
Age and Giardia intestinalis Infection Impact Canine Gut Microbiota

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dogs
2.2. Giardia Cysts Count and Genotyping
2.3. Quantification of Fecal IgA and Calprotectin
2.4. DNA Extraction
2.5. Primer Design and Library Preparation
2.6. Illumina Sequencing
2.7. Statistical Analyses
3. Results
3.1. G. intestinalis Infection Is Chronic in Puppies
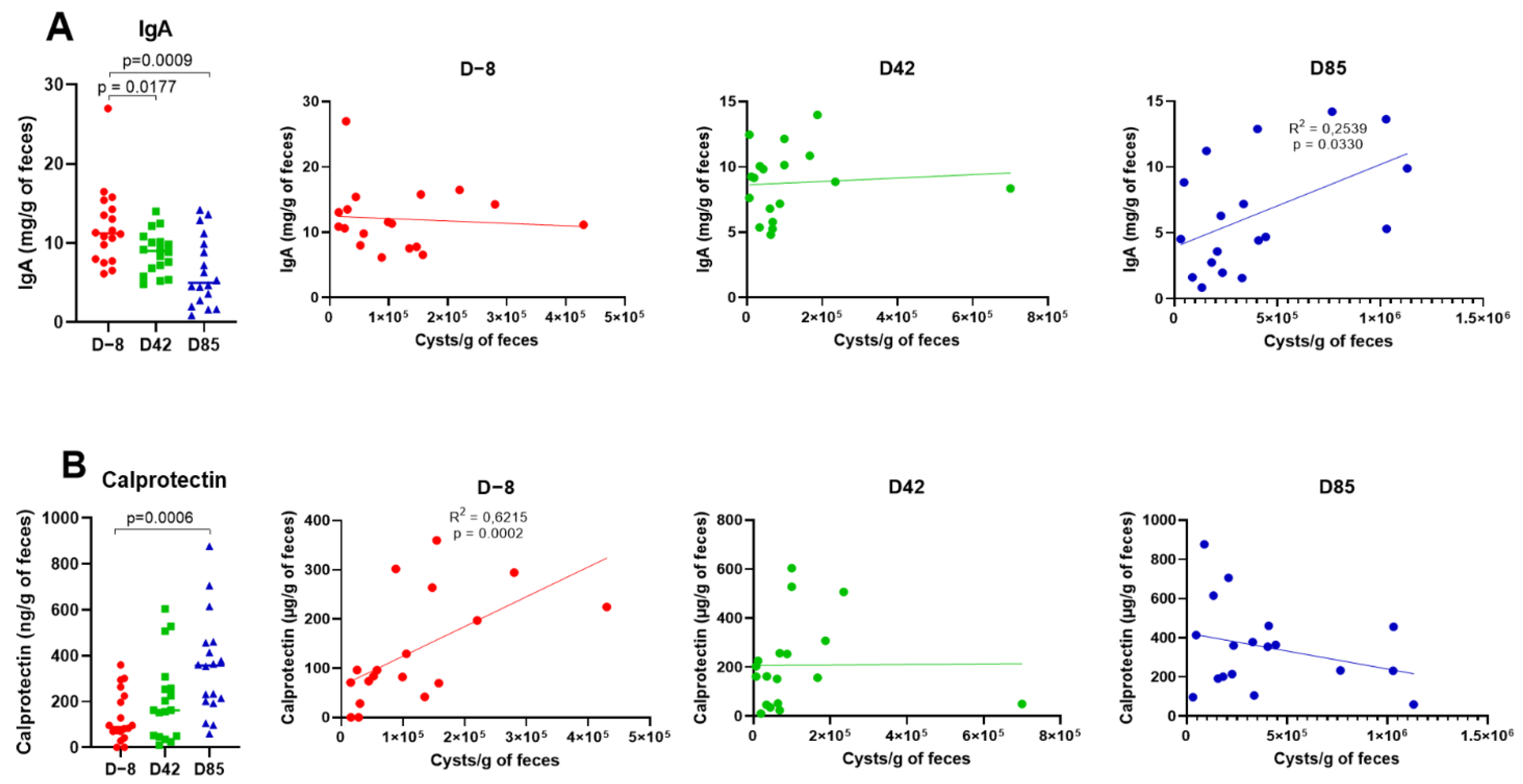
3.2. Age Influences Fecal IgA and Calprotectin Levels
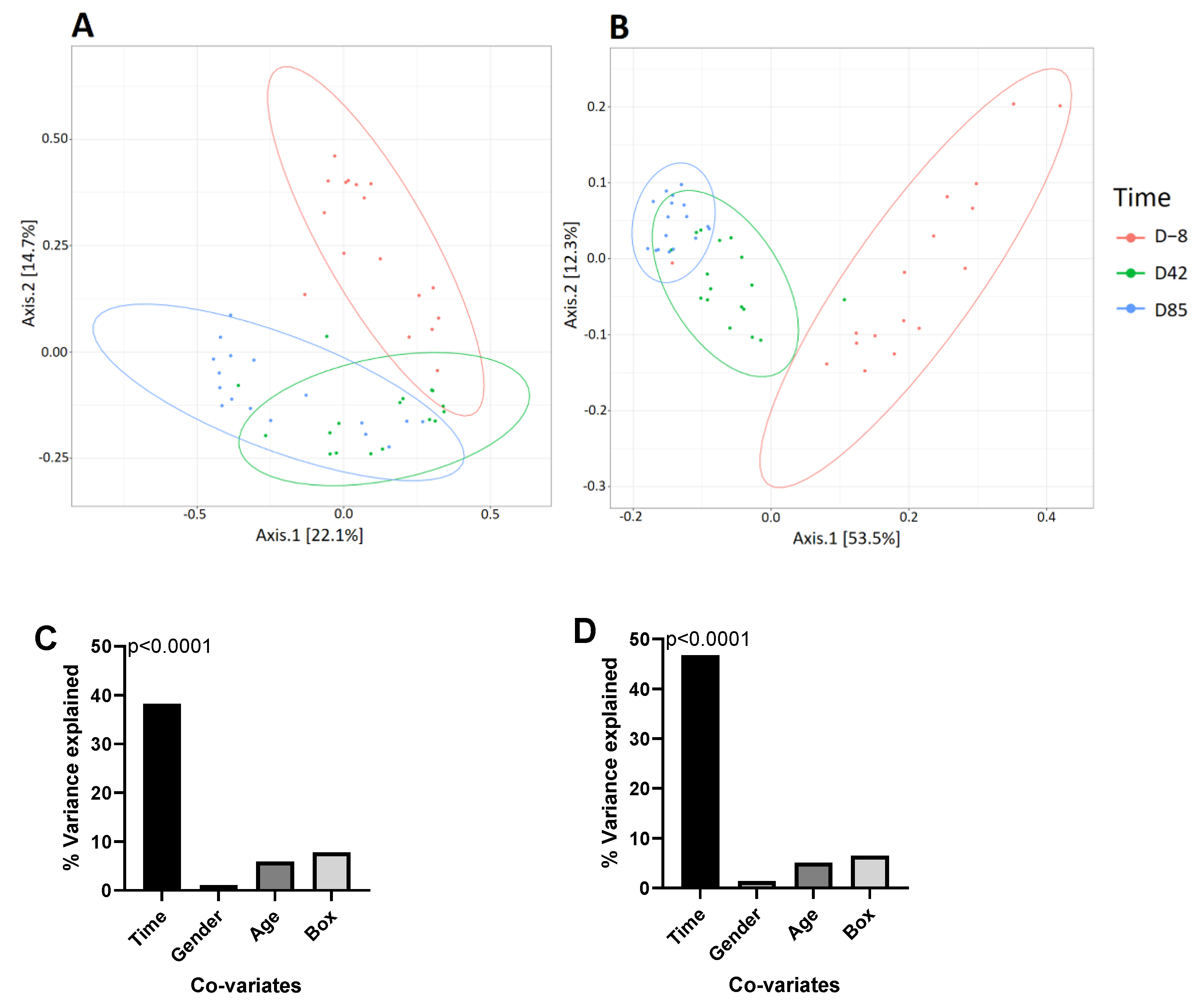
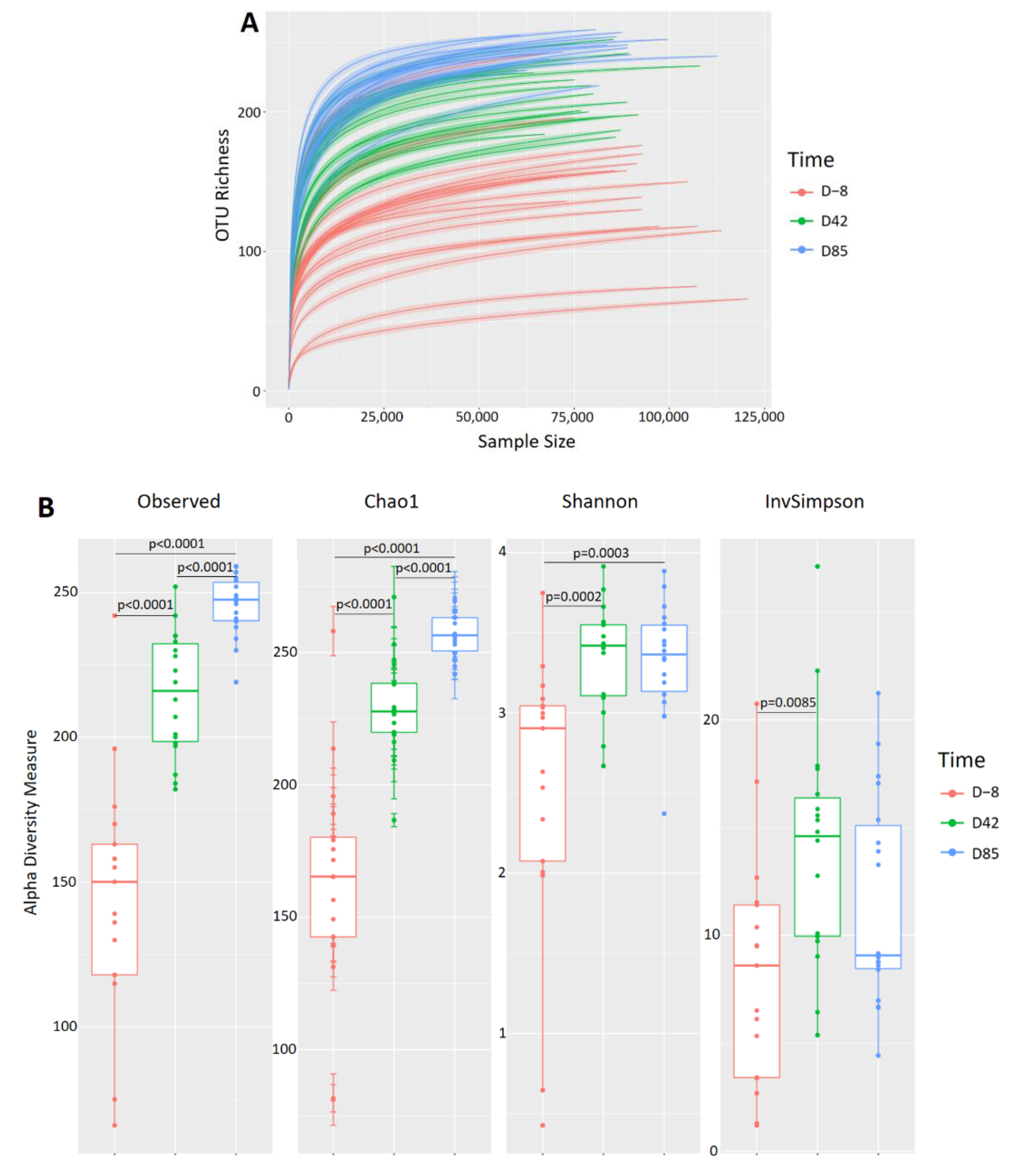
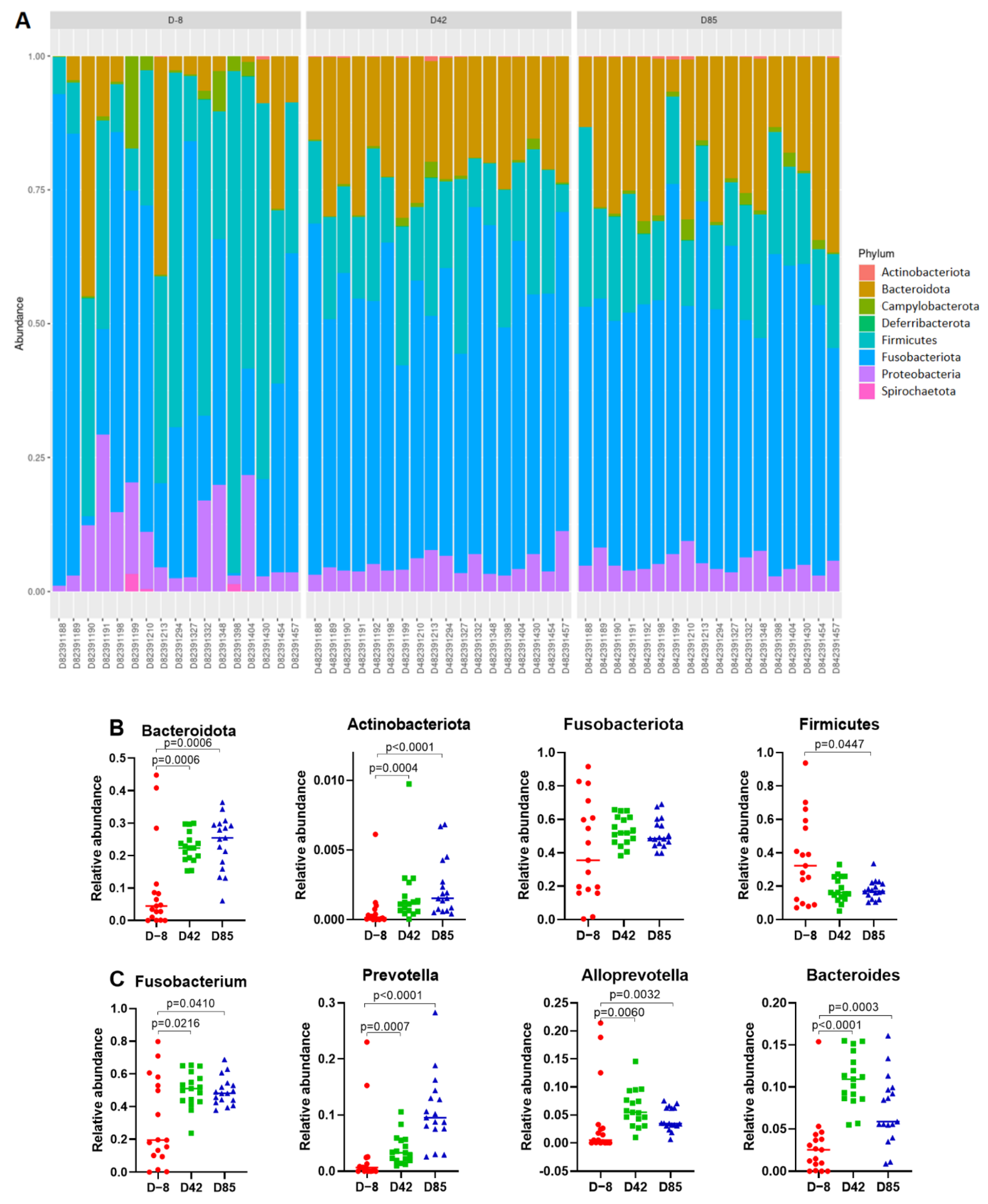
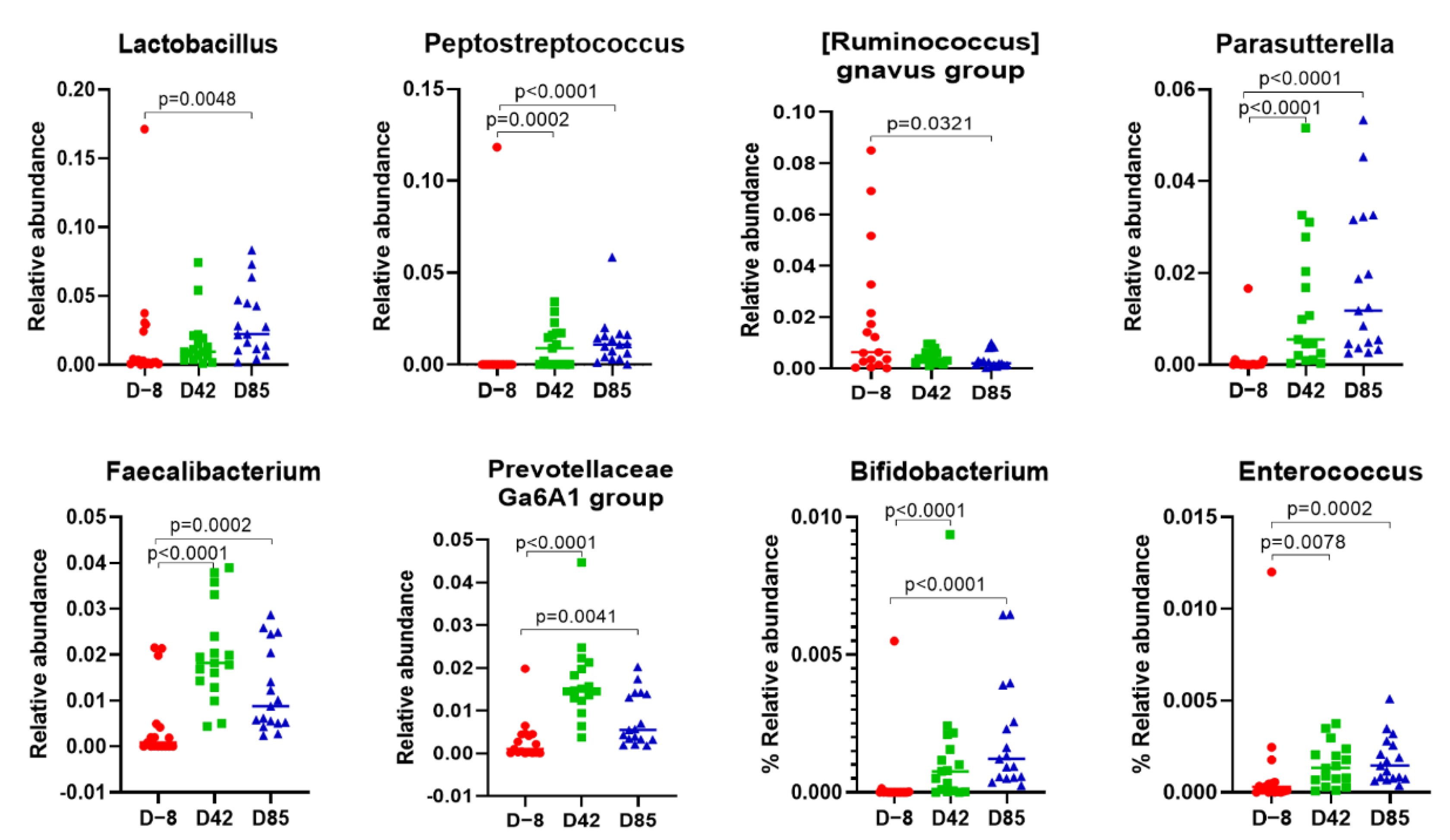
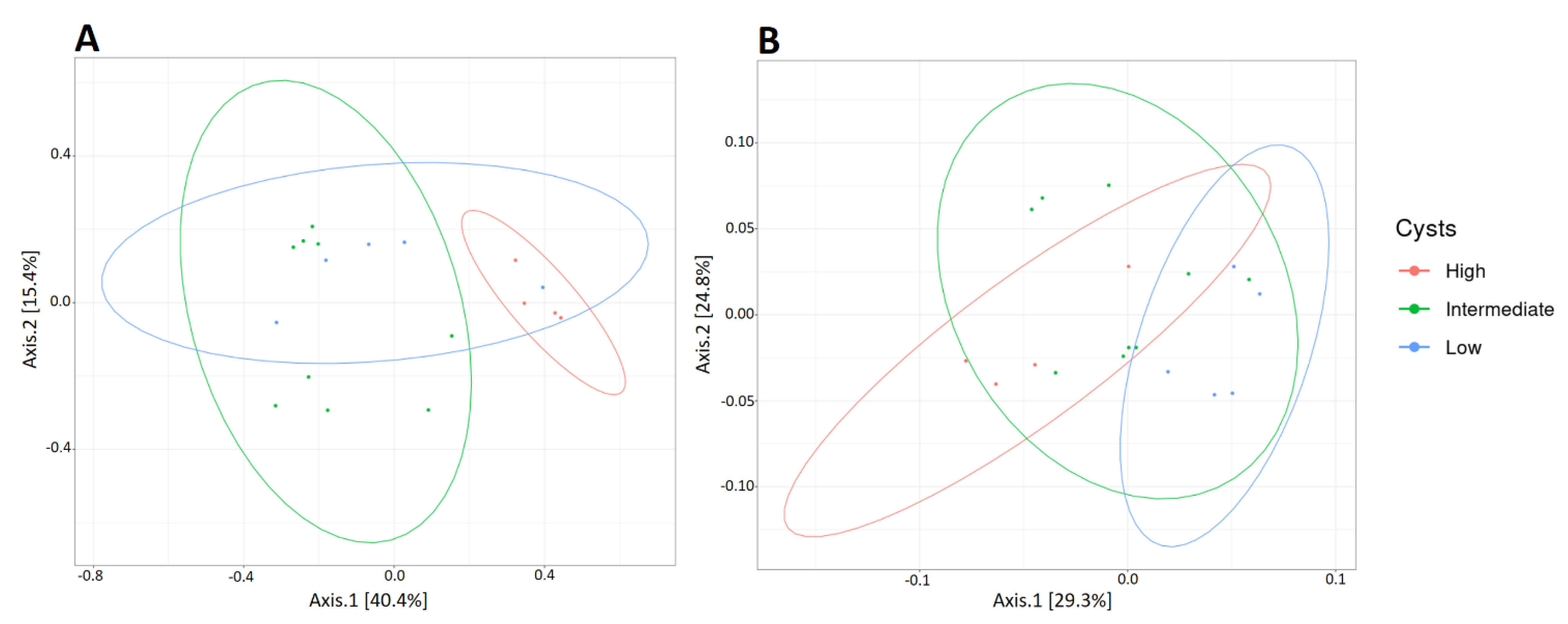
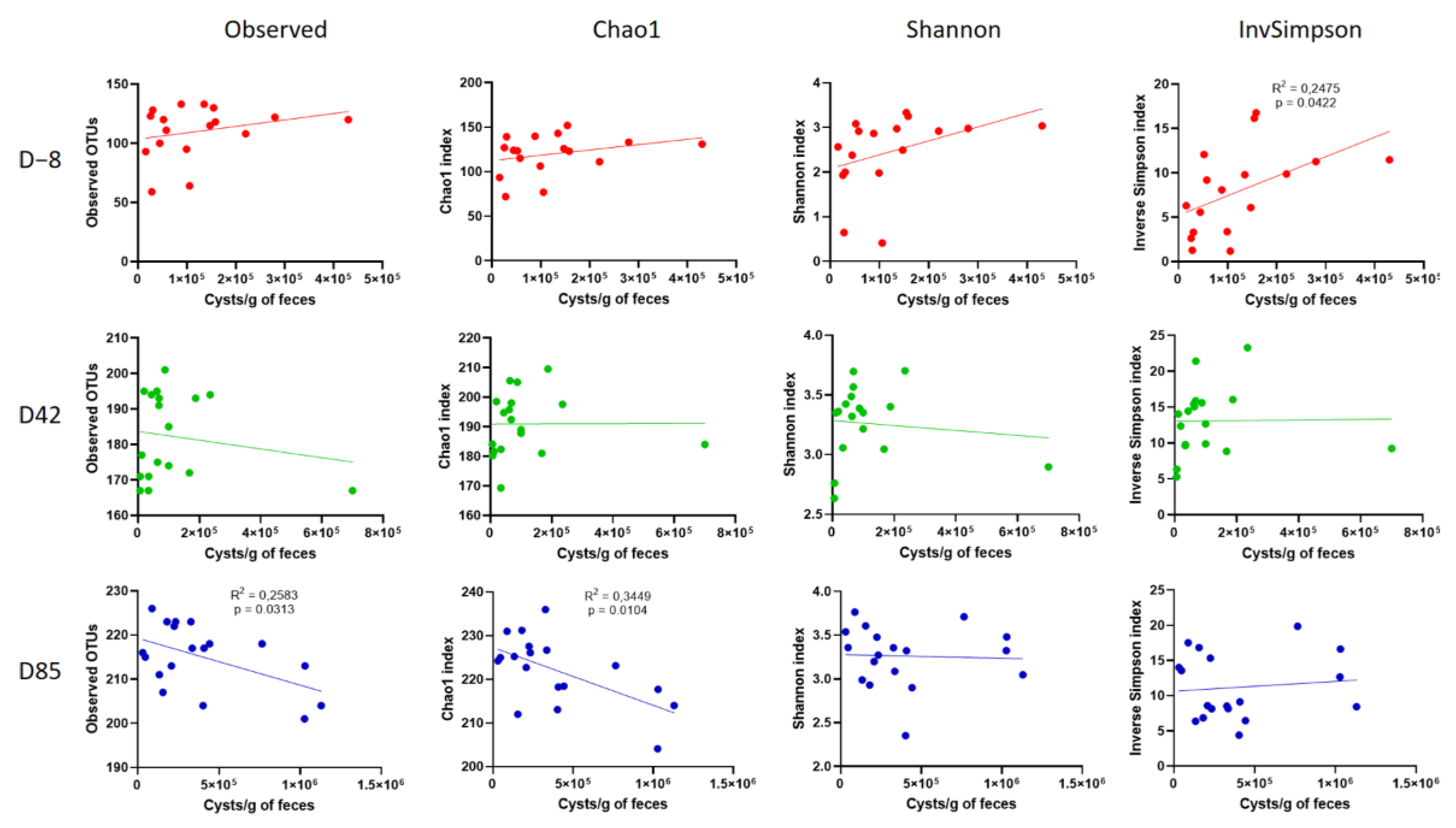
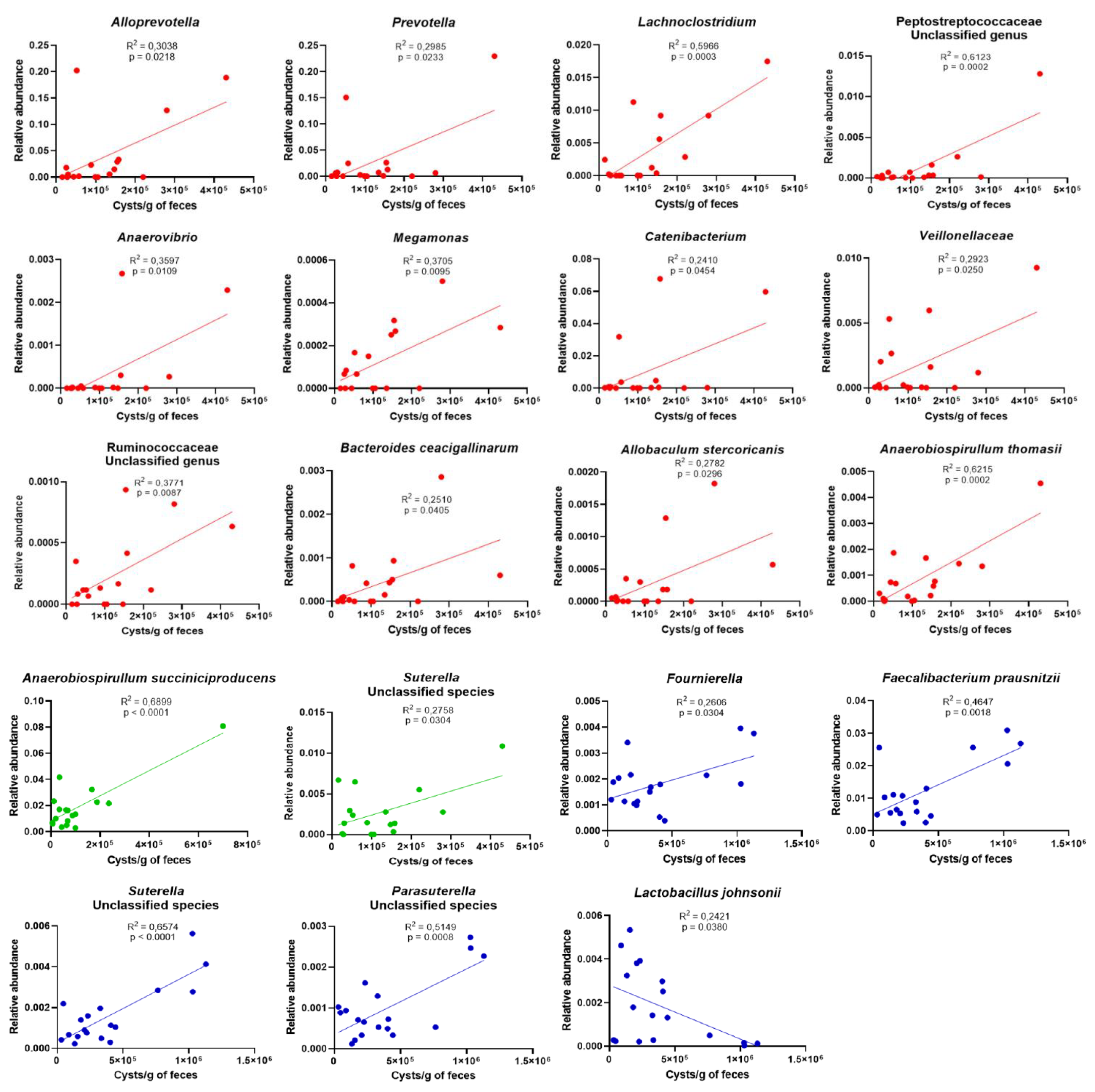
3.3. Age Influences the Richness and Diversity of the Canine Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cernikova, L.; Faso, C.; Hehl, A.B. Five Facts about Giardia Lamblia. PLoS Pathog. 2018, 14, e1007250. [Google Scholar] [CrossRef] [Green Version]
- Santin, M. Cryptosporidium and Giardia in Ruminants. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Buret, A.G.; Cacciò, S.M.; Favennec, L.; Svärd, S. Update on Giardia: Highlights from the Seventh International Giardia and Cryptosporidium Conference. Parasite 2020, 27, 49. [Google Scholar] [CrossRef]
- Adell-Aledón, M.; Köster, P.C.; de Lucio, A.; Puente, P.; Hernández-de-Mingo, M.; Sánchez-Thevenet, P.; Dea-Ayuela, M.A.; Carmena, D. Occurrence and Molecular Epidemiology of Giardia Duodenalis Infection in Dog Populations in Eastern Spain. BMC Vet. Res. 2018, 14, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, A.S.F.; Johnson, K.; Martins, R.; Sullivan, M.C.; Farias Amorim, C.; Putre, A.; Scott, A.; Wang, S.; Lindsay, B.; Baldassano, R.N.; et al. Natural Infection with Giardia Is Associated with Altered Community Structure of the Human and Canine Gut Microbiome. mSphere 2020, 5, e00670-20. [Google Scholar] [CrossRef]
- Tysnes, K.R.; Skancke, E.; Robertson, L.J. Subclinical Giardia in Dogs: A Veterinary Conundrum Relevant to Human Infection. Trends Parasitol. 2014, 30, 520–527. [Google Scholar] [CrossRef]
- Cacciò, S.M.; Lalle, M.; Svärd, S.G. Host Specificity in the Giardia Duodenalis Species Complex. Infect. Genet. Evol. 2018, 66, 335–345. [Google Scholar] [CrossRef]
- Hinney, B.; Gottwald, M.; Moser, J.; Reicher, B.; Schäfer, B.J.; Schaper, R.; Joachim, A.; Künzel, F. Examination of Anonymous Canine Faecal Samples Provides Data on Endoparasite Prevalence Rates in Dogs for Comparative Studies. Vet. Parasitol. 2017, 245, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Uiterwijk, M.; Nijsse, R.; Kooyman, F.N.J.; Wagenaar, J.A.; Mughini-Gras, L.; Ploeger, H.W. Host Factors Associated with Giardia Duodenalis Infection in Dogs across Multiple Diagnostic Tests. Parasit. Vectors 2019, 12, 556. [Google Scholar] [CrossRef] [Green Version]
- Bouzid, M.; Halai, K.; Jeffreys, D.; Hunter, P.R. The Prevalence of Giardia Infection in Dogs and Cats, a Systematic Review and Meta-Analysis of Prevalence Studies from Stool Samples. Vet. Parasitol. 2015, 207, 181–202. [Google Scholar] [CrossRef] [Green Version]
- Grellet, A.; Heilmann, R.M.; Polack, B.; Feugier, A.; Boucraut-Baralon, C.; Grandjean, D.; Grützner, N.; Suchodolski, J.S.; Steiner, J.M.; Chastant-Maillard, S. Influence of Breed Size, Age, Fecal Quality, and Enteropathogen Shedding on Fecal Calprotectin and Immunoglobulin A Concentrations in Puppies During the Weaning Period. J. Vet. Intern. Med. 2016, 30, 1056–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, M.F.; Rupp, P.; Pietsch, M.; Kaspar, A.; Beelitz, P. Giardia in a Selected Population of Dogs and Cats in Germany—Diagnostics, Coinfections and Assemblages. Vet. Parasitol. 2018, 249, 49–56. [Google Scholar] [CrossRef]
- Liao, S.; Lin, X.; Sun, Y.; Qi, N.; Lv, M.; Wu, C.; Li, J.; Hu, J.; Yu, L.; Cai, H.; et al. Occurrence and Genotypes of Cryptosporidium spp., Giardia duodenalis, and Blastocystis sp. in Household, Shelter, Breeding, and Pet Market Dogs in Guangzhou, Southern China. Sci. Rep. 2020, 10, 17736. [Google Scholar] [CrossRef] [PubMed]
- Minamoto, Y.; Otoni, C.C.; Steelman, S.M.; Büyükleblebici, O.; Steiner, J.M.; Jergens, A.E.; Suchodolski, J.S. Alteration of the Fecal Microbiota and Serum Metabolite Profiles in Dogs with Idiopathic Inflammatory Bowel Disease. Gut Microbes 2015, 6, 33–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandri, G.; Milani, C.; Mancabelli, L.; Mangifesta, M.; Lugli, G.A.; Viappiani, A.; Duranti, S.; Turroni, F.; Ossiprandi, M.C.; van Sinderen, D.; et al. Metagenomic Dissection of the Canine Gut Microbiota: Insights into Taxonomic, Metabolic and Nutritional Features. Environ. Microbiol. 2019, 21, 1331–1343. [Google Scholar] [CrossRef]
- Coelho, L.P.; Kultima, J.R.; Costea, P.I.; Fournier, C.; Pan, Y.; Czarnecki-Maulden, G.; Hayward, M.R.; Forslund, S.K.; Schmidt, T.S.B.; Descombes, P.; et al. Similarity of the Dog and Human Gut Microbiomes in Gene Content and Response to Diet. Microbiome 2018, 6, 72. [Google Scholar] [CrossRef]
- Guard, B.C.; Mila, H.; Steiner, J.M.; Mariani, C.; Suchodolski, J.S.; Chastant-Maillard, S. Characterization of the Fecal Microbiome during Neonatal and Early Pediatric Development in Puppies. PLoS ONE 2017, 12, e0175718. [Google Scholar] [CrossRef] [Green Version]
- Chun, J.L.; Ji, S.Y.; Lee, S.D.; Lee, Y.K.; Kim, B.; Kim, K.H. Difference of Gut Microbiota Composition Based on the Body Condition Scores in Dogs. J. Anim. Sci. Technol. 2020, 62, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Jha, A.R.; Shmalberg, J.; Tanprasertsuk, J.; Perry, L.; Massey, D.; Honaker, R.W. Characterization of Gut Microbiomes of Household Pets in the United States Using a Direct-to-Consumer Approach. PLoS ONE 2020, 15, e0227289. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.E.; Kim, H.-R.; Jeong, J.Y.; So, K.-M.; Lee, S.; Ji, S.Y.; Kim, M.; Lee, H.-J.; Lee, S.; Kim, K.-H.; et al. Impact of Breed on the Fecal Microbiome of Dogs under the Same Dietary Condition. J. Microbiol. Biotechnol. 2019, 29, 1947–1956. [Google Scholar] [CrossRef]
- Kubinyi, E.; Bel Rhali, S.; Sándor, S.; Szabó, A.; Felföldi, T. Gut Microbiome Composition Is Associated with Age and Memory Performance in Pet Dogs. Animals 2020, 10, 1488. [Google Scholar] [CrossRef]
- Mizukami, K.; Uchiyama, J.; Igarashi, H.; Murakami, H.; Osumi, T.; Shima, A.; Ishiahra, G.; Nasukawa, T.; Une, Y.; Sakaguchi, M. Age-Related Analysis of the Gut Microbiome in a Purebred Dog Colony. FEMS Microbiol. Lett. 2019, 366, fnz095. [Google Scholar] [CrossRef]
- Park, J.S.; Guevarra, R.B.; Kim, B.-R.; Lee, J.H.; Lee, S.H.; Cho, J.H.; Kim, H.; Cho, J.H.; Song, M.; Lee, J.-H.; et al. Intestinal Microbial Dysbiosis in Beagles Naturally Infected with Canine Parvovirus. J. Microbiol. Biotechnol. 2019, 29, 1391–1400. [Google Scholar] [CrossRef]
- Singer, S.M.; Nash, T.E. The Role of Normal Flora in Giardia Lamblia Infections in Mice. J. Infect. Dis. 2000, 181, 1510–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, M.F.; Uetanabaro, A.P.T.; Costa, A.F.; Alves, C.A.; Farias, L.M.; Bambirra, E.A.; Penna, F.J.; Vieira, E.C.; Nicoli, J.R. Influence of Bacteria from the Duodenal Microbiota of Patients with Symptomatic Giardiasis on the Pathogenicity of Giardia Duodenalis in Gnotoxenic Mice. J. Med. Microbiol. 2000, 49, 209–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerbaba, T.K.; Gupta, P.; Rioux, K.; Hansen, D.; Buret, A.G. Giardia Duodenalis-Induced Alterations of Commensal Bacteria Kill Caenorhabditis Elegans: A New Model to Study Microbial-Microbial Interactions in the Gut. Am. J. Physiol Gastrointest. Liver Physiol. 2015, 308, G550–G561. [Google Scholar] [CrossRef] [Green Version]
- Beatty, J.K.; Akierman, S.V.; Motta, J.-P.; Muise, S.; Workentine, M.L.; Harrison, J.J.; Bhargava, A.; Beck, P.L.; Rioux, K.P.; McKnight, G.W.; et al. Giardia Duodenalis Induces Pathogenic Dysbiosis of Human Intestinal Microbiota Biofilms. Int. J. Parasitol. 2017, 47, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Šlapeta, J.; Dowd, S.E.; Alanazi, A.D.; Westman, M.E.; Brown, G.K. Differences in the Faecal Microbiome of Non-Diarrhoeic Clinically Healthy Dogs and Cats Associated with Giardia Duodenalis Infection: Impact of Hookworms and Coccidia. Int. J. Parasitol. 2015, 45, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Read, C.M.; Monis, P.T.; Andrew Thompson, R.C. Discrimination of All Genotypes of Giardia Duodenalis at the Glutamate Dehydrogenase Locus Using PCR-RFLP. Infect. Genet. Evol. 2004, 4, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.M.; Fayer, R.; Bern, C.; Gilman, R.H.; Trout, J.M.; Schantz, P.M.; Das, P.; Lal, A.A.; Xiao, L. Triosephosphate Isomerase Gene Characterization and Potential Zoonotic Transmission of Giardia Duodenalis. Emerg. Infect. Dis. 2003, 9, 1444–1452. [Google Scholar] [CrossRef]
- Cacciò, S.M.; De Giacomo, M.; Pozio, E. Sequence Analysis of the β-Giardin Gene and Development of a Polymerase Chain Reaction–Restriction Fragment Length Polymorphism Assay to Genotype Giardia Duodenalis Cysts from Human Faecal Samples. Int. J. Parasitol. 2002, 32, 1023–1030. [Google Scholar] [CrossRef]
- Lalle, M.; Pozio, E.; Capelli, G.; Bruschi, F.; Crotti, D.; Cacciò, S.M. Genetic Heterogeneity at the β-Giardin Locus among Human and Animal Isolates of Giardia Duodenalis and Identification of Potentially Zoonotic Subgenotypes. Int. J. Parasitol. 2005, 35, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, R.M.; Meloni, B.P.; Groth, D.M.; Wetherall, J.D.; Reynoldson, J.A.; Thompson, R.C. Ribosomal RNA Sequencing Reveals Differences between the Genotypes of Giardia Isolates Recovered from Humans and Dogs Living in the Same Locality. J. Parasitol. 1997, 83, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Nadkarni, M.A.; Martin, F.E.; Jacques, N.A.; Hunter, N. Determination of Bacterial Load by Real-Time PCR Using a Broad-Range (Universal) Probe and Primers Set. Microbiology 2002, 148, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lluch, J.; Servant, F.; Païssé, S.; Valle, C.; Valière, S.; Kuchly, C.; Vilchez, G.; Donnadieu, C.; Courtney, M.; Burcelin, R.; et al. The Characterization of Novel Tissue Microbiota Using an Optimized 16S Metagenomic Sequencing Pipeline. PLoS ONE 2015, 10, e0142334. [Google Scholar] [CrossRef]
- Heilmann, R.M.; Suchodolski, J.S.; Steiner, J.M. Development and Analytic Validation of a Radioimmunoassay for the Quantification of Canine Calprotectin in Serum and Feces from Dogs. Am. J. Vet. Res. 2008, 69, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Godon, J.J.; Zumstein, E.; Dabert, P.; Habouzit, F.; Moletta, R. Molecular Microbial Diversity of an Anaerobic Digestor as Determined by Small-Subunit RDNA Sequence Analysis. Appl. Environ. Microbiol. 1997, 63, 2802–2813. [Google Scholar] [CrossRef] [Green Version]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef]
- Cacciò, S.M.; Beck, R.; Lalle, M.; Marinculic, A.; Pozio, E. Multilocus Genotyping of Giardia Duodenalis Reveals Striking Differences between Assemblages A and B. Int. J. Parasitol. 2008, 38, 1523–1531. [Google Scholar] [CrossRef]
- Donowitz, J.R.; Alam, M.; Kabir, M.; Ma, J.Z.; Nazib, F.; Platts-Mills, J.A.; Bartelt, L.A.; Haque, R.; Petri, W.A. A Prospective Longitudinal Cohort to Investigate the Effects of Early Life Giardiasis on Growth and All Cause Diarrhea. Clin. Infect. Dis. 2016, 63, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Hanevik, K.; Wensaas, K.-A.; Rortveit, G.; Eide, G.E.; Mørch, K.; Langeland, N. Irritable Bowel Syndrome and Chronic Fatigue 6 Years After Giardia Infection: A Controlled Prospective Cohort Study. Clin. Infect. Dis. 2014, 59, 1394–1400. [Google Scholar] [CrossRef] [PubMed]
- Riba, A.; Hassani, K.; Walker, A.; van Best, N.; von Zeschwitz, D.; Anslinger, T.; Sillner, N.; Rosenhain, S.; Eibach, D.; Maiga-Ascofaré, O.; et al. Disturbed Gut Microbiota and Bile Homeostasis in Giardia -Infected Mice Contributes to Metabolic Dysregulation and Growth Impairment. Sci. Transl. Med. 2020, 12, eaay7019. [Google Scholar] [CrossRef]
- Kim, M.; Vogtmann, E.; Ahlquist, D.A.; Devens, M.E.; Kisiel, J.B.; Taylor, W.R.; White, B.A.; Hale, V.L.; Sung, J.; Chia, N.; et al. Fecal Metabolomic Signatures in Colorectal Adenoma Patients Are Associated with Gut Microbiota and Early Events of Colorectal Cancer Pathogenesis. mBio 2020, 11, e03186-19. [Google Scholar] [CrossRef] [Green Version]
- Perrucci, S.; Berrilli, F.; Procopio, C.; Di Filippo, M.M.; Pierini, A.; Marchetti, V. Giardia Duodenalis infection in Dogs Affected by Primary Chronic Enteropathy. Open Vet. J. 2020, 10, 74–79. [Google Scholar] [CrossRef]
- Hjøllo, T.; Bratland, E.; Steinsland, H.; Radunovic, M.; Langeland, N.; Hanevik, K. Longitudinal Cohort Study of Serum Antibody Responses towards Giardia Lamblia Variant-Specific Surface Proteins in a Non-Endemic Area. Exp. Parasitol. 2018, 191, 66–72. [Google Scholar] [CrossRef]
- Langford, T.D.; Housley, M.P.; Boes, M.; Chen, J.; Kagnoff, M.F.; Gillin, F.D.; Eckmann, L. Central Importance of Immunoglobulin A in Host Defense against Giardia spp. Infect. Immun. 2002, 70, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Bridgman, S.L.; Konya, T.; Azad, M.B.; Sears, M.R.; Becker, A.B.; Turvey, S.E.; Mandhane, P.J.; Subbarao, P.; CHILD Study Investigators; Scott, J.A.; et al. Infant Gut Immunity: A Preliminary Study of IgA Associations with Breastfeeding. J. Dev. Orig. Health Dis. 2016, 7, 68–72. [Google Scholar] [CrossRef]
- Janzon, A.; Goodrich, J.K.; Koren, O.; Waters, J.L.; Ley, R.E. Interactions between the Gut Microbiome and Mucosal Immunoglobulins A, M, and G in the Developing Infant Gut. mSystems 2019, 4, e00612-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paerewijck, O.; Maertens, B.; Gagnaire, A.; De Bosscher, K.; Geldhof, P. Delayed Development of the Protective IL-17A Response Following a Giardia Muris Infection in Neonatal Mice. Sci. Rep. 2019, 9, 8959. [Google Scholar] [CrossRef] [PubMed]
- Ayling, R.M.; Kok, K. Fecal Calprotectin. In Advances in Clinical Chemistry; Elsevier: Amsterdam, The Netherlands, 2018; Volume 87, pp. 161–190. ISBN 978-0-12-815203-4. [Google Scholar]
- Ricciuto, A.; Griffiths, A.M. Clinical Value of Fecal Calprotectin. Crit. Rev. Clin. Lab. Sci. 2019, 56, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Grellet, A.; Mila, H.; Heilmann, R.M.; Feugier, A.; Gruetzner, N.; Suchodolski, J.S.; Steiner, J.M.; Chastant-Maillard, S. Effect of Age, Gestation and Lactation on Faecal IgA and Calprotectin Concentrations in Dogs. J. Nutr. Sci. 2014, 3, e41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-Méndez, M.-J.; Romero-Abal, M.-E.; Schümann, K.; Gil, Á.; Solomons, N.W. Normative Fecal Calprotectin Concentrations in Guatemalan Preschoolers Are High Relative to Children Reported Elsewhere. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 238–244. [Google Scholar] [CrossRef]
- Hestvik, E.; Tumwine, J.K.; Tylleskar, T.; Grahnquist, L.; Ndeezi, G.; Kaddu-Mulindwa, D.H.; Aksnes, L.; Olafsdottir, E. Faecal Calprotectin Concentrations in Apparently Healthy Children Aged 0–12 Years in Urban Kampala, Uganda: A Community-Based Survey. BMC Pediatr. 2011, 11, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanevik, K.; Hausken, T.; Morken, M.H.; Strand, E.A.; Mørch, K.; Coll, P.; Helgeland, L.; Langeland, N. Persisting Symptoms and Duodenal Inflammation Related to Giardia Duodenalis Infection. J. Infect. 2007, 55, 524–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.; Abe, F.; Osawa, R. Age-Related Changes in Gut Microbiota Composition from Newborn to Centenarian: A Cross-Sectional Study. BMC Microbiol. 2016, 16, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandri, G.; Argentini, C.; Milani, C.; Turroni, F.; Cristina Ossiprandi, M.; Sinderen, D.; Ventura, M. Catching a Glimpse of the Bacterial Gut Community of Companion Animals: A Canine and Feline Perspective. Microb. Biotechnol. 2020, 13, 1708–1732. [Google Scholar] [CrossRef] [PubMed]
- Omatsu, T.; Omura, M.; Katayama, Y.; Kimura, T.; Okumura, M.; Okumura, A.; Murata, Y.; Mizutani, T. Molecular Diversity of the Faecal Microbiota of Toy Poodles in Japan. J. Vet. Med. Sci. 2018, 80, 749–754. [Google Scholar] [CrossRef] [Green Version]
- Vilson, Å.; Ramadan, Z.; Li, Q.; Hedhammar, Å.; Reynolds, A.; Spears, J.; Labuda, J.; Pelker, R.; Björkstén, B.; Dicksved, J.; et al. Disentangling Factors That Shape the Gut Microbiota in German Shepherd Dogs. PLoS ONE 2018, 13, e0193507. [Google Scholar] [CrossRef]
- Masuoka, H.; Shimada, K.; Kiyosue-Yasuda, T.; Kiyosue, M.; Oishi, Y.; Kimura, S.; Yamada, A.; Hirayama, K. Transition of the Intestinal Microbiota of Dogs with Age. Biosci. Microbiota Food Health 2017, 36, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Fallani, M.; Amarri, S.; Uusijarvi, A.; Adam, R.; Khanna, S.; Aguilera, M.; Gil, A.; Vieites, J.M.; Norin, E.; Young, D.; et al. Determinants of the Human Infant Intestinal Microbiota after the Introduction of First Complementary Foods in Infant Samples from Five European Centres. Microbiology 2011, 157, 1385–1392. [Google Scholar] [CrossRef] [Green Version]
- Vallès, Y.; Artacho, A.; Pascual-García, A.; Ferrús, M.L.; Gosalbes, M.J.; Abellán, J.J.; Francino, M.P. Microbial Succession in the Gut: Directional Trends of Taxonomic and Functional Change in a Birth Cohort of Spanish Infants. PLoS Genet. 2014, 10, e1004406. [Google Scholar] [CrossRef] [Green Version]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and Functional Importance in the Gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of Short Chain Fatty Acids by the Gut Microbiota and Their Impact on Human Metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.C.; Dinsmoor, A.M.; Wang, M.; Donovan, S.M. Microbiome Composition in Pediatric Populations from Birth to Adolescence: Impact of Diet and Prebiotic and Probiotic Interventions. Dig. Dis. Sci. 2020, 65, 706–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.N.; Bidot, W.A.; Ericsson, A.C.; Franklin, C.L. Effects of Giardia Lamblia Colonization and Fenbendazole Treatment on Canine Fecal Microbiota. J. Am. Assoc. Lab. Anim. Sci. 2020. [Google Scholar] [CrossRef]
- Mejia, R.; Damania, A.; Jeun, R.; Bryan, P.E.; Vargas, P.; Juarez, M.; Cajal, P.S.; Nasser, J.; Krolewiecki, A.; Lefoulon, E.; et al. Impact of Intestinal Parasites on Microbiota and Cobalamin Gene Sequences: A Pilot Study. Parasit. Vectors 2020, 13, 200. [Google Scholar] [CrossRef] [Green Version]
- Yordanova, I.A.; Cortés, A.; Klotz, C.; Kühl, A.A.; Heimesaat, M.M.; Cantacessi, C.; Hartmann, S.; Rausch, S. RORγt+ Treg to Th17 Ratios Correlate with Susceptibility to Giardia Infection. Sci. Rep. 2019, 9, 20328. [Google Scholar] [CrossRef] [Green Version]
- Atherly, T.; Rossi, G.; White, R.; Seo, Y.-J.; Wang, C.; Ackermann, M.; Breuer, M.; Allenspach, K.; Mochel, J.P.; Jergens, A.E. Glucocorticoid and Dietary Effects on Mucosal Microbiota in Canine Inflammatory Bowel Disease. PLoS ONE 2019, 14, e0226780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilla, R.; Suchodolski, J.S. The Role of the Canine Gut Microbiome and Metabolome in Health and Gastrointestinal Disease. Front. Vet. Sci. 2020, 6, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E. Prevotella in the Gut: Choose Carefully. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 69–70. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.M. The Immune Response to Prevotella Bacteria in Chronic Inflammatory Disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Iljazovic, A.; Roy, U.; Gálvez, E.J.C.; Lesker, T.R.; Zhao, B.; Gronow, A.; Amend, L.; Will, S.E.; Hofmann, J.D.; Pils, M.C.; et al. Perturbation of the Gut Microbiome by Prevotella Spp. Enhances Host Susceptibility to Mucosal Inflammation. Mucosal Immunol. 2021, 14, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Rouhani, S.; Griffin, N.W.; Yori, P.P.; Olortegui, M.P.; Siguas Salas, M.; Rengifo Trigoso, D.; Moulton, L.H.; Houpt, E.R.; Barratt, M.J.; Kosek, M.N.; et al. Gut Microbiota Features Associated With Campylobacter Burden and Postnatal Linear Growth Deficits in a Peruvian Birth Cohort. Clin. Infect. Dis. 2020, 71, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Toro-Londono, M.A.; Bedoya-Urrego, K.; Garcia-Montoya, G.M.; Galvan-Diaz, A.L.; Alzate, J.F. Intestinal Parasitic Infection Alters Bacterial Gut Microbiota in Children. PeerJ 2019, 7, e6200. [Google Scholar] [CrossRef] [Green Version]
- Hughes, E.R.; Winter, M.G.; Duerkop, B.A.; Spiga, L.; Furtado de Carvalho, T.; Zhu, W.; Gillis, C.C.; Büttner, L.; Smoot, M.P.; Behrendt, C.L.; et al. Microbial Respiration and Formate Oxidation as Metabolic Signatures of Inflammation-Associated Dysbiosis. Cell Host Microbe 2017, 21, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Litvak, Y.; Byndloss, M.X.; Tsolis, R.M.; Bäumler, A.J. Dysbiotic Proteobacteria Expansion: A Microbial Signature of Epithelial Dysfunction. Curr. Opin. Microbiol. 2017, 39, 1–6. [Google Scholar] [CrossRef]
- Lopez, C.A.; Miller, B.M.; Rivera-Chavez, F.; Velazquez, E.M.; Byndloss, M.X.; Chavez-Arroyo, A.; Lokken, K.L.; Tsolis, R.M.; Winter, S.E.; Baumler, A.J. Virulence Factors Enhance Citrobacter Rodentium Expansion through Aerobic Respiration. Science 2016, 353, 1249–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-Chávez, F.; Zhang, L.F.; Faber, F.; Lopez, C.A.; Byndloss, M.X.; Olsan, E.E.; Xu, G.; Velazquez, E.M.; Lebrilla, C.B.; Winter, S.E.; et al. Depletion of Butyrate-Producing Clostridia from the Gut Microbiota Drives an Aerobic Luminal Expansion of Salmonella. Cell Host Microbe 2016, 19, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barash, N.R.; Maloney, J.G.; Singer, S.M.; Dawson, S.C. Giardia Alters Commensal Microbial Diversity throughout the Murine Gut. Infect. Immun. 2017, 85, e00948-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allain, T.; Fekete, E.; Buret, A.G. Giardia Cysteine Proteases: The Teeth behind the Smile. Trends Parasitol. 2019, 35, 636–648. [Google Scholar] [CrossRef]
- Bhargava, A.; Cotton, J.A.; Dixon, B.R.; Gedamu, L.; Yates, R.M.; Buret, A.G. Giardia Duodenalis Surface Cysteine Proteases Induce Cleavage of the Intestinal Epithelial Cytoskeletal Protein Villin via Myosin Light Chain Kinase. PLoS ONE 2015, 10, e0136102. [Google Scholar] [CrossRef] [Green Version]
- Courtman, N.F. Septic Peritonitis in a Dog Caused by Anaerobiospirillum Succiniproducens. Vet. Clin. Pathol. 2016, 45, 352–355. [Google Scholar] [CrossRef]
- Epstein, D.J.; Ernst, K.; Rogers, R.; Carmody, E.; Aguero-Rosenfeld, M. Closing the Brief Case: Anaerobiospirillum Succiniciproducens Bacteremia and Pyomyositis. J. Clin. Microbiol. 2017, 55, 986–987. [Google Scholar] [CrossRef] [Green Version]
- Mehmood, M.; Jaffar, N.A.; Nazim, M.; Khasawneh, F.A. Bacteremic Skin and Soft Tissue Infection Caused by Prevotella Loescheii. BMC Infect. Dis. 2014, 14, 162. [Google Scholar] [CrossRef] [Green Version]
- Mondo, E.; Barone, M.; Soverini, M.; D’Amico, F.; Cocchi, M.; Petrulli, C.; Mattioli, M.; Marliani, G.; Candela, M.; Accorsi, P.A. Gut Microbiome Structure and Adrenocortical Activity in Dogs with Aggressive and Phobic Behavioral Disorders. Heliyon 2020, 6, e03311. [Google Scholar] [CrossRef] [PubMed]
- Yusof, N.; Hamid, N.; Ma, Z.F.; Lawenko, R.M.; Wan Mohammad, W.M.Z.; Collins, D.A.; Liong, M.T.; Odamaki, T.; Xiao, J.; Lee, Y.Y. Exposure to Environmental Microbiota Explains Persistent Abdominal Pain and Irritable Bowel Syndrome after a Major Flood. Gut Pathog. 2017, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Roskens, H.; Erlandsen, S.L. Inhibition of In Vitro Attachment of Giardia Trophozoites by Mucin. J. Parasitol. 2002, 88, 869–873. [Google Scholar] [CrossRef]
- Yan, S.; Yang, B.; Zhao, J.; Zhao, J.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, W. A Ropy Exopolysaccharide Producing Strain Bifidobacterium Longum Subsp. Longum YS108R Alleviates DSS-Induced Colitis by Maintenance of the Mucosal Barrier and Gut Microbiota Modulation. Food Funct. 2019, 10, 1595–1608. [Google Scholar] [CrossRef] [PubMed]
- Praharaj, A.B.; Dehury, B.; Mahapatra, N.; Kar, S.K.; Behera, S.K. Molecular Dynamics Insights into the Structure, Function, and Substrate Binding Mechanism of Mucin Desulfating Sulfatase of Gut Microbe Bacteroides Fragilis. J. Cell. Biochem. 2018, 119, 3618–3631. [Google Scholar] [CrossRef]
- Rho, J.; Wright, D.P.; Christie, D.L.; Clinch, K.; Furneaux, R.H.; Roberton, A.M. A Novel Mechanism for Desulfation of Mucin: Identification and Cloning of a Mucin-Desulfating Glycosidase (Sulfoglycosidase) from Prevotella Strain RS2. J. Bacteriol. 2005, 187, 1543–1551. [Google Scholar] [CrossRef] [Green Version]
- Amat, C.B.; Motta, J.-P.; Fekete, E.; Moreau, F.; Chadee, K.; Buret, A.G. Cysteine Protease–Dependent Mucous Disruptions and Differential Mucin Gene Expression in Giardia duodenalis Infection. Am. J. Pathol. 2017, 187, 2486–2498. [Google Scholar] [CrossRef] [Green Version]
- Halliez, M.C.M.; Motta, J.-P.; Feener, T.D.; Guérin, G.; LeGoff, L.; François, A.; Colasse, E.; Favennec, L.; Gargala, G.; Lapointe, T.K.; et al. Giardia Duodenalis Induces Paracellular Bacterial Translocation and Causes Postinfectious Visceral Hypersensitivity. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G574–G585. [Google Scholar] [CrossRef] [Green Version]
- Allain, T.; Amat, C.B.; Motta, J.-P.; Manko, A.; Buret, A.G. Interactions of Giardia Sp. with the Intestinal Barrier: Epithelium, Mucus, and Microbiota. Tissue Barriers 2017, 5, e1274354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manko-Prykhoda, A.; Allain, T.; Motta, J.-P.; Cotton, J.A.; Feener, T.; Oyeyemi, A.; Bindra, S.; Vallance, B.A.; Wallace, J.L.; Beck, P.; et al. Giardia spp. Promote the Production of Antimicrobial Peptides and Attenuate Disease Severity Induced by Attaching and Effacing Enteropathogens via the Induction of the NLRP3 Inflammasome. Int. J. Parasitol. 2020, 50, 263–275. [Google Scholar] [CrossRef]
- Martínez, C.; Lasitschka, F.; Thöni, C.; Wohlfarth, C.; Braun, A.; Granzow, M.; Röth, R.; Dizdar, V.; Rappold, G.A.; Hausken, T.; et al. Comparative Expression Profiling in the Intestine of Patients with Giardia -induced Postinfectious Functional Gastrointestinal Disorders. Neurogastroenterol. Motil. 2020, 32. [Google Scholar] [CrossRef]
- Cerquetella, M.; Rossi, G.; Spaterna, A.; Tesei, B.; Jergens, A.; Suchodolski, J.; Bassotti, G. Is irritable bowel syndrome also present in dogs? Tierärztl. Prax. Ausg. K Kleintiere Heimtiere 2018, 46, 176–180. [Google Scholar] [CrossRef]
- Pei, L.; Ke, Y.; Zhao, H.; Wang, L.; Jia, C.; Liu, W.; Fu, Q.; Shi, M.; Cui, J.; Li, S. Role of Colonic Microbiota in the Pathogenesis of Ulcerative Colitis. BMC Gastroenterol. 2019, 19, 10. [Google Scholar] [CrossRef] [Green Version]
- Jalanka, J.; Cheng, J.; Hiippala, K.; Ritari, J.; Salojärvi, J.; Ruuska, T.; Kalliomäki, M.; Satokari, R. Colonic Mucosal Microbiota and Association of Bacterial Taxa with the Expression of Host Antimicrobial Peptides in Pediatric Ulcerative Colitis. Int. J. Mol. Sci. 2020, 21, 6044. [Google Scholar] [CrossRef]
- Lavelle, A.; Lennon, G.; O’Sullivan, O.; Docherty, N.; Balfe, A.; Maguire, A.; Mulcahy, H.E.; Doherty, G.; O’Donoghue, D.; Hyland, J.; et al. Spatial Variation of the Colonic Microbiota in Patients with Ulcerative Colitis and Control Volunteers. Gut 2015, 64, 1553–1561. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.-S.; Chang, P.-F.; Liao, C.-H.; Lee, T.-H.; Chen, Y.; Lee, Y.-C.; Wu, M.-S.; Wang, H.-P.; Ni, Y.-H. Differences of Microbiota in Small Bowel and Faeces between Irritable Bowel Syndrome Patients and Healthy Subjects. Scand. J. Gastroenterol. 2016, 51, 410–419. [Google Scholar] [CrossRef]
- Masoodi, I.; Alshanqeeti, A.S.; Alyamani, E.J.; AlLehibi, A.A.; Alqutub, A.N.; Alsayari, K.N.; Alomair, A.O. Microbial Dysbiosis in Irritable Bowel Syndrome: A Single-center Metagenomic Study in Saudi Arabia. JGH Open 2020, 4, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Tana, C.; Umesaki, Y.; Imaoka, A.; Handa, T.; Kanazawa, M.; Fukudo, S. Altered Profiles of Intestinal Microbiota and Organic Acids May Be the Origin of Symptoms in Irritable Bowel Syndrome. Neurogastroenterol. Motil. 2009. [Google Scholar] [CrossRef]
- Lo Presti, A.; Zorzi, F.; Del Chierico, F.; Altomare, A.; Cocca, S.; Avola, A.; De Biasio, F.; Russo, A.; Cella, E.; Reddel, S.; et al. Fecal and Mucosal Microbiota Profiling in Irritable Bowel Syndrome and Inflammatory Bowel Disease. Front. Microbiol. 2019, 10, 1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enqi, W.; Jingzhu, S.; Lingpeng, P.; Yaqin, L. Comparison of the Gut Microbiota Disturbance in Rat Models of Irritable Bowel Syndrome Induced by Maternal Separation and Multiple Early-Life Adversity. Front. Cell. Infect. Microbiol. 2021, 10, 581974. [Google Scholar] [CrossRef]
- Haange, S.-B.; Jehmlich, N.; Hoffmann, M.; Weber, K.; Lehmann, J.; von Bergen, M.; Slanina, U. Disease Development Is Accompanied by Changes in Bacterial Protein Abundance and Functions in a Refined Model of Dextran Sulfate Sodium (DSS)-Induced Colitis. J. Proteome Res. 2019, 18, 1774–1786. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-S.-E.; Li, W.-B.; Wang, H.-Y.; Ma, Y.-M.; Zhao, X.-H.; Yang, H.; Qian, J.-M.; Li, J.-N. VSL#3 Can Prevent Ulcerative Colitis-Associated Carcinogenesis in Mice. World J. Gastroenterol. 2018, 24, 4254–4262. [Google Scholar] [CrossRef]
- Suchodolski, J.S.; Markel, M.E.; Garcia-Mazcorro, J.F.; Unterer, S.; Heilmann, R.M.; Dowd, S.E.; Kachroo, P.; Ivanov, I.; Minamoto, Y.; Dillman, E.M.; et al. The Fecal Microbiome in Dogs with Acute Diarrhea and Idiopathic Inflammatory Bowel Disease. PLoS ONE 2012, 7, e51907. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-J.; Wu, H.; Wu, S.-D.; Lu, N.; Wang, Y.-T.; Liu, H.-N.; Dong, L.; Liu, T.-T.; Shen, X.-Z. Parasutterella, in Association with Irritable Bowel Syndrome and Intestinal Chronic Inflammation: Parasutterella May Be Related with IBS. J. Gastroenterol. Hepatol. 2018, 33, 1844–1852. [Google Scholar] [CrossRef]
- Chiodini, R.J.; Dowd, S.E.; Chamberlin, W.M.; Galandiuk, S.; Davis, B.; Glassing, A. Microbial Population Differentials between Mucosal and Submucosal Intestinal Tissues in Advanced Crohn’s Disease of the Ileum. PLoS ONE 2015, 10, e0134382. [Google Scholar] [CrossRef]
- Bloom, S.M.; Bijanki, V.N.; Nava, G.M.; Sun, L.; Malvin, N.P.; Donermeyer, D.L.; Dunne, W.M.; Allen, P.M.; Stappenbeck, T.S. Commensal Bacteroides Species Induce Colitis in Host-Genotype-Specific Fashion in a Mouse Model of Inflammatory Bowel Disease. Cell Host Microbe 2011, 9, 390–403. [Google Scholar] [CrossRef] [Green Version]
- Chao, G.; Zhang, S. The Characteristics of Intestinal Flora of IBS-D with Different Syndromes. Immun. Inflamm. Dis. 2020, 8, 615–628. [Google Scholar] [CrossRef]
- Pittayanon, R.; Lau, J.T.; Yuan, Y.; Leontiadis, G.I.; Tse, F.; Surette, M.; Moayyedi, P. Gut Microbiota in Patients With Irritable Bowel Syndrome—A Systematic Review. Gastroenterology 2019, 157, 97–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allain, T.; Chaouch, S.; Thomas, M.; Vallée, I.; Buret, A.G.; Langella, P.; Grellier, P.; Polack, B.; Bermúdez-Humarán, L.G.; Florent, I. Bile-Salt-Hydrolases from the Probiotic Strain Lactobacillus Johnsonii La1 Mediate Anti-Giardial Activity in Vitro and in Vivo. Front. Microbiol. 2018, 8, 2707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alazzaz, J.; Chaouch, S.; Boucard, A.-S.; Bermudez-Humaran, L.G.; Florent, I. Probiotics as Anti-Giardia Defenders: Overview on Putative Control Mechanisms. In Eukaryome Impact on Human Intestine Homeostasis and Mucosal Immunology; Guillen, N., Ed.; Springer International Publishing: Cham, Germany, 2020; pp. 335–349. ISBN 978-3-030-44825-7. [Google Scholar]
- Kodio, A.; Coulibaly, D.; Koné, A.K.; Konaté, S.; Doumbo, S.; Guindo, A.; Bittar, F.; Gouriet, F.; Raoult, D.; Thera, M.A.; et al. Blastocystis Colonization Is Associated with Increased Diversity and Altered Gut Bacterial Communities in Healthy Malian Children. Microorganisms 2019, 7, 649. [Google Scholar] [CrossRef] [Green Version]
- The Blastocystis Investigation Group; Audebert, C.; Even, G.; Cian, A.; Loywick, A.; Merlin, S.; Viscogliosi, E.; Chabé, M. Colonization with the Enteric Protozoa Blastocystis Is Associated with Increased Diversity of Human Gut Bacterial Microbiota. Sci. Rep. 2016, 6, 25255. [Google Scholar] [CrossRef]
- Allain, T.; Buret, A.G. Pathogenesis and post-infectious complications in giardiasis. In Advances in Parasitology; Elsevier: Amsterdam, The Netherlands, 2020; Volume 107, pp. 173–199. ISBN 978-0-12-820475-7. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Name | Sequence | Reference |
|---|---|---|---|
| gdh | GDHeF | TCAACGTYAAYCGYGGYTTCCGT | [29] |
| GDHiF | CAGTACAACTCYGCTCTCGG | ||
| GDHiR | GTTRTCCTTGCACATCTCC | ||
| tpi | AL3543 | AAATIATGCCTGCTCGTCG | [30] |
| AL3544 | CCCTTCATCGGIGGTAACTT | ||
| AL3545 | GTGGCCACCACICCCGTGCC | ||
| AL3546 | CAAACCTTITCCGCAAACC | ||
| β-giardin | G7 | AAGCCCGACGACCTCACCCGCAGTGC | [31] |
| G759 | GAGGCCGCCCTGGATCTTCGAGACGAC | ||
| G99 | GAACGAACGAGATCGAGGTCCG | [32] | |
| G609 | CTCGACGAGCTTCGTGTT | ||
| SSU rRNA | RH11 | CATCCGGTCGATCCTGCC | [33] |
| RH4 | AGTCGAAC CCTGATTCTCCGCCCAGG | ||
| GIAR-F | GACGCTCTCCCCAAGGAC | [29] | |
| GIAR-R | CTGCGTCACGCTCG | ||
| V3-V4 16S rDNA | PCR1F_343 | CTTTCCCTACACGACGCTCTTCCGATCT- ACGGRAGGCAGCAG partial P5 adapter–primer | [34] |
| PCR1_R784 | GGAGTTCAGACGTGTGCTCT- TCCGATCTTACCAGGGTATCTAATCCT partial P7 adapter–primer | ||
| PCR2_P5F | AATGATACGGCGACCACCGAGATCTACACT-CTTTCCCTACACGAC partial P5 adapter–primer targeting primer 1F | [35] | |
| PCR2_P7R | CAAGCAGAAGACGGCATACGAGAT-NNNNNN-GTGACT-GGAGTTCAGACGTGT partial P7 adapter including index–primer targeting primer 1R |
| Gene Loci | ||||
|---|---|---|---|---|
| gdh | tpi | β-giardin | SSU rRNA | |
| D−8 | Assemblage C = 6 Assemblage D = 11 Assemblage C or D = 0 Not amplified = 1 | Assemblage C = 13 Assemblage D = 2 Assemblage C or D = 0 Not amplified = 3 | Assemblage C = 2 Assemblage D = 14 Assemblage C or D = 0 Not amplified = 2 | Assemblage C = 0 Assemblage D = 2 Assemblage C or D = 14 Not amplified = 2 |
| D85 | Assemblage C = 7 Assemblage D = 2 Assemblage C or D = 0 Not amplified = 9 | Assemblage C = 7 Assemblage D = 0 Assemblage C or D = 0 Not amplified = 11 | Assemblage C = 4 Assemblage D = 1 Assemblage C or D = 0 Not amplified = 13 | Assemblage C = 3 Assemblage D = 5 Assemblage C or D = 6 Not amplified = 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boucard, A.-S.; Thomas, M.; Lebon, W.; Polack, B.; Florent, I.; Langella, P.; Bermúdez-Humarán, L.G. Age and Giardia intestinalis Infection Impact Canine Gut Microbiota. Microorganisms 2021, 9, 1862. https://doi.org/10.3390/microorganisms9091862
Boucard A-S, Thomas M, Lebon W, Polack B, Florent I, Langella P, Bermúdez-Humarán LG. Age and Giardia intestinalis Infection Impact Canine Gut Microbiota. Microorganisms. 2021; 9(9):1862. https://doi.org/10.3390/microorganisms9091862
Chicago/Turabian StyleBoucard, Anne-Sophie, Myriam Thomas, Wilfried Lebon, Bruno Polack, Isabelle Florent, Philippe Langella, and Luis G. Bermúdez-Humarán. 2021. "Age and Giardia intestinalis Infection Impact Canine Gut Microbiota" Microorganisms 9, no. 9: 1862. https://doi.org/10.3390/microorganisms9091862