, D. Ultrasensitive transglutaminase based nanosensor for early detection of celiac disease in human. Int. J. Biol. Macromol. 2017, 5, 905–911. [Google Scholar] [CrossRef]
- Kaushal, A.; Singh, S.; Kumar, A.; Kumar, D. Nano-Au/cMWCNT modified speB gene specific amperometric sensor for rapidly detecting Streptococcus pyogenes causing rheumatic heart disease. Indian J. Microbiol. 2017, 57, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Nagraik, R.; Kaushal, A.; Gupta, S.; Dhar, P.; Sethi, S.; Kumar, D. Optimized DNA-based bioassay for Leptospira interrogans detection: A novel platform for leptospirosis diagnosis. 3 Biotech 2019, 9, 284. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |




{kind=link}
{kind=link}
{kind=link}
| Animal Species | Target Tissue | MRLs (Per Kg) | Reference |
|---|---|---|---|
| All food producing animal spp. | Fat, muscle, liver | 150 µg, 150µg, 150 µg | [29] |
| All food producing animal spp. | Kidney | 200 µg | [29] |
| All food producing animal spp. | Milk | 50 µg | [29] |
| All food producing animal spp. | Eggs | 300 µg | [29] |
| Cattle, sheep’s | Fat, muscle, kidney, liver, milk | 150 µg, 150 µg, 200 µg, 150 µg, 50 µg | [30] |
| Pig, goat, rabbit | Fat, muscle, liver, kidney | 150 µg, 150 µg, 150 µg, 200 µg | [30] |
| Chicken | Fat, liver, kidney, eggs | 150 µg, 150 µg, 200 µg, 300 µg | [30] |
| Turkey | Fat, muscle, liver, kidney | 150 µg, 150 µg, 150 µg, 200 µg | [30] |
| Cattle, lamb | Milk | 50 µg | [31] |
| Cattle, lamb, swine, chicken, rabbit | Fat, muscle, liver, kidney | 150 µg, 150 µg, 150 µg, 200 µg | [31] |
| Bovine | Muscle, liver, kidney | 150 µg, 150 µg, 200 µg | [32] |
| Porcine | Liver, kidney | 150 µg, 200 µg | [32] |
| Poultry | Muscle, liver, kidney | 150 µg, 150 µg, 200 µg | [32] |
| Pig, cattle, chicken | Muscle, fat, liver | 150 µg | [33] |
| Cattle | Milk | 50 µg | [33] |
| Pig, cattle, chicken | Kidney | 200 µg | [33] |
| Country | Type of Animals | Type of Samples | Sample Size | Type of Microbes | Detection Basis | Reference |
|---|---|---|---|---|---|---|
| Nepal | Healthy chickens | Cloacal swabs | 324 | Escherichia coli | mcr-1 | [58] |
| China | Healthy chickens | Lung, spleen | 644 | Escherichia coli | mcr-1 | [59] |
| Pigs | Liver | 113 | ||||
| Cows | Milk | 61 | ||||
| Ducks | Liver | 44 | ||||
| Iran | Healthy broilers | Cloacal swabs | 503 | Klebsiella pneumoniae | mcr-1, 2, 3, 4 | [60] |
| Dead broilers | 388 | |||||
| Dead lying hens | 30 | |||||
| Dead turkeys | 23 | |||||
| Vietnam | Healthy chickens | Cloacal swabs | NS | Escherichia coli | mcr-1 | [61] |
| Pigs | ||||||
| Brazil | Healthy chickens | Breast | 20 | Escherichia coli | mcr-1 | [62] |
| Thigh | 20 | |||||
| Liver | 1 | |||||
| Denmark | Chicken meat | ND | NS | Escherichia coli | mcr-1 | [63] |
| Spain | Swine | Lymph node | NS | Salmonella enterica, Escherichia coli | mcr-1 | [64] |
| Faeces | ||||||
| Turkey | Faeces | NS | Escherichia coli | |||
| Switzerland | Chicken meat | ND | 6 | Escherichia coli | mcr-1 | [65] |
| Germany | Healthy chicken | Drumsticks | 500 | Cirobacter freundii, Klebsiella oxytoca, Pantoea agglomerans | Disc diffusion | [66] |
| Pork | Belly | 500 | Escherichia coli, Klebsiella oxytoca | |||
| Japan | Diseased swine | ND | NS | Escherichia coli | mcr-1 | [67] |
| Algeria | Healthy chickens | ND | NS | Escherichia coli | Disc diffusion | [68] |
| Taiwan | Diseased Chickens | ND | 450 | Salmonella spp. | mcr-1 | [69] |
| Pigs | 279 | |||||
| Ducks | 206 | |||||
| Turkeys | 170 | |||||
| Geese | 88 | |||||
| Great Britain | Diseased pigs | Small intestine | 3 | Escherichia coli, Salmonella typhimurium | mcr-1 | [70] |
| Great Britain | Healthy Pigs | Cecums | 2509 | Escherichia coli | Disc diffusion | [71] |
| Cattle | Distal rectums | 891 | ||||
| Sheep | Distal rectums | 973 | ||||
| Italy | Diseased pigs | Rectal swabs, faeces, intestines | NS | Escherichia coli | Disc diffusion, mcr-1 | [72] |
| Great Britain | Healthy pigs | Cecal contents | NS | Moraxella spp. | MIC and mcr-1,2 | [52] |
| France | Diseased pigs | Intestinal | 63 | Escherichia coli | Disc diffusion | [48] |
| Septicemia | 2 | |||||
| Nervous system | 1 | |||||
| Lymph node | 1 | |||||
| Urine | 1 | |||||
| Botswana | Beef | Meat cubes | 134 | Escherichia coli O157: H7 | Disc diffusion | [73] |
| Minced meat | 133 | |||||
| Fresh sausages | 133 | |||||
| India | Poultry | ND | NS | Salmonella spp. | Disc diffusion | [74] |
| India | Chickens | Faecal, cecal | 434 | Salmonella enterica | Disc diffusion | [75] |
| Ducks | Faecal | 38 | ||||
| Emus | Faecal | 35 |
| Country | Sample | Method Used | Chromatography Conditions Used | Detection Limit | Reference | |||
|---|---|---|---|---|---|---|---|---|
| Model | Column | Solvent | Flow Rate | |||||
| China | Spiked bovine milk | HPLC–MS/MS | An HPLC (Hewlett-Packard HP 1100 series, Rockville, MD, USA) integrated system consisting of a 100-well auto-sampler, a 100 µL sample loop, a degasser, a quaternary pump and a thermostated column oven set at 25 °C was used | Chromatographic separation was performed in a 250 mm × 2.1 mm, 5 µm Alltima C18 separation column (Alltech, Deerfield, MA, USA) and a corresponding C18 guard column (7.5 mm × 4.6 mm) | Mobile phase A: 0.1% formic acid in acetonitrile and mobile phase B: saturated ammonium formate:formic acid:acetonitrile:water (1:5:50:950, v/v/v/v) | Flow rate of 0.2 mL min−1 under a gradient elution program comprised of two mobile phases | 50 µg/Kg | [85] |
| China | Spiked fishery products | UPLC–MS/MS | A UPLC–MS/MS system comprised an Acquity UPLC system connected online with a Quattro Premier tandem mass spectrometer (Waters, Milford, MA, USA) | The column used was an ACQUITYTM BEH C18 reversed phase column (2.1 mm × 100 mm, 1.7 µm particle size) maintained at 40 °C | Mobile phase was 0.2% formic acid in acetonitrile and 0.2% formic acid in water | Flow rate and temperature of the drying gas (N2) were 750 L h−1 and 350 °C, respectively. The cone gas flow (N2) was 50 L h−1 | 10 µg/Kg (colistin A), 40 µg/Kg (colistin B) | [86] |
| Hungary | Spiked pig feeds | HPLC–fluorescence detector | JASCO PU-980 high pressure pump (JASCO, Kyoto, Japan) | A TSK ODS 120T column (150 × 4.6 mmID, 5 µm) was used with an injection volume of 25 µL | Mobile phase was 22:78 v/v acetonitrile–50 mM sodium sulfate, 20 mM orthophosphoric acid, 25 mM triethylamine | Flow rate of the mobile phase and post-column reagent were 1.5 and 1.0 mL min−1, respectively | 20 mg/Kg | [87] |
| Spain | Spiked animal feeds | HPLC–fluorescence detector | Thermo HPLC system equipped with a P200 gradient pump | Analytical column (150 × 4.6 mm i.d.) used was packed with Ultracarb 5 µm ODS 30%C. Guard columns (50 × 4.6 mm i.d.) were packed with dry 40 µm Pelliguard LC-18 | Mobile phases with methanol and acetonitile | Flow of 1.5 mL | 5 mg/Kg | [88] |
| France | Spiked bovine milk and tissues (muscle, liver, kidney, fat) | HPLC–MS | The HPLC system consisted of a solvent delivery pump (model P2000, Thermo Separation Products, Les Ulis, France), an injection valve (model 7725i, Rheodyne, Cotati, CA, USA) | An analytical column (125 × 4 mm i.d.) pre-packed with 5 μmNucleosil C18 (Macherey-Nagel, Düren, Germany) | Mobile phase was acetonitrile and a 0.035 M triethylamine solution adjusted to pH 2.5 with phosphoric acid and mixed in 17:83 (v/v) proportions | The flow rate was 1.5 mL/min | 25 µg/L (milk), 100 µg/Kg (tissues) | [89] |
| China | Swine liver, chicken eggs, feed, swine muscles, chicken muscles, bovine muscles, sheep muscles, bovine raw milk | UHPLC–MS/MS | An Acquity ultra-performance liquid chromatography system (Waters, Milford, MA, USA) | An Acquity BEH C18 column (50 mm × 2.1 mm i.d., 1.7 μm particle size) (Waters, Milford, MA, USA) | Mobile phases comprised of 0.5% formic acid in water (solvent A) and 0.5% formic acid in acetonitrile (solvent B) | Flow rate was 0.4 mL/min with the following gradient program: 0–0.5 min, 95% A; 0.5–3.0 min, 95–50% A; 3.0–4.0 min, 50–5% A; 4.0–4.1 min, 5–95% A; 4.1–5.5 min, 95% A | 5–30 µg/Kg | [90] |
| Belgium | Spiked swine manure | UHPLC–MS/MS | An Acquity UPLC H-class system (Waters, Milford, MA, USA) | Reversed-phase Kinetex C18 column (100 mm × 2.1 mm i.d., 1.7 µm) with a SecurityGuard Ultra guard cartridge system (Phenomenex, Utrecht, The Netherlands) | The elution was performed gradually with changing amounts of H2O/MeCN (95/5) + 0.5% FA + 0.1% ammonium formate (solvent A) and MeCN + 0.1% FA (solvent B). The gradient (15 min) was initiated with 95% of solvent A (0–1 min), followed by a linear decrease of A to 75% (1–3 min). From min 3–5, there was a linear decrease of solvent A to 0% and this was held until min 7. Re-equilibration of the gradient at 95% A was held from min 7–15 | Flow at 400 µL/min | 20.2 µg/Kg (colistin A), 15 µg/Kg (colistin B) | [91] |
| China | Spiked swine and poultry feeds | UHPLC–MS/MS | LC–MS/MS system (Thermo Electron Corp., Wyman, Waltham, MA, USA) consisting of a Finnigan Surveyor Plus system with an online degasser, a Surveyor autosampler and a TSQ Quantum triple quadrupole mass spectrometer equipped with an electrospray interface operating in the positive mode (ESI+) | Separation was performed on 150 mm × 2.1 mm, 5 μm Hypersil Gold C18 analytical columns (Thermo Electron Corporation, Waltham, MA, USA) | Mobile phase A consist formic acid in water and mobile phase B formic acid in ACN | Flow-rate of 0.2 mL min−1 | 27.5 µg/Kg (colistin A), 25.7 µg/Kg (colistin B) | [92] |
| China | Spiked piglet premix, pig feed additive, poultry complete feed, pig complete feed and fattening pig premix | UHPLC–MS/MS | Shimadzu liquid chromatography system (Shimadzu, Kyoto, Japan) | Separations were carried out on a Phenomenex Kinetex Biphenyl column (50 mm × 2.1 mm i.d., 2.6 µm particle size, Phenomenex, Torrance, CA, USA) | Mobile phase consisted of 0.1% FA in ACN solution (A) and 0.1% FA in water solution (B) with the following gradient elution program: 0 min, 6% A; 2 min, 6% A; 5 min, 40% A; 14 min, 70% A; 14.1 min, 6% A; 18 min, 6% A | Flow rate of 0.2 mL/min. | 5–20 µg/Kg (colistin A), and (colistin B) | [93] |
| Canada | Spiked chicken muscle | UPLC–MS/MS | Waters Acquity UPLC interfaced to a Waters Micromass triple quadrupole Premier mass spectrometer equipped with an ESI source and controlled by MassLynx 4.1 software(Waters, Milford, MA, USA) | Poroshell 120, 100 × 2.1 mm id, 2.7 μm (Agilent Technologies, Mississauga, ON Canada) | Mobile phase A (0.1% formic acid in water)and mobile phase B (methanol) | Flow rate of 0.40 mL/min | 39 µg/Kg (colistin A), 50 µg/Kg (colistin B) | [94] |
| Italy | Spiked bovine milk, meat | HPLC–MS | Thermo Ultimate 3000 High Performance Liquid Chromatography system (Thermo Scientific, San Jose, CA, USA) | InfinityLab Poroshell 120 HILIC column (100 × 2.1 mm; 2.7 μm, Agilent Technologies, Santa Clara, CA, USA) connected with the InfinityLab Poroshell 120 HILIC guard column (5 × 2.1 mm, 2.7 μm) | Eluent A was an aqueous solution containing 1% (v/v) formic acid (FA) and 1 mM ammonium formate (AF), eluent B was acetonitrile. The gradient was initiated with 20% eluent A for 2 min, continued with linear increase to 35% A in 5 min. In 1 min eluent A increased to 95% and this condition was maintained for 7 min. The system returned to 20% B in 0.1 min and was re-equilibrated for 4 min (run time: 17 min) | Flow rate was 0.25 mL min−1 | 33 µg/Kg | [95] |
| Hong Kong | Spiked bovine milk and tissues | HPLC–MS/MS | An integrated HPLC system (Hewlett–Packard HP 1100series, Rockville, MD, USA) consisting of a 100-well autosampler, a degasser, two-channel binary pump, and atemperature control oven (set at 25 °C), and interfaced with a TSQ Quantum Discovery mass spectrometer (Thermo-Finnigan, San Jose, CA, USA) | 150 mm×2.1 mm, 5 μm Phenomenex Luna C18 analytical column (Torrance, CA, USA) connected to a 7.5 mm × 4.6 mm Alltech Alltima C18 guard column (Deerfield, IL, USA) | Mobile phases, which were comprised of a mixture of (A) 0.1% formic acid in water and (B) 0.1% formic acid in acetonitrile, were delivered under a gradient elution program (0–4 min: 95% A, 5% B; 4–8 min: 30% A, 70% B and held for 4 min; 12 min: 95% A, 5% B and held for 3 min to restore initial conditions before the next injection | Flow-rate of 0.25 mL min−1 | 1–16 µg/Kg (colistin A), 6–14 µg/Kg (colistin B) | [96] |
| Switzerland | Spiked bovine liver, kidney, muscle, egg, milk | UHPLC–MS/MS | Acquity system (sample and solvent manager) from Waters (Millford, MA, USA) | Kinetex C18, 2.1 × 150 mm × 2.6 µm column with an installed pre-filter (Krud-katcher), both from Phenomenex (Torrance CA, USA) | Mobile phase A: 50 mL acetonitrile, 3 mL of formic acid and 0.1 mL of trifluoroacetic acid were transferred into a 1000 mL volumetricflask and diluted to volume with purified water; Mobile phase B: 50 mL of purified water, 3 mL of formic acidand 0.1 mL of trifluoroacetic acid were transferred into a 1000-mLvolumetric flask and diluted to volume with ACN | Linear gradient was used: 0–2 min with 8% B and flow0.4 mL min−1, 2–7 min with 8–20% B, 7–8 min with 20–30% B, 8–11 min with 30–100% B, 11–11.1 min with 100% B and flow 0.4–0.8 mL min−1, 11.1–12.5 min with 100%, 12.5–12.51 min with 100–8% B and flow 0.8–0.4 mL min−1. 12.51–14 min with 8% B and flow 0.4 mL min−1 | Muscle 15 µg/Kg (colistin A), 30 µg/Kg (colistin B); kidney 30 µg/Kg (colistin A), 30 µg/Kg (colistin B); liver 30 µg/Kg (colistin A), 30 µg/Kg (colistin B); egg 20 µg/Kg (colistin A), 30 µg/Kg (colistin B); milk 20 µg/Kg (colistin A), 40 µg/Kg (colistin B); | [97] |
| France | Spiked milk | Disc diffusion method (STAR protocol) | NA | NA | NA | NA | 1 mg/L | [98] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, H.; Chen, B.-H.; Kuca, K.; Nepovimova, E.; Kaushal, A.; Nagraik, R.; Bhatia, S.K.; Dhanjal, D.S.; Kumar, V.; Kumar, A.; et al. Understanding of Colistin Usage in Food Animals and Available Detection Techniques: A Review. Animals 2020, 10, 1892. https://doi.org/10.3390/ani10101892
Kumar H, Chen B-H, Kuca K, Nepovimova E, Kaushal A, Nagraik R, Bhatia SK, Dhanjal DS, Kumar V, Kumar A, et al. Understanding of Colistin Usage in Food Animals and Available Detection Techniques: A Review. Animals. 2020; 10(10):1892. https://doi.org/10.3390/ani10101892
Chicago/Turabian StyleKumar, Harsh, Bing-Huei Chen, Kamil Kuca, Eugenie Nepovimova, Ankur Kaushal, Rupak Nagraik, Shashi Kant Bhatia, Daljeet Singh Dhanjal, Vinod Kumar, Anil Kumar, and et al. 2020. "Understanding of Colistin Usage in Food Animals and Available Detection Techniques: A Review" Animals 10, no. 10: 1892. https://doi.org/10.3390/ani10101892