
Adherent-Invasive and Non-Invasive Escherichia coli Isolates Differ in Their Effects on Caenorhabditis elegans’ Lifespan

 , , and
, , and 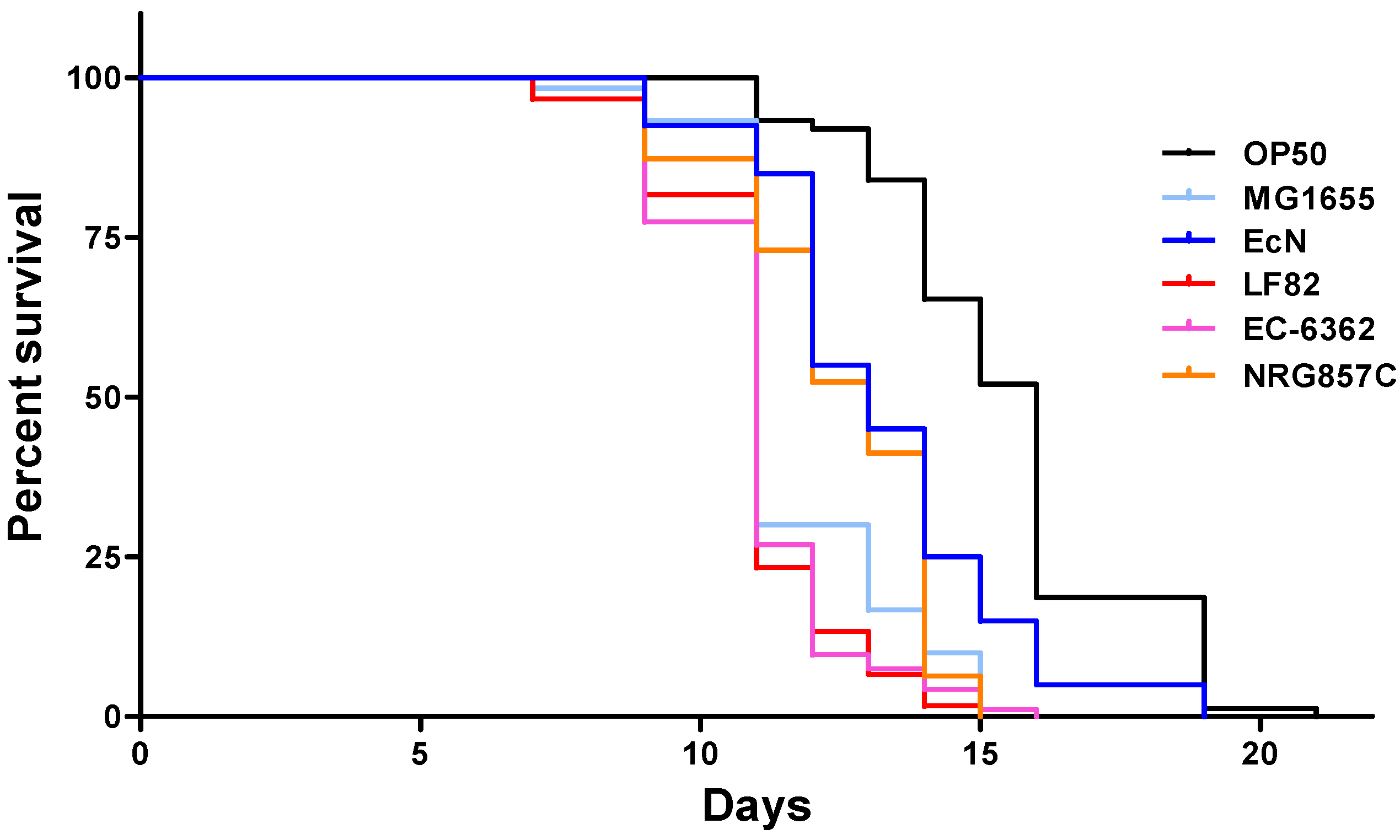{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Bacterial Strains and Culture Conditions
2.3. C. elegans Strains, Culture Conditions, and Longevity Assays
2.4. Cell Lines, Cell Culture, and Epithelial Cell Invasion Assays
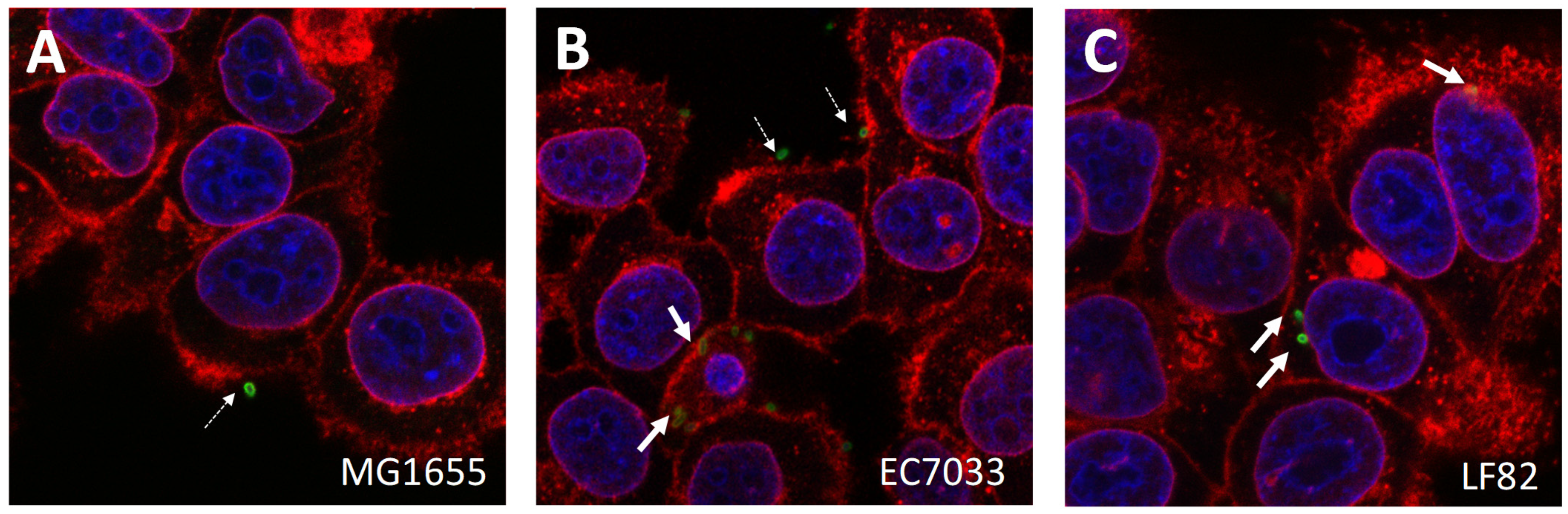
2.5. Microscopy
2.6. Statistical Analysis
3. Results
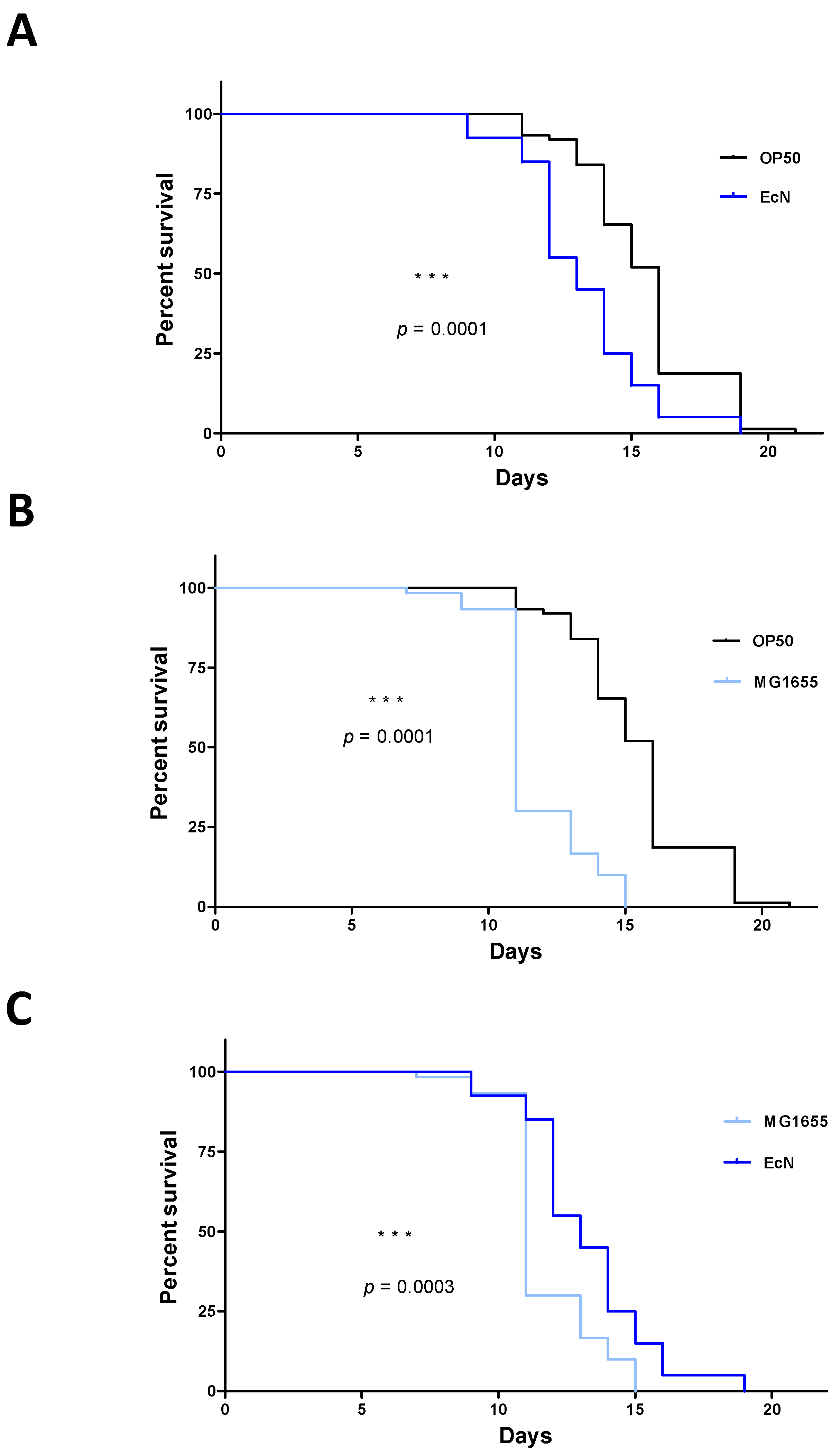
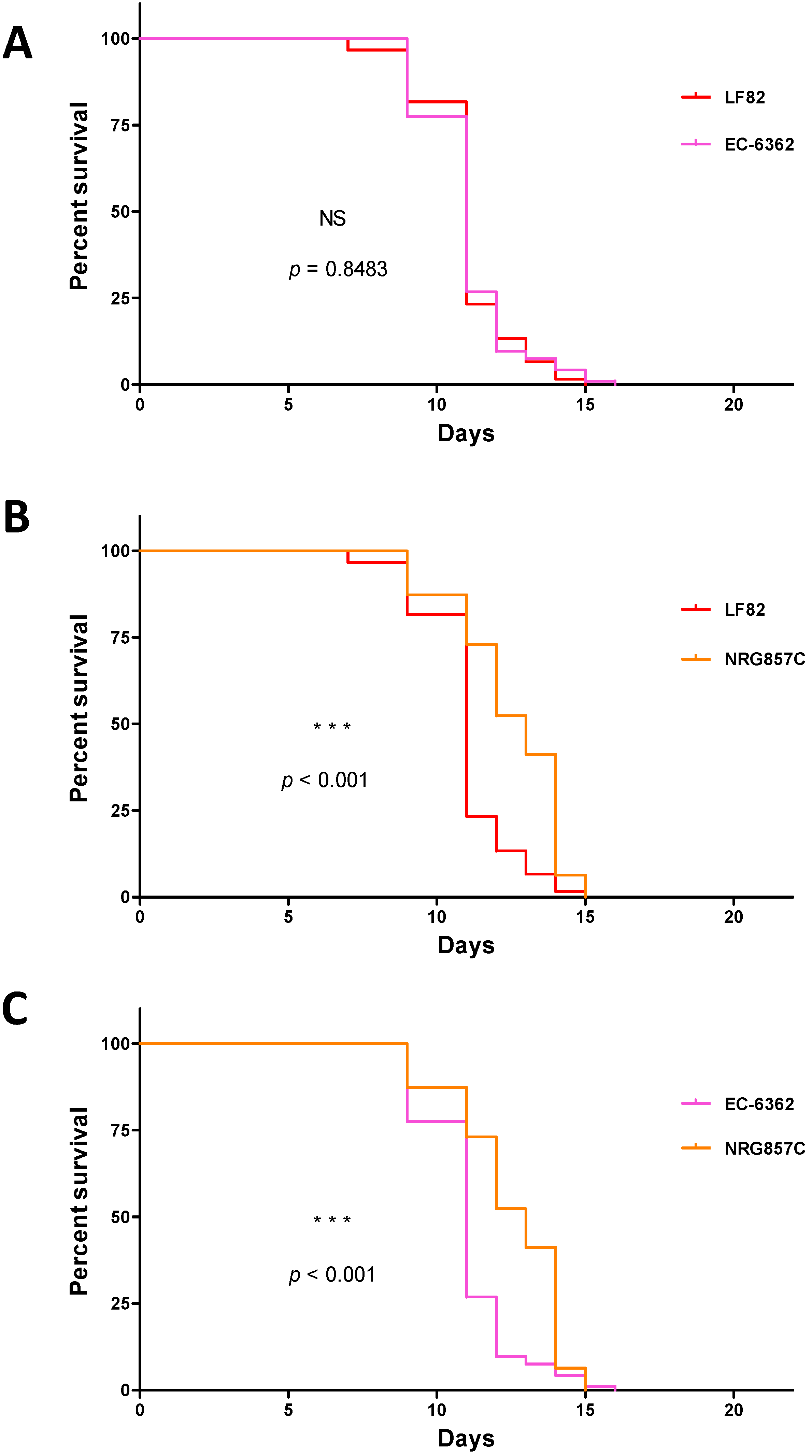
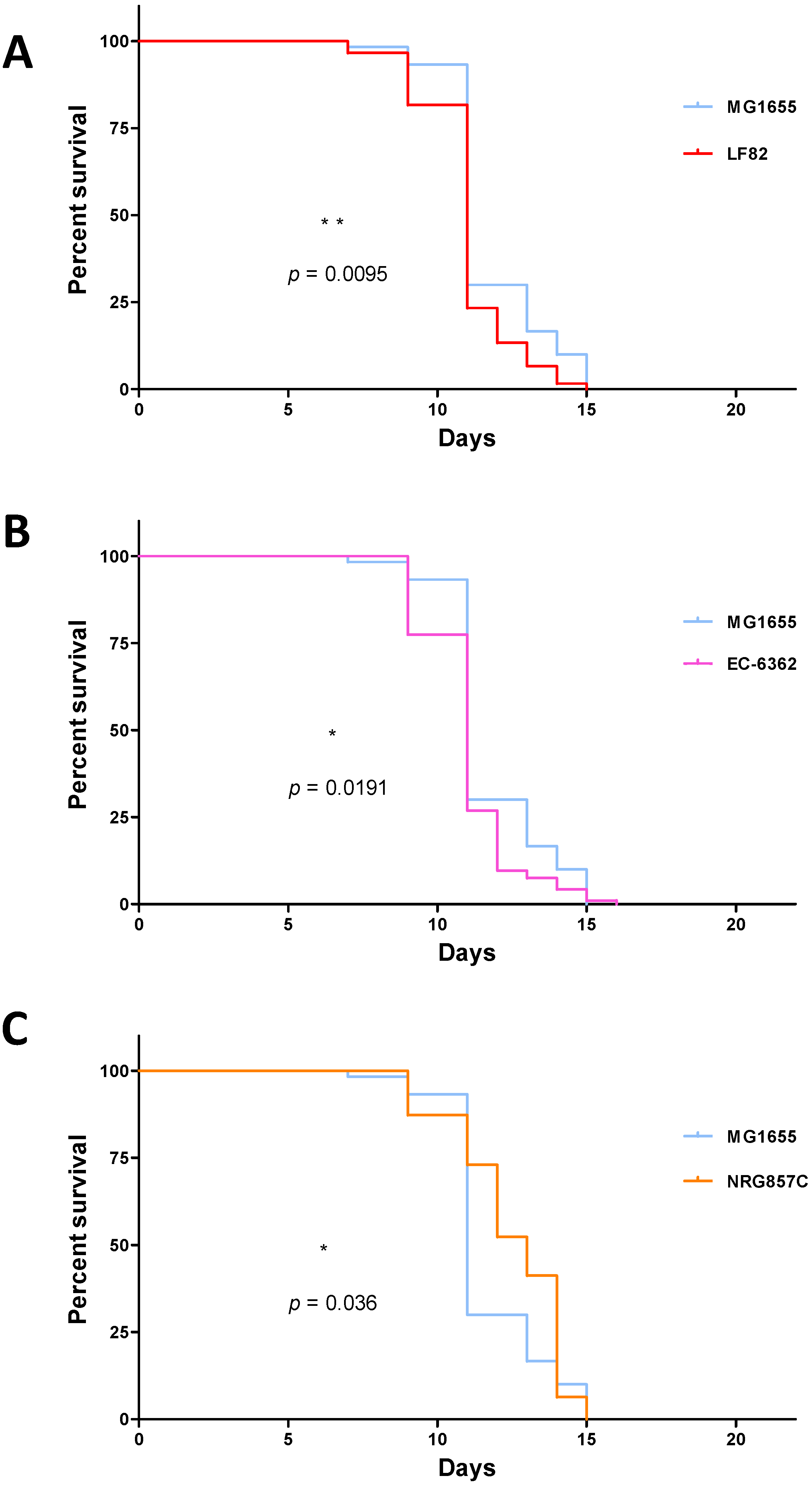
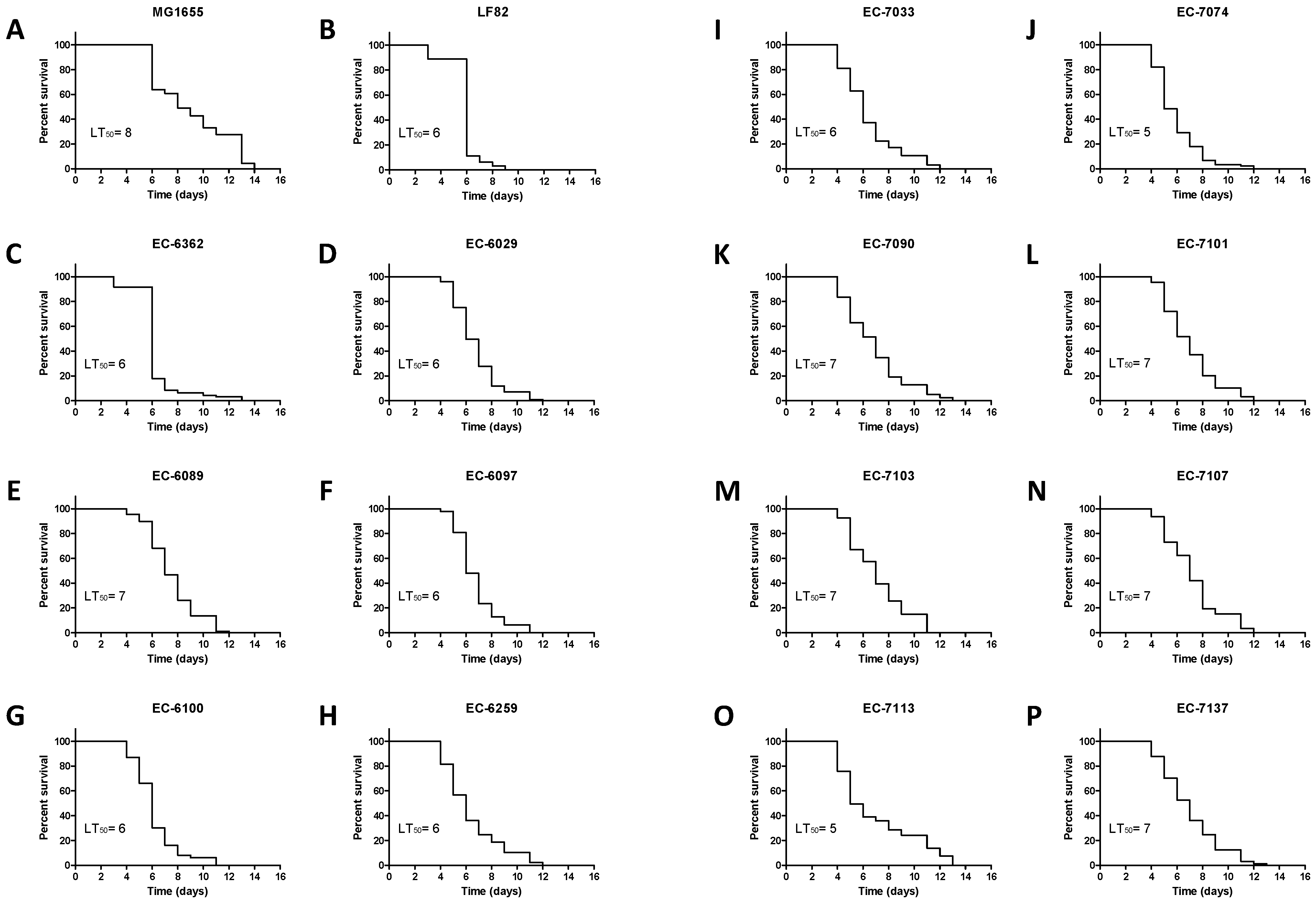
3.1. Strain-Specific E. coli Infection Impacts the Survival of C. elegans
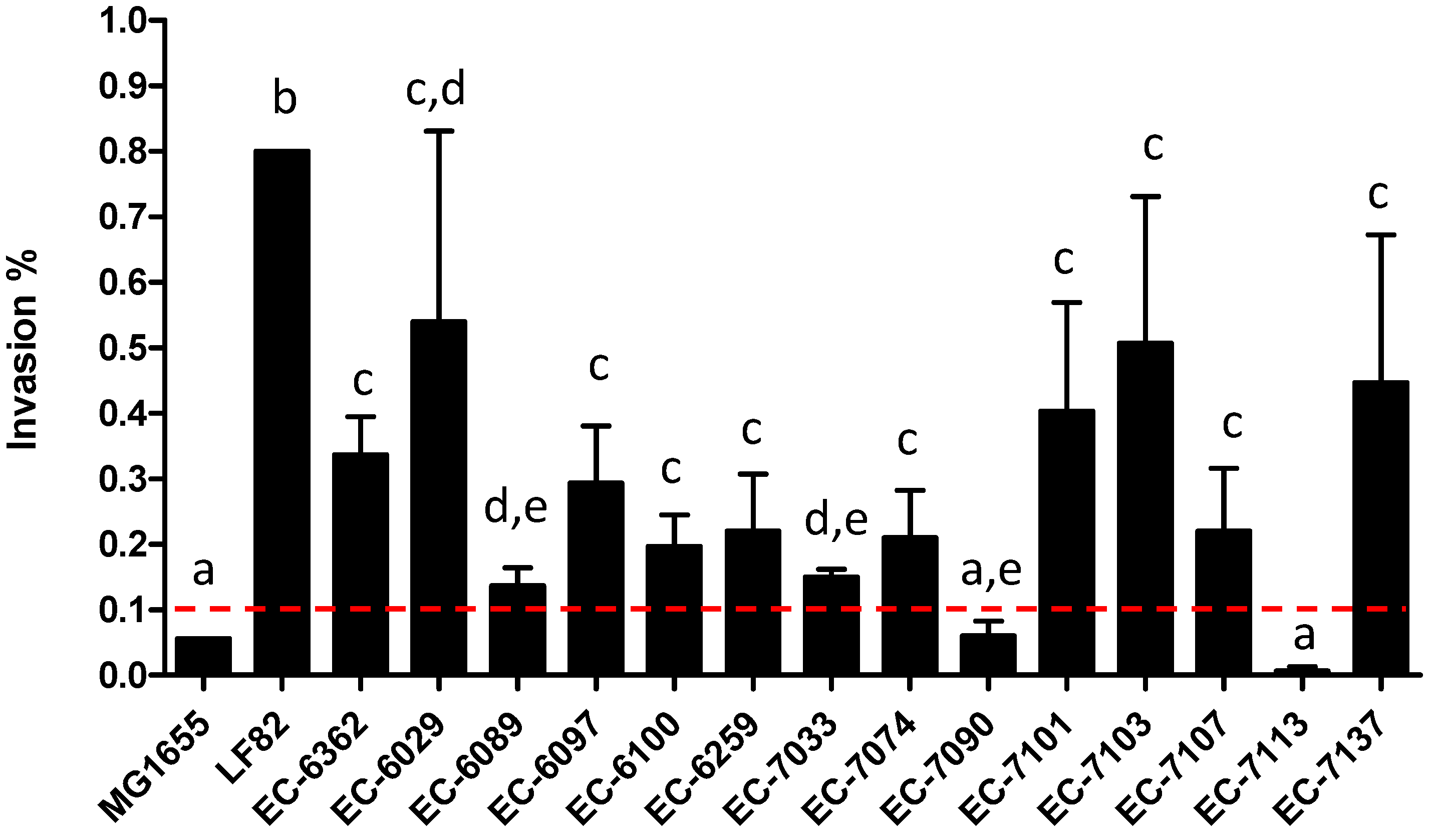
3.2. AIEC Strains Differ in Their Cell Invasion Properties In Vitro
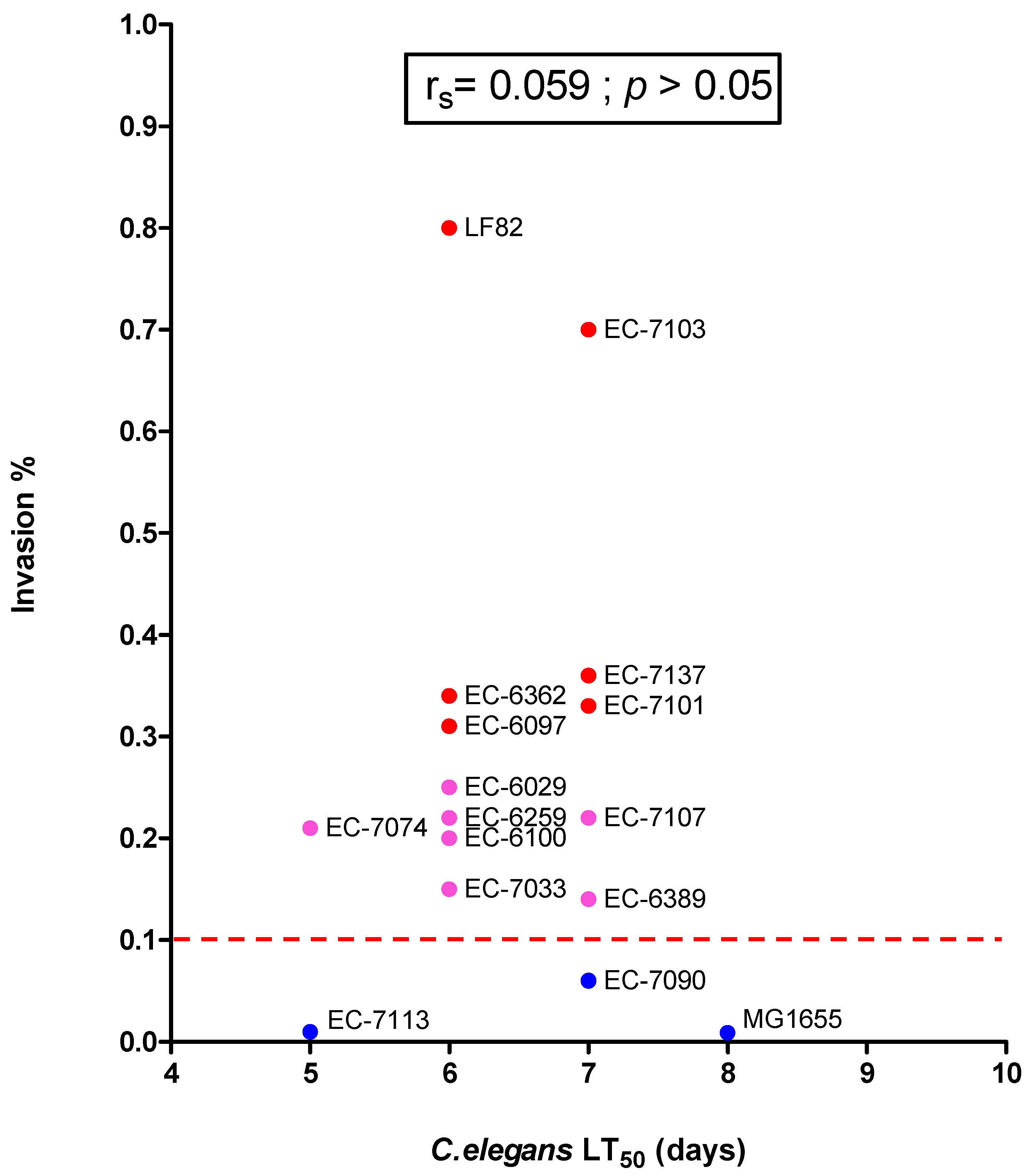
3.3. Assessment of the AIEC’s Pathogenicity in C. elegans
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Furey, T.S.; Sethupathy, P.; Sheikh, S.Z. Redefining the IBDs Using Genome-Scale Molecular Phenotyping. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 296–311. [Google Scholar] [CrossRef]
- Annese, V. Genetics and Epigenetics of IBD. Pharm. Res. 2020, 159, 104892. [Google Scholar] [CrossRef]
- Pascal, V.; Pozuelo, M.; Borruel, N.; Casellas, F.; Campos, D.; Santiago, A.; Martinez, X.; Varela, E.; Sarrabayrouse, G.; Machiels, K.; et al. A Microbial Signature for Crohn’s Disease. Gut 2017, 66, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. The Gut Microbiota Shapes Intestinal Immune Responses during Health and Disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef]
- Tamboli, C.P.; Neut, C.; Desreumaux, P.; Colombel, J.F. Dysbiosis in Inflammatory Bowel Disease. Gut 2004, 53, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darfeuille-Michaud, A.; Neut, C.; Barnich, N.; Lederman, E.; Di Martino, P.; Desreumaux, P.; Gambiez, L.; Joly, B.; Cortot, A.; Colombel, J.F. Presence of Adherent Escherichia coli Strains in Ileal Mucosa of Patients with Crohn’s Disease. Gastroenterology 1998, 115, 1405–1413. [Google Scholar] [CrossRef]
- Boudeau, J.; Glasser, A.L.; Masseret, E.; Joly, B.; Darfeuille-Michaud, A. Invasive Ability of an Escherichia coli Strain Isolated from the Ileal Mucosa of a Patient with Crohn’s Disease. Infect. Immun. 1999, 67, 4499–4509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmela, C.; Chevarin, C.; Xu, Z.; Torres, J.; Sevrin, G.; Hirten, R.; Barnich, N.; Ng, S.C.; Colombel, J.-F. Adherent-Invasive Escherichia coli in Inflammatory Bowel Disease. Gut 2018, 67, 574–587. [Google Scholar] [CrossRef]
- Martinez-Medina, M.; Aldeguer, X.; Lopez-Siles, M.; González-Huix, F.; López-Oliu, C.; Dahbi, G.; Blanco, J.E.; Blanco, J.; Garcia-Gil, L.J.; Darfeuille-Michaud, A. Molecular Diversity of Escherichia coli in the Human Gut: New Ecological Evidence Supporting the Role of Adherent-Invasive E. coli (AIEC) in Crohn’s Disease. Inflamm. Bowel. Dis. 2009, 15, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.W.; Dogan, B.; Rishniw, M.; Goldstein, R.E.; Klaessig, S.; McDonough, P.L.; German, A.J.; Yates, R.M.; Russell, D.G.; Johnson, S.E.; et al. Adherent and Invasive Escherichia coli Is Associated with Granulomatous Colitis in Boxer Dogs. Infect. Immun. 2006, 74, 4778–4792. [Google Scholar] [CrossRef] [Green Version]
- Drouet, M.; Vignal, C.; Singer, E.; Djouina, M.; Dubreuil, L.; Cortot, A.; Desreumaux, P.; Neut, C. AIEC Colonization and Pathogenicity: Influence of Previous Antibiotic Treatment and Preexisting Inflammation. Inflamm. Bowel. Dis. 2012, 18, 1923–1931. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Hasegawa, M.; Hayashi, A.; Caballero-Flores, G.; Alteri, C.J.; Lawley, T.D.; Kamada, N.; Núñez, G.; Inohara, N. Lipopolysaccharide O Structure of Adherent and Invasive Escherichia coli Regulates Intestinal Inflammation via Complement C3. PLoS Pathog. 2020, 16, e1008928. [Google Scholar] [CrossRef] [PubMed]
- Nadalian, B.; Yadegar, A.; Houri, H.; Olfatifar, M.; Shahrokh, S.; Asadzadeh Aghdaei, H.; Suzuki, H.; Zali, M.R. Prevalence of the Pathobiont Adherent-Invasive Escherichia coli and Inflammatory Bowel Disease: A Systematic Review and Meta-Analysis. J. Gastroenterol. Hepatol. 2021, 36, 852–863. [Google Scholar] [CrossRef] [PubMed]
- Desilets, M.; Deng, X.; Deng, X.; Rao, C.; Ensminger, A.W.; Krause, D.O.; Sherman, P.M.; Gray-Owen, S.D. Genome-Based Definition of an Inflammatory Bowel Disease-Associated Adherent-Invasive Escherichia coli Pathovar. Inflamm. Bowel. Dis. 2016, 22, 1992–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prorok-Hamon, M.; Friswell, M.K.; Alswied, A.; Roberts, C.L.; Song, F.; Flanagan, P.K.; Knight, P.; Codling, C.; Marchesi, J.R.; Winstanley, C.; et al. Colonic Mucosa-Associated Diffusely Adherent AfaC+ Escherichia coli Expressing LpfA and Pks Are Increased in Inflammatory Bowel Disease and Colon Cancer. Gut 2014, 63, 761–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, A.G.; Lundsmith, E.T.; Hamilton, K.E. Inflammation and Colorectal Cancer. Curr. Colorectal Cancer Rep. 2017, 13, 341–351. [Google Scholar] [CrossRef]
- Rahmouni, O.; Vignal, C.; Titécat, M.; Foligné, B.; Pariente, B.; Dubuquoy, L.; Desreumaux, P.; Neut, C. High Carriage of Adherent Invasive E. coli in Wildlife and Healthy Individuals. Gut Pathog. 2018, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Astley, D.J.; Masters, N.; Kuballa, A.; Katouli, M. Commonality of Adherent-Invasive Escherichia coli Isolated from Patients with Extraintestinal Infections, Healthy Individuals and the Environment. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 181–192. [Google Scholar] [CrossRef]
- DeJong, E.N.; Surette, M.G.; Bowdish, D.M.E. The Gut Microbiota and Unhealthy Aging: Disentangling Cause from Consequence. Cell Host Microbe 2020, 28, 180–189. [Google Scholar] [CrossRef]
- Kotlowski, R.; Bernstein, C.N.; Sepehri, S.; Krause, D.O. High Prevalence of Escherichia coli Belonging to the B2+D Phylogenetic Group in Inflammatory Bowel Disease. Gut 2007, 56, 669–675. [Google Scholar] [CrossRef] [Green Version]
- Mirsepasi-Lauridsen, H.C.; Vallance, B.A.; Krogfelt, K.A.; Petersen, A.M. Escherichia coli Pathobionts Associated with Inflammatory Bowel Disease. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef] [Green Version]
- Shaler, C.R.; Elhenawy, W.; Coombes, B.K. The Unique Lifestyle of Crohn’s Disease-Associated Adherent-Invasive Escherichia coli. J. Mol. Biol. 2019, 431, 2970–2981. [Google Scholar] [CrossRef] [PubMed]
- Shawki, A.; McCole, D.F. Mechanisms of Intestinal Epithelial Barrier Dysfunction by Adherent-Invasive Escherichia coli. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camprubí-Font, C.; Lopez-Siles, M.; Ferrer-Guixeras, M.; Niubó-Carulla, L.; Abellà-Ametller, C.; Garcia-Gil, L.J.; Martinez-Medina, M. Comparative Genomics Reveals New Single-Nucleotide Polymorphisms That Can Assist in Identification of Adherent-Invasive Escherichia coli. Sci. Rep. 2018, 8, 2695. [Google Scholar] [CrossRef] [PubMed]
- Camprubí-Font, C.; Ewers, C.; Lopez-Siles, M.; Martinez-Medina, M. Genetic and Phenotypic Features to Screen for Putative Adherent-Invasive Escherichia coli. Front. Microbiol. 2019, 10, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tawfik, A.; Knight, P.; Duckworth, C.A.; Pritchard, D.M.; Rhodes, J.M.; Campbell, B.J. Replication of Crohn’s Disease Mucosal E. coli Isolates inside Macrophages Correlates with Resistance to Superoxide and Is Dependent on Macrophage NF-Kappa B Activation. Pathogens 2019, 8, 74. [Google Scholar] [CrossRef] [Green Version]
- Martin, H.M.; Campbell, B.J.; Hart, C.A.; Mpofu, C.; Nayar, M.; Singh, R.; Englyst, H.; Williams, H.F.; Rhodes, J.M. Enhanced Escherichia coli Adherence and Invasion in Crohn’s Disease and Colon Cancer. Gastroenterology 2004, 127, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Gerbaba, T.K.; Green-Harrison, L.; Buret, A.G. Modeling Host-Microbiome Interactions in Caenorhabditis elegans. J. Nematol. 2017, 49, 348–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ermolaeva, M.A.; Schumacher, B. Insights from the Worm: The C. elegans Model for Innate Immunity. Semin. Immunol. 2014, 26, 303–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poupet, C.; Chassard, C.; Nivoliez, A.; Bornes, S. Caenorhabditis elegans, a Host to Investigate the Probiotic Properties of Beneficial Microorganisms. Front. Nutr. 2020, 7, 135. [Google Scholar] [CrossRef]
- Aballay, A.; Yorgey, P.; Ausubel, F.M. Salmonella Typhimurium Proliferates and Establishes a Persistent Infection in the Intestine of Caenorhabditis elegans. Curr. Biol. 2000, 10, 1539–1542. [Google Scholar] [CrossRef] [Green Version]
- Labrousse, A.; Chauvet, S.; Couillault, C.; Kurz, C.L.; Ewbank, J.J. Caenorhabditis elegans Is a Model Host for Salmonella Typhimurium. Curr. Biol. 2000, 10, 1543–1545. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Choe, J.; Kim, J.; Oh, S.; Park, S.; Kim, S.; Kim, Y. Heat-Killed Lactobacillus Spp. Cells Enhance Survivals of Caenorhabditis elegans against Salmonella and Yersinia Infections. Lett. Appl. Microbiol. 2015, 61, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Mellies, J.L.; Barron, A.M.S.; Haack, K.R.; Korson, A.S.; Oldridge, D.A. The Global Regulator Ler Is Necessary for Enteropathogenic Escherichia coli Colonization of Caenorhabditis elegans. Infect. Immun. 2006, 74, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anyanful, A.; Dolan-Livengood, J.M.; Lewis, T.; Sheth, S.; Dezalia, M.N.; Sherman, M.A.; Kalman, L.V.; Benian, G.M.; Kalman, D. Paralysis and Killing of Caenorhabditis elegans by Enteropathogenic Escherichia coli Requires the Bacterial Tryptophanase Gene. Mol. Microbiol. 2005, 57, 988–1007. [Google Scholar] [CrossRef]
- Chou, T.-C.; Chiu, H.-C.; Kuo, C.-J.; Wu, C.-M.; Syu, W.-J.; Chiu, W.-T.; Chen, C.-S. Enterohaemorrhagic Escherichia coli O157:H7 Shiga-like Toxin 1 Is Required for Full Pathogenicity and Activation of the P38 Mitogen-Activated Protein Kinase Pathway in Caenorhabditis elegans. Cell. Microbiol. 2013, 15, 82–97. [Google Scholar] [CrossRef]
- Schifano, E.; Marazzato, M.; Ammendolia, M.G.; Zanni, E.; Ricci, M.; Comanducci, A.; Goldoni, P.; Conte, M.P.; Uccelletti, D.; Longhi, C. Virulence Behavior of Uropathogenic Escherichia coli Strains in the Host Model Caenorhabditis elegans. Microbiologyopen 2019, 8, e00756. [Google Scholar] [CrossRef]
- Conte, M.P.; Aleandri, M.; Marazzato, M.; Conte, A.L.; Ambrosi, C.; Nicoletti, M.; Zagaglia, C.; Gambara, G.; Palombi, F.; De Cesaris, P.; et al. The Adherent/Invasive Escherichia coli Strain LF82 Invades and Persists in Human Prostate Cell Line RWPE-1, Activating a Strong Inflammatory Response. Infect. Immun. 2016, 84, 3105–3113. [Google Scholar] [CrossRef] [Green Version]
- Simonsen, K.T.; Nielsen, G.; Bjerrum, J.V.; Kruse, T.; Kallipolitis, B.H.; Møller-Jensen, J. A Role for the RNA Chaperone Hfq in Controlling Adherent-Invasive Escherichia coli Colonization and Virulence. PLoS ONE 2011, 6, e16387. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics Of Caenorhabditis elegans. Genetics 1974, 77, 71. [Google Scholar] [CrossRef]
- Samuel, B.S.; Rowedder, H.; Braendle, C.; Félix, M.-A.; Ruvkun, G. Caenorhabditis elegans Responses to Bacteria from Its Natural Habitats. Proc. Natl. Acad. Sci. USA 2016, 113, E3941–E3949. [Google Scholar] [CrossRef] [Green Version]
- Blattner, F.R.; Plunkett, G.; Bloch, C.A.; Perna, N.T.; Burland, V.; Riley, M.; Collado-Vides, J.; Glasner, J.D.; Rode, C.K.; Mayhew, G.F.; et al. The Complete Genome Sequence of Escherichia coli K-12. Science 1997, 277, 1453–1462. [Google Scholar] [CrossRef] [Green Version]
- Wassenaar, T.M. Insights from 100 Years of Research with Probiotic E. coli. Eur. J. Microbiol. Immunol. 2016, 6, 147–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnenborn, U. Escherichia coli Strain Nissle 1917-from Bench to Bedside and Back: History of a Special Escherichia coli Strain with Probiotic Properties. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [Green Version]
- Small, C.-L.N.; Reid-Yu, S.A.; McPhee, J.B.; Coombes, B.K. Persistent Infection with Crohn’s Disease-Associated Adherent-Invasive Escherichia coli Leads to Chronic Inflammation and Intestinal Fibrosis. Nat. Commun. 2013, 4, 1957. [Google Scholar] [CrossRef] [PubMed]
- Nash, J.H.; Villegas, A.; Kropinski, A.M.; Aguilar-Valenzuela, R.; Konczy, P.; Mascarenhas, M.; Ziebell, K.; Torres, A.G.; Karmali, M.A.; Coombes, B.K. Genome Sequence of Adherent-Invasive Escherichia coli and Comparative Genomic Analysis with Other E. coli Pathotypes. BMC Genom. 2010, 11, 667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreux, N.; Denizot, J.; Martinez-Medina, M.; Mellmann, A.; Billig, M.; Kisiela, D.; Chattopadhyay, S.; Sokurenko, E.; Neut, C.; Gower-Rousseau, C.; et al. Point Mutations in FimH Adhesin of Crohn’s Disease-Associated Adherent-Invasive Escherichia coli Enhance Intestinal Inflammatory Response. PLoS Pathog. 2013, 9, e1003141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Sordi, L.; Khanna, V.; Debarbieux, L. The Gut Microbiota Facilitates Drifts in the Genetic Diversity and Infectivity of Bacterial Viruses. Cell Host Microbe 2017, 22, 801–808.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasimatis, K.R.; Moerdyk-Schauwecker, M.J.; Phillips, P.C. Auxin-Mediated Sterility Induction System for Longevity and Mating Studies in Caenorhabditis elegans. G3 Genes Genomes Genet. 2018, 8, 2655–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilberger, B.; Baumanns, S.; Spieth, S.T.; Wenzel, U.; Eckert, G.P. Infertility Induced by Auxin in PX627 Caenorhabditis elegans Does Not Affect Mitochondrial Functions and Aging Parameters. Aging 2020, 12, 12268–12284. [Google Scholar] [CrossRef] [PubMed]
- Mayorgas, A.; Dotti, I.; Martínez-Picola, M.; Esteller, M.; Bonet-Rossinyol, Q.; Ricart, E.; Salas, A.; Martínez-Medina, M. A Novel Strategy to Study the Invasive Capability of Adherent-Invasive Escherichia coli by Using Human Primary Organoid-Derived Epithelial Monolayers. Front. Immunol. 2021, 12, 646906. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.A.; Barnich, N.; Sauvanet, P.; Darcha, C.; Gelot, A.; Darfeuille-Michaud, A. Crohn’s Disease-Associated Escherichia coli LF82 Aggravates Colitis in Injured Mouse Colon via Signaling by Flagellin. Inflamm. Bowel. Dis. 2008, 14, 1051–1060. [Google Scholar] [CrossRef]
- Chokr, D.; Cornu, M.; Neut, C.; Bortolus, C.; Charlet, R.; Desreumaux, P.; Speca, S.; Sendid, B. Adherent Invasive Escherichia coli (AIEC) Strain LF82, but Not Candida albicans, Plays a Profibrogenic Role in the Intestine. Gut Pathog. 2021, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Small, C.L.; Xing, L.; McPhee, J.B.; Law, H.T.; Coombes, B.K. Acute Infectious Gastroenteritis Potentiates a Crohn’s Disease Pathobiont to Fuel Ongoing Inflammation in the Post-Infectious Period. PLoS Pathog. 2016, 12, e1005907. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.T.; Dalmasso, G.; Müller, S.; Carrière, J.; Seibold, F.; Darfeuille-Michaud, A. Crohn’s Disease-Associated Adherent Invasive Escherichia coli Modulate Levels of MicroRNAs in Intestinal Epithelial Cells to Reduce Autophagy. Gastroenterology 2014, 146, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Dalmasso, G.; Nguyen, H.T.T.; Faïs, T.; Massier, S.; Barnich, N.; Delmas, J.; Bonnet, R. Crohn’s Disease-Associated Adherent-Invasive Escherichia coli Manipulate Host Autophagy by Impairing SUMOylation. Cells 2019, 8, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elhenawy, W.; Hordienko, S.; Gould, S.; Oberc, A.M.; Tsai, C.N.; Hubbard, T.P.; Waldor, M.K.; Coombes, B.K. High-Throughput Fitness Screening and Transcriptomics Identify a Role for a Type IV Secretion System in the Pathogenesis of Crohn’s Disease-Associated Escherichia coli. Nat. Commun. 2021, 12, 2032. [Google Scholar] [CrossRef]
- Viennois, E.; Bretin, A.; Dubé, P.E.; Maue, A.C.; Dauriat, C.J.G.; Barnich, N.; Gewirtz, A.T.; Chassaing, B. Dietary Emulsifiers Directly Impact Adherent-Invasive E.coli Gene Expression to Drive Chronic Intestinal Inflammation. Cell Rep. 2020, 33, 108229. [Google Scholar] [CrossRef] [PubMed]
- Vazeille, E.; Chassaing, B.; Buisson, A.; Dubois, A.; de Vallée, A.; Billard, E.; Neut, C.; Bommelaer, G.; Colombel, J.-F.; Barnich, N.; et al. GipA Factor Supports Colonization of Peyer’s Patches by Crohn’s Disease-Associated Escherichia coli. Inflamm. Bowel. Dis. 2016, 22, 68–81. [Google Scholar] [CrossRef] [Green Version]
- Gibold, L.; Garenaux, E.; Dalmasso, G.; Gallucci, C.; Cia, D.; Mottet-Auselo, B.; Faïs, T.; Darfeuille-Michaud, A.; Nguyen, H.T.T.; Barnich, N.; et al. The Vat-AIEC Protease Promotes Crossing of the Intestinal Mucus Layer by Crohn’s Disease-Associated Escherichia coli. Cell. Microbiol. 2016, 18, 617–631. [Google Scholar] [CrossRef]
- Sánchez-Diener, I.; Zamorano, L.; López-Causapé, C.; Cabot, G.; Mulet, X.; Peña, C.; Del Campo, R.; Cantón, R.; Doménech-Sánchez, A.; Martínez-Martínez, L.; et al. Interplay among Resistance Profiles, High-Risk Clones, and Virulence in the Caenorhabditis elegans Pseudomonas aeruginosa Infection Model. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sifri, C.D.; Begun, J.; Ausubel, F.M. The Worm Has Turned—Microbial Virulence Modeled in Caenorhabditis elegans. Trends Microbiol. 2005, 13, 119–127. [Google Scholar] [CrossRef]
- Sotto, A.; Lina, G.; Richard, J.-L.; Combescure, C.; Bourg, G.; Vidal, L.; Jourdan, N.; Etienne, J.; Lavigne, J.-P. Virulence Potential of Staphylococcus aureus Strains Isolated from Diabetic Foot Ulcers: A New Paradigm. Diabetes Care 2008, 31, 2318–2324. [Google Scholar] [CrossRef] [Green Version]
- Scott, E.; Holden-Dye, L.; O’Connor, V.; Wand, M.E. Intra Strain Variation of the Effects of Gram-Negative ESKAPE Pathogens on Intestinal Colonization, Host Viability, and Host Response in the Model Organism Caenorhabditis elegans. Front. Microbiol. 2019, 10, 3113. [Google Scholar] [CrossRef] [PubMed]
- Browning, D.F.; Wells, T.J.; França, F.L.S.; Morris, F.C.; Sevastsyanovich, Y.R.; Bryant, J.A.; Johnson, M.D.; Lund, P.A.; Cunningham, A.F.; Hobman, J.L.; et al. Laboratory Adapted Escherichia coli K-12 Becomes a Pathogen of Caenorhabditis elegans upon Restoration of O Antigen Biosynthesis. Mol. Microbiol. 2013, 87, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Rosovitz, M.J.; Myers, G.S.A.; Mongodin, E.F.; Fricke, W.F.; Gajer, P.; Crabtree, J.; Sebaihia, M.; Thomson, N.R.; Chaudhuri, R.; et al. The Pangenome Structure of Escherichia coli: Comparative Genomic Analysis of E. coli Commensal and Pathogenic Isolates. J. Bacteriol. 2008, 190, 6881–6893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavigne, J.-P.; Blanc-Potard, A.-B.; Bourg, G.; Moreau, J.; Chanal, C.; Bouziges, N.; O’callaghan, D.; Sotto, A. Virulence Genotype and Nematode-Killing Properties of Extra-Intestinal Escherichia coli Producing CTX-M Beta-Lactamases. Clin. Microbiol. Infect. 2006, 12, 1199–1206. [Google Scholar] [CrossRef] [PubMed]
- Antão, E.-M.; Wieler, L.H.; Ewers, C. Adhesive Threads of Extraintestinal Pathogenic Escherichia coli. Gut Pathog. 2009, 1, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Moon, Y. Worm-Based Alternate Assessment of Probiotic Intervention against Gut Barrier Infection. Nutrients 2019, 11, 2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migliore, F.; Macchi, R.; Landini, P.; Paroni, M. Phagocytosis and Epithelial Cell Invasion by Crohn’s Disease-Associated Adherent-Invasive Escherichia coli Are Inhibited by the Anti-Inflammatory Drug 6-Mercaptopurine. Front. Microbiol. 2018, 9, 964. [Google Scholar] [CrossRef] [PubMed]
- Aygun, H.; Karamese, M.; Ozic, C.; Uyar, F. The Effects of Mucosal Media on Some Pathogenic Traits of Crohn’s Disease-Associated Escherichia coli LF82. Future Microbiol. 2018, 13, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Agus, A.; Richard, D.; Faïs, T.; Vazeille, E.; Chervy, M.; Bonnin, V.; Dalmasso, G.; Denizot, J.; Billard, E.; Bonnet, R.; et al. Propionate Catabolism by CD-Associated Adherent-Invasive E. coli Counteracts Its Anti-Inflammatory Effect. Gut Microbes 2021, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Dogan, B.; Guo, C.; Herlekar, D.; Stewart, K.; Scherl, E.J.; Simpson, K.W. Short Chain Fatty Acids Modulate the Growth and Virulence of Pathosymbiont Escherichia coli and Host Response. Antibiotics 2020, 9, 462. [Google Scholar] [CrossRef] [PubMed]
- Ormsby, M.J.; Johnson, S.A.; Carpena, N.; Meikle, L.M.; Goldstone, R.J.; McIntosh, A.; Wessel, H.M.; Hulme, H.E.; McConnachie, C.C.; Connolly, J.P.R.; et al. Propionic Acid Promotes the Virulent Phenotype of Crohn’s Disease-Associated Adherent-Invasive Escherichia coli. Cell Rep. 2020, 30, 2297–2305.e5. [Google Scholar] [CrossRef] [Green Version]
- Lepanto, M.S.; Rosa, L.; Cutone, A.; Scotti, M.J.; Conte, A.L.; Marazzato, M.; Zagaglia, C.; Longhi, C.; Berlutti, F.; Musci, G.; et al. Bovine Lactoferrin Pre-Treatment Induces Intracellular Killing of AIEC LF82 and Reduces Bacteria-Induced DNA Damage in Differentiated Human Enterocytes. Int. J. Mol. Sci. 2019, 20, 5666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickerson, K.P.; McDonald, C. Crohn’s Disease-Associated Adherent-Invasive Escherichia coli Adhesion Is Enhanced by Exposure to the Ubiquitous Dietary Polysaccharide Maltodextrin. PLoS ONE 2012, 7, e52132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anyanful, A.; Easley, K.A.; Benian, G.M.; Kalman, D. Conditioning Protects C. elegans from Lethal Effects of Enteropathogenic E. coli by Activating Genes That Regulate Lifespan and Innate Immunity. Cell Host Microbe 2009, 5, 450–462. [Google Scholar] [CrossRef] [Green Version]
- JebaMercy, G.; Balamurugan, K. Effects of Sequential Infections of Caenorhabditis elegans with Staphylococcus aureus and Proteus mirabilis. Microbiol. Immunol. 2012, 56, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, T.; Yasui, C.; Hoshino, K.; Arikawa, K.; Nishikawa, Y. Influence of Lactic Acid Bacteria on Longevity of Caenorhabditis elegans and Host Defense against Salmonella Enterica Serovar Enteritidis. Appl. Environ. Microbiol. 2007, 73, 6404–6409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martorell, P.; Alvarez, B.; Llopis, S.; Navarro, V.; Ortiz, P.; Gonzalez, N.; Balaguer, F.; Rojas, A.; Chenoll, E.; Ramón, D.; et al. Heat-Treated Bifidobacterium longum CECT-7347: A Whole-Cell Postbiotic with Antioxidant, Anti-Inflammatory, and Gut-Barrier Protection Properties. Antioxidants 2021, 10, 536. [Google Scholar] [CrossRef]
- Kwon, G.; Lee, J.; Lim, Y.-H. Dairy Propionibacterium Extends the Mean Lifespan of Caenorhabditis elegans via Activation of the Innate Immune System. Sci. Rep. 2016, 6, 31713. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Pooranachithra, M.; Balamurugan, K.; Goel, G. Probiotic Mediated Colonization Resistance against E. coli Infection in Experimentally Challenged Caenorhabditis elegans. Microb. Pathog. 2019, 127, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Grompone, G.; Martorell, P.; Llopis, S.; González, N.; Genovés, S.; Mulet, A.P.; Fernández-Calero, T.; Tiscornia, I.; Bollati-Fogolín, M.; Chambaud, I.; et al. Anti-Inflammatory Lactobacillus rhamnosus CNCM I-3690 Strain Protects against Oxidative Stress and Increases Lifespan in Caenorhabditis elegans. PLoS ONE 2012, 7, e52493. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Sousa Figueiredo, M.B.; Pradel, E.; George, F.; Mahieux, S.; Houcke, I.; Pottier, M.; Fradin, C.; Neut, C.; Daniel, C.; Bongiovanni, A.; et al. Adherent-Invasive and Non-Invasive Escherichia coli Isolates Differ in Their Effects on Caenorhabditis elegans’ Lifespan. Microorganisms 2021, 9, 1823. https://doi.org/10.3390/microorganisms9091823
de Sousa Figueiredo MB, Pradel E, George F, Mahieux S, Houcke I, Pottier M, Fradin C, Neut C, Daniel C, Bongiovanni A, et al. Adherent-Invasive and Non-Invasive Escherichia coli Isolates Differ in Their Effects on Caenorhabditis elegans’ Lifespan. Microorganisms. 2021; 9(9):1823. https://doi.org/10.3390/microorganisms9091823
Chicago/Turabian Stylede Sousa Figueiredo, Maria Beatriz, Elizabeth Pradel, Fanny George, Séverine Mahieux, Isabelle Houcke, Muriel Pottier, Chantal Fradin, Christel Neut, Catherine Daniel, Antonino Bongiovanni, and et al. 2021. "Adherent-Invasive and Non-Invasive Escherichia coli Isolates Differ in Their Effects on Caenorhabditis elegans’ Lifespan" Microorganisms 9, no. 9: 1823. https://doi.org/10.3390/microorganisms9091823