
Bacteria, Fungi and Microalgae for the Bioremediation of Marine Sediments Contaminated by Petroleum Hydrocarbons in the Omics Era

 ,
, 
Abstract
:1. Introduction
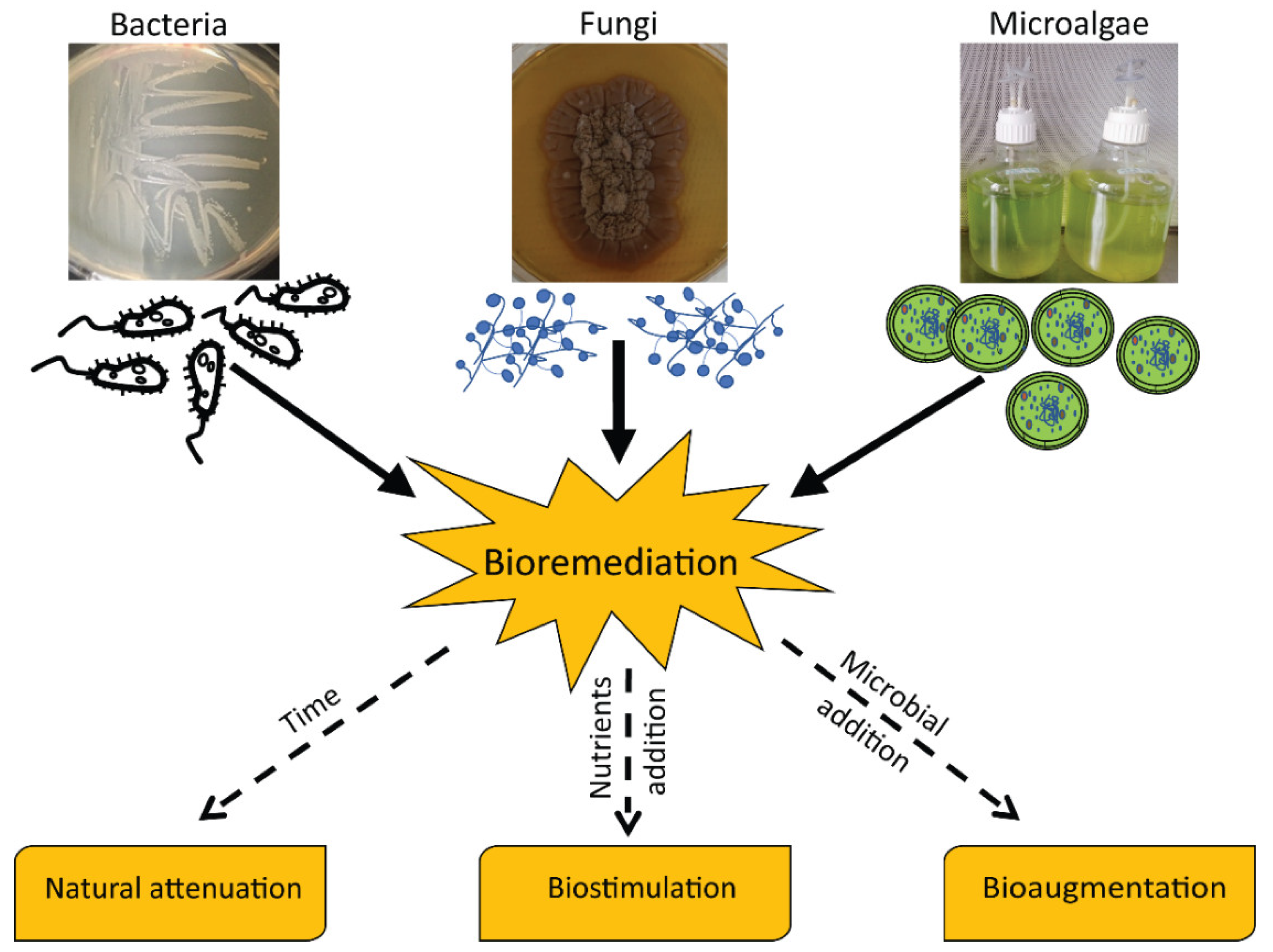
2. Factors Affecting the Bioremediation of Sediments Polluted by Petroleum Hydrocarbons
3. Bacterial-Mediated Degradation of Petroleum Hydrocarbons
4. Fungi-Mediated Degradation of Petroleum Hydrocarbons
5. Microalgae Involved in Hydrocarbon Removal
6. Towards an Omics Bioremediation Approach
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wuana, R.A.; Okieimen, F.E. Heavy Metals in Contaminated Soil: A Review of Sources, Chemistry, Risks and Best Available Strategies for Remediation. ISRN Ecol. 2011, 2011, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Salem, D.M.S.A.; Khaled, A.; El Nemr, A. Assessment of pesticides and polychlorinated biphenyls (PCBs) in sediments of the Egyptian Mediterranean Coast. Egypt. J. Aquat. Res. 2013, 39, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Chekroun, K.B.; Sánchez, E.; Baghour, M. The role of algae in bioremediation of organic pollutants. Int. Res. J. Public Environ. Health 2014, 1, 19–32. [Google Scholar]
- Zheng, B.; Zhao, X.; Liu, L.; Li, Z.; Lei, K.; Zhang, L.; Qin, Y.; Gan, Z.; Gao, S.; Jiao, L. Effects of hydrodynamics on the distribution of trace persistent organic pollutants and macrobenthic communities in Bohai Bay. Chemosphere 2011, 84, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Farrington, J.W.; Takada, H. Persistent organic pollutants (POPs), polycyclic aromatic hydrocarbons (PAHs), and plastics: Examples of the status, trend, and cycling of organic chemicals of environmental concern in the ocean. Oceanography 2014, 27, 196–213. [Google Scholar] [CrossRef]
- Moazampour, S.R.; Nabavi, S.M.B.; Roozbahani, M.M.; Khodadadi, M. Determination of total petroleum hydrocarbons and selected heavy metal (Pb, CO, V, Ni) concentration levels in surficial sediments of the Arvand River Estuary and their impact on benthic macroinvertebrates assemblages. Int. J. Environ. Anal. Chem. 2021. [Google Scholar] [CrossRef]
- Halanych, K.M.; Ainsworth, C.H.; Cordes, E.E.; Dodge, R.E.; Huettel, M.; Mendelssohn, I.A.; Murawski, S.A.; Paris-Limouzy, C.B.; Schwing, P.T.; Shaw, R.F.; et al. Effects of petroleum by-products and dispersants on ecosystems. Oceanography 2021, 34, 152–163. [Google Scholar] [CrossRef]
- Crini, G.; Lichtfouse, E. Wastewater treatment: An overview.Green Adsorbents for Pollutant Removal. Springer Nat. 2018, 18, 1–22. [Google Scholar]
- Peng, W.; Li, X.; Xiao, S.; Fan, W. Review of remediation technologies for sediments contaminated by heavy metals. J. Soils Sediments 2018, 18, 1701–1719. [Google Scholar] [CrossRef]
- Lofrano, G.; Libralato, G.; Minetto, D.; De Gisi, S.; Todaro, F.; Conte, B.; Calabrò, D.; Quatraro, L.; Notarnicola, M. In situ remediation of contaminated marinesediment: An overview. Environ. Sci. Pollut. Res. 2017, 24, 5189–5206. [Google Scholar] [CrossRef] [Green Version]
- European Marine Strategy Framework. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000. 2000. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32000L0060 (accessed on 5 August 2021).
- Megharaj, M.; Naidu, R. Soil and brownfield bioremediation. Microb. Biotechnol. 2017, 10, 1244–1249. [Google Scholar] [CrossRef]
- Brar, A.; Kumar, M.; Vivekanand, V.; Pareek, N. Photoautotrophic microorganisms and bioremediation of industrial effluents: Current status and future prospects. 3 Biotech 2017, 7, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beolchini, F.; Dell’Anno, A.; De Propris, L.; Ubaldini, S.; Cerrone, F.; Danovaro, R. Auto- and heterotrophic acidophilic bacteria enhance the bioremediation efficiency of sediments contaminated by heavy metals. Chemosphere 2009, 74, 1321–1326. [Google Scholar] [CrossRef] [PubMed]
- Das, N.; Chandran, P. Microbial Degradation of Petroleum Hydrocarbon Contaminants: An Overview. Biotechnol. Res. Int. 2011, 2011, 941810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, G.O.; Fufeyin, P.T.; Okoro, S.E.; Ehinomen, I.; Biology, E. Bioremediation, Biostimulation and Bioaugmention: A Review. Int. J. Environ. Bioremediation Biodegrad. 2015, 3, 28–39. [Google Scholar] [CrossRef]
- Fodelianakis, S.; Antoniou, E.; Mapelli, F.; Magagnini, M.; Nikolopoulou, M.; Marasco, R.; Barbato, M.; Tsiola, A.; Tsikopoulou, I.; Giaccaglia, L.; et al. Allochthonous bioaugmentation in ex situ treatment of crude oil-polluted sediments in the presence of an effective degrading indigenous microbiome. J. Hazard. Mater. 2015, 287, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Daccò, C.; Girometta, C.; Asemoloye, M.D.; Carpani, G.; Picco, A.M.; Tosi, S. Key fungal degradation patterns, enzymes and their applications for the removal of aliphatic hydrocarbons in polluted soils: A review. Int. Biodeterior. Biodegrad. 2020, 147, 104866. [Google Scholar] [CrossRef]
- Dell’Anno, F.; Brunet, C.; van Zyl, L.J.; Trindade, M.; Golyshin, P.N.; Dell’anno, A.; Ianora, A.; Sansone, C. Degradation of hydrocarbons and heavy metal reduction by marine bacteria in highly contaminated sediments. Microorganisms 2020, 8, 1402. [Google Scholar] [CrossRef]
- Dell’Anno, F.; Rastelli, E.; Tangherlini, M.; Corinaldesi, C.; Brunet, C.; Balzano, S.; Ianora, A.; Musco, L.; Dell’Anno, A. Highly contaminated marine sediments can host rare bacterial taxa potentially useful for bioremediation. Front. Microbiol. 2021, 12, 584850. [Google Scholar] [CrossRef]
- Garbisu, C.; Garaiyurrebaso, O.; Epelde, L.; Grohmann, E. Plasmid-Mediated Bioaugmentation for the Bioremediation of Contaminated Soils. Front. Microbiol. 2017, 8, 1966. [Google Scholar] [CrossRef] [Green Version]
- Ojuederie, O.B.; Babalola, O.O. Microbial and plant-assisted bioremediation of heavy metal polluted environments: A review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Liu, W.; Tian, S.; Wang, W.; Qi, Q.; Jiang, P.; Gao, X.; Li, F.; Li, H.; Yu, H. Petroleum Hydrocarbon-Degrading Bacteria for the Remediation of Oil Pollution Under Aerobic Conditions: A Perspective Analysis. Front. Microbiol. 2018, 9, 2885. [Google Scholar] [CrossRef] [PubMed]
- Yakimov, M.M.; Timmis, K.N.; Golyshin, P.N. Obligate oil-degrading marine bacteria. Curr. Opin. Biotechnol. 2007, 18, 257–266. [Google Scholar] [CrossRef]
- Morel, M.; Meux, E.; Mathieu, Y.; Thuillier, A.; Chibani, K.; Harvengt, L.; Jacquot, J.P.; Gelhaye, E. Xenomic networks variability and adaptation traits in wood decaying fungi. Microb. Biotechnol. 2013, 6, 248–263. [Google Scholar] [CrossRef] [PubMed]
- Durairaj, P.; Malla, S.; Nadarajan, S.P.; Lee, P.G.; Jung, E.; Park, H.H.; Kim, B.G.; Yun, H. Fungal cytochrome P450 monooxygenases of Fusarium oxysporum for the synthesis of ω-hydroxy fatty acids in engineered Saccharomyces cerevisiae. Microb. Cell Factories 2015, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Lladó, S.; Covino, S.; Solanas, A.M.; Viñas, M.; Petruccioli, M.; D’annibale, A. Comparative assessment of bioremediation approaches to highly recalcitrant PAH degradation in a real industrial polluted soil. J. Hazard. Mater. 2013, 248–249, 407–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaji, V.; Arulazhagan, P.; Ebenezer, P. Enzymatic bioremediation of polyaromatic hydrocarbons by fungal consortia enriched from petroleum contaminated soil and oil seeds. J. Environ. Biol. 2014, 35, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.T.; Lee, J.F.; Liu, K.H.; Liao, Y.F.; Yang, V. Immobilization of fungal laccase onto a nonionic surfactant-modified clay material: Application to PAH degradation. Environ. Sci. Pollut. Res. 2016, 23, 4024–4035. [Google Scholar] [CrossRef]
- Lei, A.P.; Hu, Z.L.; Wong, Y.S.; Tam, N.F.Y. Removal of fluoranthene and pyrene by different microalgal species. Bioresour. Technol. 2007, 98, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Takáčová, A.; Smolinská, M.; Ryba, J.; Mackuľak, T.; Jokrllová, J.; Hronec, P.; Čík, G. Biodegradation of Benzo[a]Pyrene through the use of algae. Cent. Eur. J. Chem. 2014, 12, 1133–1143. [Google Scholar] [CrossRef]
- De Llasera, M.P.G.; Olmos-Espejel, J.J.; Díaz-Flores, G.; Montaño-Montiel, A. Biodegradation of benzo(a)pyrene by two freshwater microalgae Selenastrum capricornutum and Scenedesmus acutus: A comparative study useful for bioremediation. Environ. Sci. Pollut. Res. 2016, 23, 3365–3375. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current state of knowledge in microbial degradation of polycyclic aromatic hydrocarbons (PAHs): A review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, R.; Khardenavis, A.A.; Purohit, H.J. Diverse Metabolic Capacities of Fungi for Bioremediation. Indian J. Microbiol. 2016, 56, 247–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Pohnert, G.; Wei, D. Extracellular metabolites from industrial microalgae and their biotechnological potential. Mar. Drugs 2016, 14, 191. [Google Scholar] [CrossRef] [PubMed]
- Casillo, A.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Exopolysaccharides from marine and marine extremophilic bacteria: Structures, properties, ecological roles and applications. Mar. Drugs 2018, 16, 69. [Google Scholar] [CrossRef] [Green Version]
- Perales-Vela, H.V.; Peña-Castro, J.M.; Cañizares-Villanueva, R.O. Heavy metal detoxification in eukaryotic microalgae. Chemosphere 2006, 64, 1–10. [Google Scholar] [CrossRef]
- Sharma, R.; Bhardwaj, R.; Handa, N.; Gautam, V.; Kohli, S.K.; Bali, S.; Kaur, P.; Thukral, A.K.; Arora, S.; Ohri, P.; et al. Responses of Phytochelatins and Metallothioneins in Alleviation of Heavy Metal Stress in Plants: An Overview. In Plant Metal Interaction: Emerging Remediation Techniques, 1st ed.; Amar, P., Ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 263–283. [Google Scholar] [CrossRef]
- Khullar, S.; Reddy, M.S. Cadmium and arsenic responses in the ectomycorrhizal fungus Laccaria bicolor: Glutathione metabolism and its role in metal(loid) homeostasis. Environ. Microbiol. Rep. 2019, 11, 53–61. [Google Scholar] [CrossRef]
- Lozada, M.; Marcos, M.S.; Commendatore, M.G.; Gil, M.N.; Dionisi, H.M. The bacterial community structure of hydrocarbon-polluted marine environments as the basis for the definition of an ecological index of hydrocarbon exposure. Microbes Environ. 2014, 29, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Tangherlini, M.; Corinaldesi, C.; Rastelli, E.; Musco, L.; Armiento, G.; Danovaro, R.; Dell’Anno, A. Chemical contamination can promote turnover diversity of benthic prokaryotic assemblages: The case study of the Bagnoli-Coroglio bay (southern Tyrrhenian Sea). Mar. Environ. Res. 2020, 160, 105040. [Google Scholar] [CrossRef]
- Mao, J.; Luo, Y.; Teng, Y.; Li, Z. Bioremediation of polycyclic aromatic hydrocarbon-contaminated soil by a bacterial consortium and associated microbial community changes. Int. Biodeterior. Biodegrad. 2012, 70, 141–147. [Google Scholar] [CrossRef]
- Abdel-Shafy, H.I.; Mansour, M.S.M. A review on polycyclic aromatic hydrocarbons: Source, environmental impact, effect on human health and remediation. Egypt. J. Pet. 2016, 25, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Azubuike, C.C.; Chikere, C.B.; Okpokwasili, G.C. Bioremediation techniques–classification based on site of application: Principles, advantages, limitations and prospects. World J. Microbiol. Biotechnol. 2016, 32, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.W.C.; Lai, K.M.; Wan, C.K.; Ma, K.K.; Fang, M. Isolation and optimization of PAH-degradative bacteria from contaminated soil for PAHs bioremediation. Water Air Soil Pollut. 2002, 139, 1–13. [Google Scholar] [CrossRef]
- Da Silva, A.C.; de Oliveira, F.J.; Bernardes, D.S.; de França, F.P. Bioremediation of marine sediments impacted by petroleum. Appl. Biochem. Biotechnol. 2009, 153, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Perdigão, R.; Almeida, C.M.R.; Santos, F.; Carvalho, M.F.; Mucha, A.P. Optimization of an autochthonous bacterial consortium obtained from beach sediments for bioremediation of petroleum hydrocarbons. Water 2021, 13, 66. [Google Scholar] [CrossRef]
- Gros, J.; Reddy, C.M.; Aeppli, C.; Nelson, R.K.; Carmichael, C.A.; Arey, J.S. Resolving biodegradation patterns of persistent saturated hydrocarbons in weathered oil samples from the Deepwater Horizon disaster. Environ. Sci. Technol. 2014, 48, 1628–1637. [Google Scholar] [CrossRef]
- Bagby, S.C.; Reddy, C.M.; Aeppli, C.; Fisher, G.B.; Valentine, D.L. Persistence and biodegradation of oil at the ocean floor following Deepwater Horizon. Proc. Natl. Acad. Sci. USA 2017, 114, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Salazar, J.; Loza, A.; Ornelas-Ocampo, K.; Gutierrez-Rios, R.M.; Pardo-López, L. Bacteria From the Southern Gulf of Mexico: Baseline, Diversity, Hydrocarbon-Degrading Potential and Future Applications. Front. Mar. Sci. 2021, 8, 625477. [Google Scholar] [CrossRef]
- Zaki, M.S.; Authman, M.M.N.; Abbas, H.H.H. Bioremediation of Petroleum Contaminants in Aquatic Environments. Life Sci. J. 2015, 12, 109–121. [Google Scholar] [CrossRef]
- Alexander, M. Biodegradation: Problems of molecular recalcitrance and microbial fallibility. Adv. Appl. Microbiol. 1965, 7, 35–80. [Google Scholar]
- Leahy, J.; Colwell, R. Microbial degradation of hydrocarbons in the environment. Microbiol. Rev. 1990, 54, 305–315. [Google Scholar] [CrossRef]
- Slater, G.; White, H.; Eglinton, T.; Reddy, C. Determination of microbial carbon sources in petroleum contaminated sediments using molecular 14C analysis. Environ. Sci. Technol. 2005, 39, 2552–2558. [Google Scholar] [CrossRef]
- McKew, B.A.; Coulon, F.; Yakimov, M.M.; Denaro, R.; Genovese, M.; Smith, C.J.; Osborn, A.M.; Timmis, K.N.; McGenity, T.J. Efficacy of intervention strategies for bioremediation of crude oil in marine systems and effects on indigenous hydrocarbonoclastic bacteria. Environ. Microbiol. 2007, 9, 1562–1571. [Google Scholar] [CrossRef] [PubMed]
- Calvo, C.; Manzanera, M.; Silva-Castro, G.A.; Uad, I.; González-López, J. Application of bioemulsifiers in soil oil bioremediation processes. Future prospects. Sci. Total Environ. 2009, 407, 3634–3640. [Google Scholar] [CrossRef] [PubMed]
- McKew, B.A.; Coulon, F.; Osborn, A.M.; Timmis, K.N.; McGenity, T.J. Determining the identity and roles of oil-metabolizing marine bacteria from the Thames estuary, UK. Environ. Microbiol. 2007, 9, 165–176. [Google Scholar] [CrossRef]
- Evans, F.F.; Rosado, A.S.; Sebastián, G.V.; Casella, R.; Machado, P.L.O.A.; Holmström, C.; Kjelleberg, S.; Van Elsas, J.D.; Seldin, L. Impact of oil contamination and biostimulation on the diversity of indigenous bacterial communities in soil microcosms. FEMS Microbiol. Ecol. 2004, 49, 295–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maculay, B.M. Understanding the behaviour of oil-degrading micro-organisms to enhance the microbial remediation of spilled petroleum. Appl. Ecol. Environ. Res. 2014, 13, 247–262. [Google Scholar] [CrossRef]
- Yang, S.Z.; Jin, H.J.; Wei, Z.; He, R.X.; Ji, Y.J.; Li, X.M.; Yu, S.P. Bioremediation of Oil Spills in Cold Environments: A Review. Pedosphere 2009, 19, 371–381. [Google Scholar] [CrossRef]
- Ahmad, M.; Yang, Q.; Zhang, Y.; Ling, J.; Sajjad, W.; Qi, S.; Zhou, W.; Zhang, Y.; Lin, X.; Zhang, Y.; et al. The distinct response of phenanthrene enriched bacterial consortia to different PAHs and their degradation potential: A mangrove sediment microcosm study. J. Hazard. Mater. 2019, 380, 120863. [Google Scholar] [CrossRef]
- Ayangbenro, A.S.; Babalola, O.O. A new strategy for heavy metal polluted environments: A review of microbial biosorbents. Int. J. Environ. Res. Public Health 2017, 14, 94. [Google Scholar] [CrossRef]
- Durán, R.E.; Méndez, V.; Rodríguez-castro, L.; Barra-sanhueza, B.; Salvà-serra, F.; Moore, E.R.B.; Castro-nallar, E.; Seeger, M. Genomic and Physiological Traits of the Marine Bacterium Alcaligenes aquatilis QD168 Isolated From Quintero Bay, Central Chile, Reveal a Robust Adaptive Response to Environmental Stressors. Front. Microbiol. 2019, 10, 528. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Lin, J.; Lin, J.; Wang, W.; Li, S. Biodegradation of Petroleum Hydrocarbons by Bacillus subtilis BL-27, a Strain with Weak Hydrophobicity. Molecules 2019, 24, 3021. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, S.; Arumugam, A.; Chandran, P. Optimization of Enterobacter cloacae (KU923381) for diesel oil degradation using response surface methodology (RSM). J. Microbiol. 2017, 55, 104–111. [Google Scholar] [CrossRef]
- Chaudhary, D.K.; Kim, D.U.; Kim, D.; Kim, J. Flavobacterium petrolei sp. nov., a novel psychrophilic, diesel-degrading bacterium isolated from oil-contaminated Arctic soil. Sci. Rep. 2019, 9, 4134. [Google Scholar] [CrossRef]
- Pacwa-Płociniczak, M.; Płaza, G.A.; Poliwoda, A.; Piotrowska-Seget, Z. Characterization of hydrocarbon-degrading and biosurfactant-producing Pseudomonas sp. P-1 strain as a potential tool for bioremediation of petroleum-contaminated soil. Environ. Sci. Pollut. Res. 2014, 21, 9385–9395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahjoubi, M.; Cappello, S.; Souissi, Y.; Jaouani, A.; Cherif, A. Microbial Bioremediation of Petroleum Hydrocarbon Contaminated Marine Environments. Recent Insights in Petroleum Science and Engineering. IntechOpen 2017, 326–350. [Google Scholar] [CrossRef] [Green Version]
- Kasai, Y.; Kishira, H.; Harayama, S. Bacteria belonging to the genus Cycloclasticus play a primary role in the degradation of aromatic hydrocarbons released in a marine environment. Appl. Environ. Microbiol. 2002, 68, 5625–5633. [Google Scholar] [CrossRef] [Green Version]
- Yakimov, M.; Giuliano, L.; Gentile, G.; Crisafi, E.; Chernikova, T.; Abraham, W.; Lünsdorf, H.; Timmis, K.; Golyshin, P. Oleispira antarctica gen. nov., sp. nov., a novel hydrocarbonoclastic marine bacterium isolated from Antarctic coastal sea water. Int. J. Syst. Evol. Microbiol. 2003, 53, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Chernikova, T.N.; Bargiela, R.; Toshchakov, S.V.; Shivaraman, V.; Lunev, E.A.; Yakimov, M.M.; Thomas, D.N.; Golyshin, P.N. Hydrocarbon-Degrading Bacteria Alcanivorax and Marinobacter Associated With Microalgae Pavlova lutheri and Nannochloropsis oculata. Front. Microbiol. 2020, 11, 572931. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Hamouda, R.A. Biodegradation of crude oil by some cyanobacteria under heterotrophic conditions. Desalin. Water Treat. 2014, 52, 1448–1454. [Google Scholar] [CrossRef]
- Kalhor, A.X.; Movafeghi, A.; Mohammadi-Nassab, A.D.; Abedi, E.; Bahrami, A. Potential of the green alga Chlorella vulgaris for biodegradation of crude oil hydrocarbons. Mar. Pollut. Bull. 2017, 123, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Nweze, N.; Aniebonam, C. Bioremediation of petroleum products impacted freshwater using locally available algae. Bio-Research 2009, 7, 484–490. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Hamouda, R.A.; Nizam, A.A. Biodegradation of crude oil by Scenedesmus obliquus and Chlorella vulgaris growing under heterotrophic conditions. Int. Biodeterior. Biodegrad. 2013, 82, 67–72. [Google Scholar] [CrossRef]
- Chavan, A.; Mukherji, S. Treatment of hydrocarbon-rich wastewater using oil degrading bacteria and phototrophic microorganisms in rotating biological contactor: Effect of N:P ratio. J. Hazard. Mater. 2008, 154, 63–72. [Google Scholar] [CrossRef]
- Semple, K.T.; Cain, R.B.; Schmidt, S. Biodegradation of aromatic compounds by microalgae. FEMS Microbiol. Lett. 1999, 170, 291–300. [Google Scholar] [CrossRef]
- Patel, J.G.; Kumar, J.I.N.; Kumar, R.N.; Khan, S.R. Enhancement of pyrene degradation efficacy of Synechocystis sp., by construction of an artificial microalgal-bacterial consortium. Cogent Chem. 2015, 1, 1064193. [Google Scholar] [CrossRef]
- Chan, S.; Luan, T.; Wong, M.; Tam, N. Removal and biodegradation of polycyclic aromatic hydrocarbons by Selenastrum capricornutum. Environ. Toxicol. Chem. 2006, 25, 1772–1779. [Google Scholar] [CrossRef] [PubMed]
- Al-Hawash, A.B.; Zhang, X.; Ma, F. Removal and biodegradation of different petroleum hydrocarbons using the filamentous fungus Aspergillus sp. RFC-1. Microbiologyopen 2019, 8, e00619. [Google Scholar] [CrossRef] [Green Version]
- Obuekwe, C.; Badrudeen, A.M.; Al-Saleh, E.; Mulder, J.L. Growth and hydrocarbon degradation by three desert fungi under conditions of simultaneous temperature and salt stress. Int. Biodeter. Biodegrad. 2005, 56, 197–205. [Google Scholar] [CrossRef]
- Wang, C.; Liu, H.; Li, J.; Sun, H. Degradation of PAHs in soil by Lasiodiplodia theobromae and enhanced benzo[a]pyrene degradation by the addition of Tween-80. Environ. Sci. Pollut. Res. 2014, 21, 10614–10625. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Li, P.; Frank, S.; Xiong, X. Biodegradation of benzo[a]pyrene in soil by Mucor sp. SF06 and Bacillus sp. SB02 co-immobilized on vermiculite. J. Environ. Sci. 2006, 18, 1204–1209. [Google Scholar] [CrossRef]
- Govarthanan, M.; Fuzisawa, S.; Hosogai, T.; Chang, Y.C. Biodegradation of aliphatic and aromatic hydrocarbons using the filamentous fungus Penicillium sp. CHY-2 and characterization of its manganese peroxidase activity. RSC Adv. 2017, 7, 20716–20723. [Google Scholar] [CrossRef] [Green Version]
- Hasan, I.F. Biodegradation of Kerosene by Aspergillus niger and Rhizopus stolinifer. J. Appl. Environ. Microbiol. 2014, 2, 31–36. [Google Scholar] [CrossRef]
- Zafra, G.; Moreno-Montaño, A.; Absalón, Á.E.; Cortés-Espinosa, D.V. Degradation of polycyclic aromatic hydrocarbons in soil by a tolerant strain of Trichoderma asperellum. Environ. Sci. Pollut. Res. 2015, 22, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Krallish, I.; Gonta, S.; Savenkova, L.; Bergauer, P.; Margesin, R. Phenol degradation by immobilized cold-adapted yeast strains of Cryptococcus terreus and Rhodotorula creatinivora. Extremophiles 2006, 10, 441–449. [Google Scholar] [CrossRef]
- Shahsavari, E.; Schwarz, A.; Aburto-Medina, A.; Ball, A.S. Biological Degradation of Polycyclic Aromatic Compounds (PAHs) in Soil: A Current Perspective. Curr. Pollut. Rep. 2019, 5, 84–92. [Google Scholar] [CrossRef]
- Yan, S.; Wu, G. Reorganization of gene network for degradation of polycyclic aromatic hydrocarbons (PAHs) in Pseudomonas aeruginosa PAO1 under several conditions. J. Appl. Genet. 2017, 58, 545–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkumari, J.; Singha, L.P.; Pandey, P. Genomic insights of aromatic hydrocarbon degrading Klebsiella pneumoniae AWD5 with plant growth promoting attributes: A paradigm of soil isolate with elements of biodegradation. 3 Biotech 2018, 8, e00619. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Sanz, D.; Redondo-Nieto, M.; Guirado, M.; Jiménez, O.P.; Millán, R.; Martin, M.; Rivilla, R. Metagenomic Insights into the Bacterial Functions of a Diesel-Degrading Consortium for the Rhizoremediation of Diesel-Polluted Soil. Genes 2019, 10, 456. [Google Scholar] [CrossRef] [Green Version]
- Loss, E.M.O.; Yu, J.H. Bioremediation and microbial metabolism of benzo(a)pyrene. Mol. Microbiol. 2018, 109, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Carmona, M.; Zamarro, M.T.; Blazquez, B.; Durante-Rodriguez, G.; Juarez, J.F.; Valderrama, J.A.; Barragan, M.J.L.; Garcia, J.L.; Diaz, E. Anaerobic Catabolism of Aromatic Compounds: A Genetic and Genomic View. Microbiol. Mol. Biol. Rev. 2009, 73, 71–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabus, R.; Boll, M.; Heider, J.; Meckenstock, R.U.; Buckel, W.; Einsle, O.; Ermler, U.; Golding, B.T.; Gunsalus, R.P.; Kroneck, P.M.H.; et al. Anaerobic microbial degradation of hydrocarbons: From enzymatic reactions to the environment. J. Mol. Microbiol. Biotechnol. 2016, 26, 5–28. [Google Scholar] [CrossRef] [Green Version]
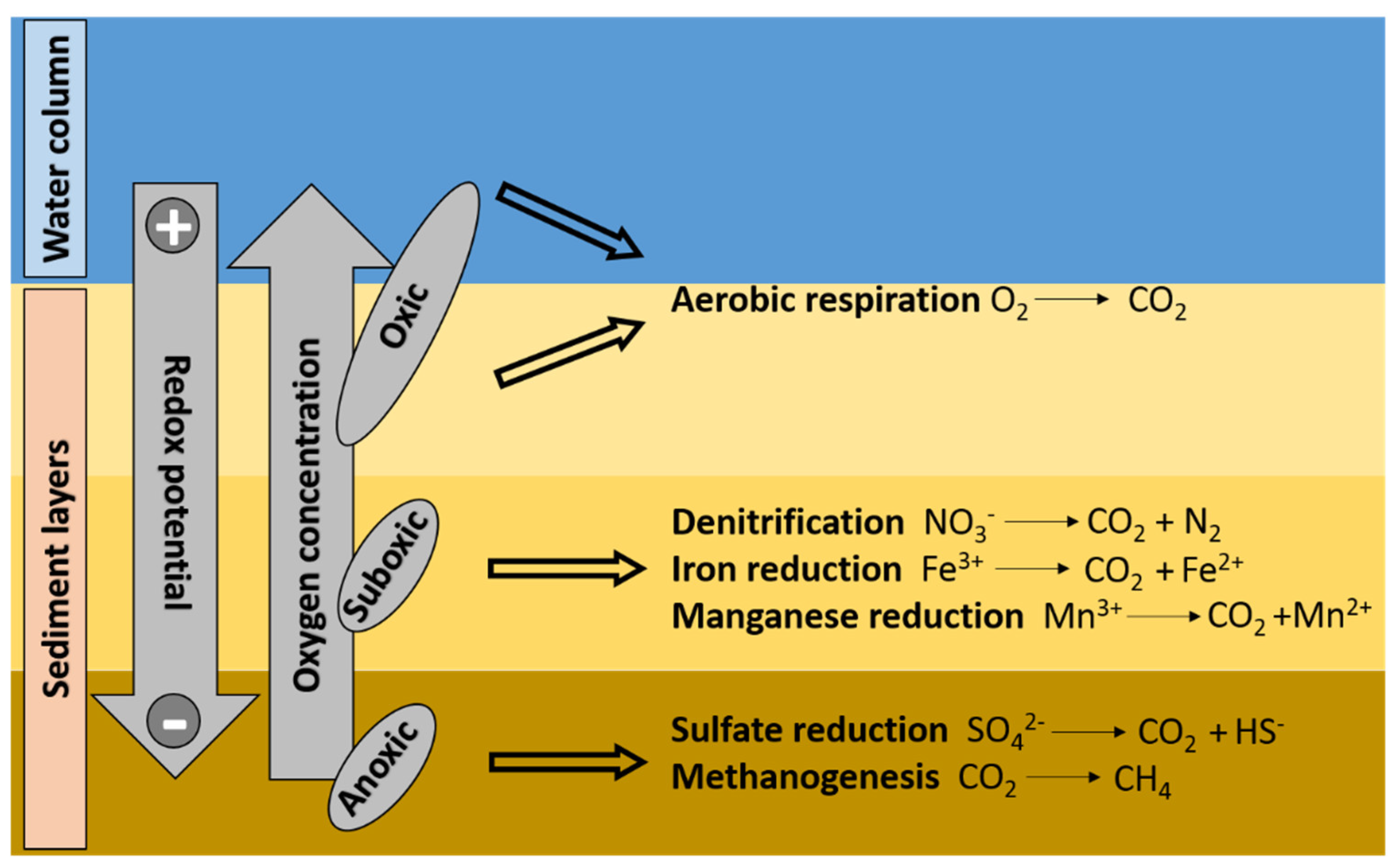
- Hastings, D.W.; Schwing, P.T.; Brooks, G.R.; Larson, R.A.; Morford, J.L.; Roeder, T.; Quinn, K.A.; Bartlett, T.; Romero, I.C.; Hollander, D.J. Changes in sediment redox conditions following the BP DWH blowout event. Deep. Res. Part II Top. Stud. Oceanogr. 2016, 129, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Vandieken, V.; Finke, N.; Thamdrup, B. Hydrogen, acetate, and lactate as electron donors for microbial manganese reduction in a manganese-rich coastal marine sediment. FEMS Microbiol. Ecol. 2014, 87, 733–745. [Google Scholar] [CrossRef] [Green Version]
- Stauffert, M.; Cravo-Laureau, C.; Duran, R. Structure of hydrocarbonoclastic nitrate-reducing bacterial communities in bioturbated coastal marine sediments. FEMS Microbiol. Ecol. 2014, 89, 580–593. [Google Scholar] [CrossRef] [Green Version]
- Dell’Anno, A.; Beolchini, F.; Gabellini, M.; Rocchetti, L.; Pusceddu, A.; Danovaro, R. Bioremediation of petroleum hydrocarbons in anoxic marine sediments: Consequences on the speciation of heavy metals. Mar. Pollut. Bull. 2009, 58, 1808–1814. [Google Scholar] [CrossRef] [PubMed]
- Hazen, T.C.; Prince, R.C.; Mahmoudi, N. Marine Oil Biodegradation. Environ. Sci. Technol. 2016, 50, 2121–2129. [Google Scholar] [CrossRef]
- Mahjoubi, M.; Aliyu, H.; Cappello, S.; Naifer, M.; Souissi, Y.; Cowan, D.A.; Cherif, A. The genome of Alcaligenes aquatilis strain BU33N: Insights into hydrocarbon degradation capacity. PLoS ONE 2019, 14, e0221574. [Google Scholar] [CrossRef]
- Sinha, R.; Krishnan, K.; Kurian, P. Complete genome sequence and comparative genome analysis of Alcanivorax sp. IO_7, a marine alkane-degrading bacterium isolated from hydrothermally-influenced deep seawater of southwest Indian ridge. Genomics 2021, 113, 884–891. [Google Scholar] [CrossRef]
- Zadjelovic, V.; Gibson, M.I.; Dorador, C.; Christie-Oleza, J.A. Genome of Alcanivorax sp. 24: A hydrocarbon degrading bacterium isolated from marine plastic debris. Mar. Genom. 2020, 49, 100686. [Google Scholar] [CrossRef]
- Tofalos, A.E.; Daghio, M.; González, M.; Papacchini, M.; Franzetti, A.; Seeger, M. Toluene degradation by Cupriavidus metallidurans CH34 in nitrate-reducing conditions and in Bioelectrochemical Systems. FEMS Microbiol. Lett. 2018, 365, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, T.; Biddle, J.F.; Teske, A.; Aitken, M.D. Cultivation-dependent and cultivation-independent characterization of hydrocarbon-degrading bacteria in Guaymas Basin sediments. Front. Microbiol. 2015, 6, 695. [Google Scholar] [CrossRef] [PubMed]
- Messina, E.; Denaro, R.; Crisafi, F.; Smedile, F.; Cappello, S.; Genovese, M.; Genovese, L.; Giuliano, L.; Russo, D.; Ferrer, M.; et al. Genome sequence of obligate marine polycyclic aromatic hydrocarbons-degrading bacterium Cycloclasticus sp. 78-ME, isolated from petroleum deposits of the sunken tanker Amoco Milford Haven, Mediterranean Sea. Mar. Genom. 2016, 25, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, L.; Shaoa, Z. Polycyclic Aromatic Hydrocarbon (PAH) Degradation Pathways of the Obligate Marine PAH Degrader Cycloclasticus sp. Strain P1. Appl. Environ. Microbiol. 2018, 84, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, T.; Whitman, W.B.; Huntemann, M.; Copeland, A.; Chen, A.; Kyrpides, N.; Markowitz, V.; Pillay, M.; Ivanova, N.; Mikhailova, N.; et al. Genome Sequence of Halomonas sp. Strain MCTG39a, a Hydrocarbon-Degrading and Exopolymeric Substance-Producing Bacterium. Genome Announc. 2015, 3, e00793-15. [Google Scholar] [CrossRef] [Green Version]
- Cheffi, M.; Hentati, D.; Chebbi, A.; Mhiri, N.; Sayadi, S.; Marqués, A.M.; Chamkha, M. Isolation and characterization of a newly naphthalene-degrading Halomonas pacifica, strain Cnaph3: Biodegradation and biosurfactant production studies. 3 Biotech 2020, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- Zenati, B.; Chebbi, A.; Badis, A.; Eddouaouda, K.; Boutoumi, H.; El Hattab, M.; Hentati, D.; Chelbi, M.; Sayadi, S.; Chamkha, M.; et al. A non-toxic microbial surfactant from Marinobacter hydrocarbonoclasticus SdK644 for crude oil solubilization enhancement. Ecotoxicol. Environ. Saf. 2018, 154, 100–107. [Google Scholar] [CrossRef]
- Gregson, B.H.; Metodieva, G.; Metodiev, M.V.; Golyshin, P.N.; McKew, B.A. Protein expression in the obligate hydrocarbon-degrading psychrophile Oleispira antarctica RB-8 during alkane degradation and cold tolerance. Environ. Microbiol. 2020, 22, 1870–1883. [Google Scholar] [CrossRef] [Green Version]
- Mangwani, N.; Kumari, S.; Das, S. Involvement of quorum sensing genes in biofilm development and degradation of polycyclic aromatic hydrocarbons by a marine bacterium Pseudomonas aeruginosa N6P6. Appl. Microbiol. Biotechnol. 2015, 99, 10283–10297. [Google Scholar] [CrossRef] [PubMed]
- Mangwani, N.; Kumari, S.; Das, S. Effect of synthetic N-acylhomoserine lactones on cell-cell interactions in marine Pseudomonas and biofilm mediated degradation of polycyclic aromatic hydrocarbons. Chem. Eng. J. 2016, 302, 172–186. [Google Scholar] [CrossRef]
- Farag, S.; Soliman, N.A.; Abdel-Fattah, Y.R. Statistical optimization of crude oil bio-degradation by a local marine bacterium isolate Pseudomonas sp. sp48. J. Genet. Eng. Biotechnol. 2018, 16, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Muriel-Millán, L.F.; Rodríguez-Mejía, J.L.; Godoy-Lozano, E.E.; Rivera-Gómez, N.; Gutierrez-Rios, R.M.; Morales-Guzmán, D.; Trejo-Hernández, M.R.; Estradas-Romero, A.; Pardo-López, L. Functional and Genomic Characterization of a Pseudomonas aeruginosa Strain Isolated From the Southwestern Gulf of Mexico Reveals an Enhanced Adaptation for Long-Chain Alkane Degradation. Front. Mar. Sci. 2019, 6, 572. [Google Scholar] [CrossRef]
- Purnomo, A.S.; Rizqi, H.D.; Harmelia, L.; Anggraeni, S.D.; Melati, R.E.; Damayanti, Z.H.; Shafwah, O.M.; Kusuma, F.C. Biodegradation of crude oil by Ralstonia pickettii under high salinity medium. Malays. J. Fundam. Appl. Sci. 2019, 15, 377–380. [Google Scholar] [CrossRef]
- Khalid, F.E.; Lim, Z.S.; Sabri, S.; Gomez-Fuentes, C.; Zulkharnain, A.; Ahmad, S.A. Bioremediation of diesel contaminated marine water by bacteria: A review and bibliometric analysis. J. Mar. Sci. Eng. 2021, 9, 155. [Google Scholar] [CrossRef]
- Somee, M.R.; Shavandi, M.; Dastgheib, S.M.M.; Amoozegar, M.A. Bioremediation of oil-based drill cuttings by a halophilic consortium isolated from oil-contaminated saline soil. 3 Biotech 2018, 8, 229. [Google Scholar] [CrossRef]
- Mcgenity, T.J.; Folwell, B.D.; Mckew, B.A.; Sanni, G.O. Marine crude-oil biodegradation: A central role for interspecies interactions. Aquat. Biosyst. 2012, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamal, M.T.; Pugazhendi, A. Degradation of petroleum hydrocarbons and treatment of refinery wastewater under saline condition by a halophilic bacterial consortium enriched from marine environment (Red Sea), Jeddah, Saudi Arabia. 3 Biotech 2018, 8, 276. [Google Scholar] [CrossRef]
- Zhao, D.; Kumar, S.; Zhou, J.; Wang, R.; Li, M.; Xiang, H. Isolation and complete genome sequence of Halorientalis hydrocarbonoclasticus sp. nov., a hydrocarbon-degrading haloarchaeon. Extremophiles 2017, 21, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Hentati, D.; Cheffi, M.; Hadrich, F.; Makhloufi, N.; Rabanal, F.; Manresa, A.; Sayadi, S.; Chamkha, M. Investigation of halotolerant marine Staphylococcus sp. CO100, as a promising hydrocarbon-degrading and biosurfactant-producing bacterium, under saline conditions. J. Environ. Manag. 2021, 277, 111480. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Fu, X.; Gao, Y.; Duan, L.; Xu, C.; Sun, W.; Li, Y.; Meng, X.; Xiao, X. Characterization of a biosurfactant-producing bacteria isolated from Marine environment: Surface activity, chemical characterization and biodegradation. J. Environ. Chem. Eng. 2020, 8, 104277. [Google Scholar] [CrossRef]
- Radmann, M.; de Morais, E.G.; de Oliveira, C.F.; Zanfonato, K.; Costa, J.A.V. Microalgae cultivation for biosurfactant production. Afr. J. Microbiol. Res. 2015, 9, 2283–2289. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Lee, S.J.; Kim, S.H.; Park, I.H.; Lee, Y.S.; Chung, S.Y.; Choi, Y.L. Characterization of new biosurfactant produced by Klebsiella sp. Y6-1 isolated from waste soybean oil. Bioresour. Technol. 2008, 99, 2288–2292. [Google Scholar] [CrossRef]
- Nikolopoulou, M.; Pasadakis, N.; Norf, H.; Kalogerakis, N. Enhanced ex situ bioremediation of crude oil contaminated beach sand by supplementation with nutrients and rhamnolipids. Mar. Pollut. Bull. 2013, 77, 37–44. [Google Scholar] [CrossRef]
- Pourfadakari, S.; Ghafari, S.; Takdastan, A.; Jorfi, S. A salt resistant biosurfactant produced by moderately halotolerant Pseudomonas aeruginosa (AHV-KH10) and its application for bioremediation of diesel-contaminated sediment in saline environment. Biodegradation 2021, 32, 327–341. [Google Scholar] [CrossRef]
- Wei, Z.; Wang, J.J.; Gaston, L.A.; Li, J.; Fultz, L.M.; DeLaune, R.D.; Dodla, S.K. Remediation of crude oil-contaminated coastal marsh soil: Integrated effect of biochar, rhamnolipid biosurfactant and nitrogen application. J. Hazard. Mater. 2020, 396, 122595. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Hou, D.; Jiang, D.; Chen, W. Bioremediation of marine oil spills by immobilized oil-degrading bacteria and nutrition emulsion. Biodegradation 2021, 32, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, Y.; Bao, M.; Li, S. Solid inoculants as a practice for bioaugmentation to enhance bioremediation of hydrocarbon contaminated areas. Chemosphere 2021, 263, 128175. [Google Scholar] [CrossRef]
- Ezekwe, C.I.; Utong, I.C. Hydrocarbon Pollution and Potential Ecological Risk of Heavy Metals in the Sediments of the Oturuba Creek, Niger Delta, Nigeria. J. Environ. Geogr. 2017, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Fonti, V.; Dell’Anno, A.; Beolchini, F. Biogeochemical Interactions in the Application of Biotechnological Strategies to Marine Sediments Contaminated with Metals. Nova Biotechnol. Chim. 2015, 14, 12–31. [Google Scholar] [CrossRef] [Green Version]
- Caporale, A.G.; Violante, A. Chemical Processes Affecting the Mobility of Heavy Metals and Metalloids in Soil Environments. Curr. Pollut. Rep. 2016, 2, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Hawksworth, D.L. The fascination of fungi: Exploring fungal diversity. Mycologist 1997, 11, 18–22. [Google Scholar] [CrossRef]
- Blackwell, M. The fungi: 1, 2, 3 … 5.1 million species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef]
- Amend, A.; Burgaud, G.; Cunliffe, M.; Edgcomb, V.P.; Ettinger, C.L.; Gutiérrez, M.H.; Heitman, J.; Hom, E.F.Y.; Ianiri, G.; Jones, A.C.; et al. Fungi in the marine environment: Open questions and unsolved problems. MBio 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Aranda, E. Promising approaches towards biotransformation of polycyclic aromatic hydrocarbons with Ascomycota fungi. Curr. Opin. Biotechnol. 2016, 38, 1–8. [Google Scholar] [CrossRef]
- Asemoloye, M.D.; Tosi, S.; Daccò, C.; Wang, X.; Xu, S.; Marchisio, M.A.; Gao, W.; Jonathan, S.G.; Pecoraro, L. Hydrocarbon degradation and enzyme activities of Aspergillus oryzae and Mucor irregularis isolated from nigerian crude oil-polluted sites. Microorganisms 2020, 8, 1912. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef] [Green Version]
- Horel, A.; Schiewer, S. Microbial degradation of different hydrocarbon fuels with mycoremediation of volatiles. Microorganisms 2020, 8, 163. [Google Scholar] [CrossRef] [Green Version]
- Adenipekun, C. Uses of mushrooms in bioremediation: A review. Biotechnol. Mol. Biol. Rev. 2012, 7, 62–68. [Google Scholar] [CrossRef]
- Al-Nasrawi, H. Biodegradation of Crude Oil by Fungi Isolated from Gulf of Mexico. J. Bioremediation Biodegrad. 2012, 3, 3. [Google Scholar]
- Bik, H.M.; Halanych, K.M.; Sharma, J.; Thomas, W.K. Dramatic shifts in benthic microbial eukaryote communities following the deepwater horizon oil spill. PLoS ONE 2012, 7, e38550. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, O.; Shinde, V.L.; Meena, R.M.; Damare, S.; Shenoy, B.D. The ITS-based phylogeny of fungi associated with tarballs. Mar. Pollut. Bull. 2016, 113, 277–281. [Google Scholar] [CrossRef]
- Barone, G.; Varrella, S.; Tangherlini, M.; Rastelli, E.; Dell’Anno, A.; Danovaro, R.; Corinaldesi, C. Marine fungi: Biotechnological perspectives from deep-hypersaline anoxic basins. Diversity 2019, 11, 113. [Google Scholar] [CrossRef] [Green Version]
- Barnes, N.M.; Khodse, V.B.; Lotlikar, N.P.; Meena, R.M.; Damare, S.R. Bioremediation potential of hydrocarbon-utilizing fungi from select marine niches of India. 3 Biotech 2018, 8, 21. [Google Scholar] [CrossRef]
- Maamar, A.; Lucchesi, M.E.; Debaets, S.; van Long, N.N.; Quemener, M.; Coton, E.; Bouderbala, M.; Burgaud, G.; Matallah-Boutiba, A. Highlighting the crude oil bioremediation potential of marine fungi isolated from the Port of Oran (Algeria). Diversity 2020, 12, 196. [Google Scholar] [CrossRef]
- Velez, P.; Gasca-Pineda, J.; Riquelme, M. Cultivable fungi from deep-sea oil reserves in the Gulf of Mexico: Genetic signatures in response to hydrocarbons. Mar. Environ. Res. 2020, 153, 104816. [Google Scholar] [CrossRef]
- Passarini, M.R.Z.; Rodrigues, M.V.N.; da Silva, M.; Sette, L.D. Marine-derived filamentous fungi and their potential application for polycyclic aromatic hydrocarbon bioremediation. Mar. Pollut. Bull. 2011, 62, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Vieira, G.A.L.; Magrini, M.J.; Bonugli-Santos, R.C.; Rodrigues, M.V.N.; Sette, L.D. Polycyclic aromatic hydrocarbons degradation by marine-derived basidiomycetes: Optimization of the degradation process. Braz. J. Microbiol. 2018, 49, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Asemoloye, M.D.; Jonathan, S.G.; Ahmad, R. Synergistic plant-microbes interactions in the rhizosphere: A potential headway for the remediation of hydrocarbon polluted soils. Int. J. Phytoremediation 2019, 21, 71–83. [Google Scholar] [CrossRef]
- Kadri, T.; Rouissi, T.; Brar, S.K.; Cledon, M.; Sarma, S.; Verma, M. Biodegradation of polycyclic aromatic hydrocarbons (PAHs) by fungal enzymes: A review. J. Environ. Sci. 2017, 51, 52–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, X.; Li, C.; Zhang, L.; Ning, G.; Shi, W.; Zhang, X.; Yang, Z. Ligninolytic enzyme involved in removal of high molecular weight polycyclic aromatic hydrocarbons by Fusarium strain ZH-H2. Environ. Sci. Pollut. Res. 2020, 27, 42969–42978. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Chandra, R. Ligninolytic enzymes and its mechanisms for degradation of lignocellulosic waste in environment. Heliyon 2020, 6, e03170. [Google Scholar] [CrossRef]
- Duan, X.; Chen, Y.; Yan, Y.; Feng, L.; Chen, Y.; Zhou, Q. New method for algae comprehensive utilization: Algae-derived biochar enhances algae anaerobic fermentation for short-chain fatty acids production. Bioresour. Technol. 2019, 289, 121637. [Google Scholar] [CrossRef] [PubMed]
- Cerniglia, C.E.; Van Baalen, C.; Gibson, D.T. Metabolism of naphthalene by the cyanobacterium Oscillatoria sp., strain JCM. J. Gen. Microbiol. 1980, 116, 485–494. [Google Scholar] [CrossRef] [Green Version]
- Lei, A.P.; Wong, Y.S.; Tam, N.F.Y. Removal of pyrene by different microalgal species. Water Sci. Technol. 2002, 46, 195–201. [Google Scholar] [CrossRef]
- Hong, Y.-W.; Yuan, D.-X.; Lin, Q.-M.; Yang, T.-L. Accumulation and biodegradation of phenanthrene and fluoranthene by the algae enriched from a mangrove aquatic ecosystem. Mar. Pollut. Bull. 2008, 56, 1400–1405. [Google Scholar] [CrossRef] [PubMed]
- Das, B.; Deka, S. A cost-effective and environmentally sustainable process for phycoremediation of oil field formation water for its safe disposal and reuse. Sci. Rep. 2019, 9, 15232. [Google Scholar] [CrossRef] [PubMed]
- Al-Hussieny, A.A.; Imran, S.G.; Jabur, Z.A. The use of local blue-green algae in the bioremediation of hydrocarbon pollutants in wastewater from oil refineries. Plant Arch. 2020, 20, 797–802. [Google Scholar]
- Tripathi, B.N.; Kumar, D. Prospects and Challenges in Algal Biotechnology; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 9789811019500. [Google Scholar]
- Ghasemi, Y.; Rasoul-Amini, S.; Fotooh-Abadi, E. The biotransformation, biodegradation, and bioremediation of organic compounds by microalgae. J. Phycol. 2011, 47, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.R.; Paniagua-Michel, J. Bioremediation of Hexadecane and Diesel Oil is Enhanced by Photosynthetically Produced Marine Biosurfactants. J. Bioremediation Biodegrad. 2013, 01, 1–5. [Google Scholar] [CrossRef]
- Ammar, S.H.; Khadim, H.J.; Mohamed, A.I. Cultivation of Nannochloropsis oculata and Isochrysis galbana microalgae in produced water for bioremediation and biomass production. Environ. Technol. Innov. 2018, 10, 132–142. [Google Scholar] [CrossRef]
- Gutierrez, T.; Green, D.H.; Nichols, P.D.; Whitman, W.B.; Semple, K.T.; Aitken, M.D. Polycyclovorans algicola gen. nov., sp. nov., an aromatic-hydrocarbon-degrading marine bacterium found associated with laboratory cultures of marine phytoplankton. Appl. Environ. Microbiol. 2013, 79, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abed, R.; Köster, J. The direct role of aerobic heterotrophic bacteria associated with cyanobacteria in the degradation of oil compounds. Int. Biodeterior. Biodegrad. 2005, 55, 29–37. [Google Scholar] [CrossRef]
- Gutierrez, T.; Nichols, P.D.; Whitman, W.B.; Aitken, M.D. Porticoccus hydrocarbonoclasticus sp. nov., an aromatic hydrocarbon-degrading bacterium identified in laboratory cultures of marine phytoplankton. Appl. Environ. Microbiol. 2012, 78, 628–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, T.; Rhodes, G.; Mishamandani, S.; Berry, D.; Whitman, W.B.; Nichols, P.D.; Semple, K.T.; Aitken, M.D. Polycyclic aromatic hydrocarbon degradation of phytoplankton-associated Arenibacter spp. and description of Arenibacter algicola sp. nov., an aromatic hydrocarbon-degrading bacterium. Appl. Environ. Microbiol. 2014, 80, 618–628. [Google Scholar] [CrossRef] [Green Version]
- Amin, S.A.; Green, D.H.; Hart, M.C.; Kupper, F.C.; Sunda, W.G.; Carrano, C.J. Photolysis of ion–siderophore chelates promotes bacteria–algal mutualism. Environ. Sci. 2009, 106, 17071–17076. [Google Scholar]
- Kazamia, E.; Czesnick, H.; Van Nguyen, T.T.; Croft, M.T.; Sherwood, E.; Sasso, S.; Hodson, S.J.; Warren, M.J.; Smith, A.G. Mutualistic interactions between vitamin B12-dependent algae and heterotrophic bacteria exhibit regulation. Environ. Microbiol. 2012, 14, 1466–1476. [Google Scholar] [CrossRef]
- Luo, S.; Chen, B.; Lin, L.; Wang, X.; Tam, N.F.Y.; Luan, T. Pyrene degradation accelerated by constructed consortium of bacterium and microalga: Effects of degradation products on the microalgal growth. Environ. Sci. Technol. 2014, 48, 13917–13924. [Google Scholar] [CrossRef]
- Tang, X.; He, L.Y.; Tao, X.Q.; Dang, Z.; Guo, C.L.; Lu, G.N.; Yi, X.Y. Construction of an artificial microalgal-bacterial consortium that efficiently degrades crude oil. J. Hazard. Mater. 2010, 181, 1158–1162. [Google Scholar] [CrossRef]
- Thompson, H.; Angelova, A.; Bowler, B.; Jones, M.; Gutierrez, T. Enhanced crude oil biodegradative potential of natural phytoplankton-associated hydrocarbonoclastic bacteria. Environ. Microbiol. 2018, 19, 2843–2861. [Google Scholar] [CrossRef]
- Fu, P.; Secundo, F. Algae and their bacterial consortia for soil bioremediation. Chem. Eng. Trans. 2016, 49, 427–432. [Google Scholar]
- Hung, C.M.; Huang, C.P.; Hsieh, S.L.; Tsai, M.L.; Chen, C.W.; Dong, C. Di Biochar derived from red algae for efficient remediation of 4-nonylphenol from marine sediments. Chemosphere 2020, 254, 126916. [Google Scholar] [CrossRef]
- Subashchandrabose, S.R.; Logeshwaran, P.; Venkateswarlu, K.; Naidu, R.; Megharaj, M. Pyrene degradation by Chlorella sp. MM3 in liquid medium and soil slurry: Possible role of dihydrolipoamide acetyltransferase in pyrene biodegradation. Algal Res. 2017, 23, 223–232. [Google Scholar] [CrossRef]
- Czaplicki, L.M.; Gunsch, C.K. Reflection on Molecular Approaches Influencing State-of-the-Art Bioremediation Design: Culturing to Microbial Community Fingerprinting to Omics. J. Environ. Eng. 2016, 142, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Yang, H.L.; Tu, Z.C.; Wang, X.L. High-throughput sequencing of microbial community diversity and dynamics during douchi fermentation. PLoS ONE 2016, 11, e0168166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malla, M.A.; Dubey, A.; Yadav, S.; Kumar, A.; Hashem, A.; Abd-Allah, E.F. Understanding and designing the strategies for the microbe-mediated remediation of environmental contaminants using omics approaches. Front. Microbiol. 2018, 9, 1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, M.; Singh, D.; Vikram, S.; Singh, V.; Kumar, S. Metagenomic Approach towards Bioprospection of Novel Biomolecule(s) and Environmental Bioremediation. Annu. Res. Rev. Biol. 2018, 22, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ngara, T.R.; Zhang, H. Recent Advances in Function-based Metagenomic Screening. Genom. Proteom. Bioinform. 2018, 16, 405–415. [Google Scholar] [CrossRef]
- Popovic, A.; Tchigvintsev, A.; Tran, H.; Chernikova, T.N.; Golyshina, O.V.; Yakimov, M.M.; Golyshin, P.N.; Yakunin, A.F. Metagenomics as a tool for enzyme discovery: Hydrolytic enzymes from marine-related metagenomes. Adv. Exp. Med. Biol. 2015, 883, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Strazzulli, A.; Fusco, S.; Cobucci-Ponzano, B.; Moracci, M.; Contursi, P. Metagenomics of microbial and viral life in terrestrial geothermal environments. Rev. Environ. Sci. Biotechnol. 2017, 16, 425–454. [Google Scholar] [CrossRef] [Green Version]
- Coughlan, L.M.; Cotter, P.D.; Hill, C.; Alvarez-Ordóñez, A. Biotechnological applications of functional metagenomics in the food and pharmaceutical industries. Front. Microbiol. 2015, 6, 672. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.H.; Ye, C.C.; Zhou, Q.Z.; Wu, X.Y.; Yuan, J.P.; Peng, J.; Deng, H.; Wang, J.H. Genome sequencing reveals the potential of Achromobacter sp. HZ01 for bioremediation. Front. Microbiol. 2017, 8, 1507. [Google Scholar] [CrossRef] [Green Version]
- Zakaria, N.N.; Convey, P.; Gomez-Fuentes, C.; Zulkharnain, A.; Sabri, S.; Shaharuddin, N.A.; Ahmad, S.A. Oil bioremediation in the marine environment of antarctica: A review and bibliometric keyword cluster analysis. Microorganisms 2021, 9, 419. [Google Scholar] [CrossRef]
- Vázquez, S.; Monien, P.; Minetti, R.P.; Jürgens, J.; Curtosi, A.; Primitz, J.V.; Frickenhaus, S.; Abele, D.; Mac Cormack, W.; Helmke, E. Bacterial communities and chemical parameters in soils and coastal sediments in response to diesel spills at Carlini Station, Antarctica. Sci. Total Environ. 2017, 605–606, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Shukla, P. Designing synthetic microbial communities for effectual bioremediation: A review. Biocatal. Biotransformation 2020, 38, 405–414. [Google Scholar] [CrossRef]
- Jaiswal, S.; Shukla, P. Alternative Strategies for Microbial Remediation of Pollutants via Synthetic Biology. Front. Microbiol. 2020, 11, 808. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, S.; Méndez, V.; Aguila, P.; Seeger, M. Bioremediation of petroleum hydrocarbons: Catabolic genes, microbial communities, and applications. Appl. Microbiol. Biotechnol. 2014, 98, 4781–4794. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Erdner, D.L.; Rosenheim, B.E.; Shetty, P.; Seitz, K.W.; Baker, B.J.; Liu, Z. Hydrocarbon degradation and response of seafloor sediment bacterial community in the northern Gulf of Mexico to light Louisiana sweet crude oil. ISME J. 2018, 12, 2532–2543. [Google Scholar] [CrossRef]
- Bashiardes, S.; Zilberman-Schapira, G.; Elinav, E. Use of metatranscriptomics in microbiome research. Bioinform. Biol. Insights 2016, 10, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, D.P.; Kukhtin, A.; Mokhiber, R.; Knickerbocker, C.; Ogles, D.; Rudy, G.; Golova, J.; Long, P.; Peacock, A. Monitoring microbial community structure and dynamics during in situ u(vi) bioremediation with a field-portable microarray analysis system. Environ. Sci. Technol. 2010, 44, 5516–5522. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.C.; Ipcho, S.V.S.; Trengove, R.D.; Oliver, R.P.; Solomon, P.S. Assessing the impact of transcriptomics, proteomics and metabolomics on fungal phytopathology. Mol. Plant Pathol. 2009, 10, 703–715. [Google Scholar] [CrossRef]
- Arsène-Ploetze, F.; Bertin, P.N.; Carapito, C. Proteomic tools to decipher microbial community structure and functioning. Environ. Sci. Pollut. Res. 2015, 22, 13599–13612. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.; Mann, M. Is Proteomics the New Genomics? Cell 2007, 130, 395–398. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Cho, K.; Yun, S.H.; Kim, J.Y.; Kwon, K.H.; Yoo, J.S.; Kim, S. Il Analysis of aromatic catabolic pathways in Pseudomonas putida KT 2440 using a combined proteomic approach: 2-DE/MS and cleavable isotope-coded affinity tag analysis. Proteomics 2006, 6, 1301–1318. [Google Scholar] [CrossRef] [PubMed]
- Swati; Kumari, M.; Ghosh, P.; Thakur, I.S. Evaluation of a biosurfactant producing bacterial strain Pseudomonas sp. ISTPY2 for efficient pyrene degradation and landfill soil bioremediation through soil microcosm and proteomic studies. Bioresour. Technol. Reports 2020, 12, 100607. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by Gas Chromatography-Mass Spectrometry: The combination of targeted and untargeted profiling. Curr Protoc Mol. Biol 2016, 114, 1–32. [Google Scholar] [CrossRef]
- Singh, O.V. Proteomics and metabolomics: The molecular make-up of toxic aromatic pollutant bioremediation. Proteomics 2006, 6, 5481–5492. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.; Dubey, D.; Sharma, M.; Patel, A.; Guleria, A.; Pruthi, P.A.; Kumar, D.; Pruthi, V.; Poluri, K.M. NMR-Based Metabolomic Approach to Elucidate the Differential Cellular Responses during Mitigation of Arsenic(III, V) in a Green Microalga. ACS Omega 2018, 3, 11847–11856. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Organisms | Genus | Ref. |
|---|---|---|
| Bacteria | Alcaligens | [63] |
| Bacillus | [64] | |
| Enterobacter | [65] | |
| Flavobacterium | [66] | |
| Pseuodmonas | [67] | |
| Alcanivorax | [24] | |
| Thallassolituus | [68] | |
| Cycloclasticus | [69] | |
| Oleispira | [70] | |
| Marinobacter | [71] | |
| Microalgae/Cyanobacteria | Spirulina | [72] |
| Chlorella | [73] | |
| Spirogyra | [74] | |
| Scenedesmus | [75] | |
| Oscillatoria | [76] | |
| Chlorococcum | [77] | |
| Synechocystis | [78] | |
| Nannochloropsis | [71] | |
| Selenastrum | [79] | |
| Fungi | Aspergillus | [80] |
| Curvularia | [28] | |
| Drechslera | [81] | |
| Fusarium | [81] | |
| Lasiodiplodia | [82] | |
| Mucor | [83] | |
| Penicillium | [84] | |
| Rhizopus | [85] | |
| Trichoderma | [86] | |
| Cryptococcus | [87] |
| Oil-Degrading Bacteria. | Hydrocarbon Specificity | Reference |
|---|---|---|
| Alcaligenes aquatilis BU33N | Crude oil and phenanthrene | [100] |
| Alcanivorax sp. IO_7 | Alkane | [101] |
| Alcanivorax sp. 24 | Alkanes | [102] |
| Cupriavidus metallidurans CH34 | Toluene | [103] |
| Cycloclasticus sp. strain BG-2 | Phenanthrene | [104] |
| Cycloclasticus sp. 78-ME | Polycyclic aromatic hydrocarbons | [105] |
| Cycloclasticus sp. strain P1 | Naphthalene, phenanthrene, pyrene | [106] |
| Halomonas sp. strain MCTG39a | Hexadecane | [107] |
| Halomonas pacifica strain Cnaph3 | Naphthalene | [108] |
| Marinobacter hydrocarbonoclasticus SdK644 | Crude oil | [109] |
| Oleispira antarctica RB-8 | Aliphatic alkanes | [110] |
| Pseudomonas aeruginosa N6P6 | Phenanthrene and pyrene | [111] |
| Pseudomonas pseudoalcaligenes NP103 | Phenanthrene and pyrene | [112] |
| Pseudomonas sp. sp48 | Phenol, naphtalene, pentadecane | [113] |
| Pseudomonas aeruginosa GOM1 | Hexadecane | [114] |
| Ralstonia pickettii | Crude oil | [115] |
| Species | Location | Compounds | Ref. |
|---|---|---|---|
| Aspergillus sydowii NIOSN-SK56C42 | Deep sea sediment (Arabian sea) | Crude oil, alkanes | [145] |
| Acremonium sclerotigenum NIOSN-M109 | Mangrove sediment (Panaji, Goa) | ||
| Penicillium citrinum NIOSN-M126 | Mangrove sediment (Panaji, Goa) | ||
| Aspergillusflavus NIOSN-SK56S22 | Deep sea (Arabian sea) | ||
| Penicillium polonicum | Marine sediments (Port of Oran, Algeria) | Crude oil | [146] |
| Penicillium cyclopium | |||
| Penicillium mononematosum | |||
| Penicillium chrysogenum | |||
| Aureobasidium sp. | Deep sea oil reserves (Gulf of Mexico) | hexadecane and 1-hexadecene | [147] |
| P. brevicompactum | |||
| Phialocephala sp. | |||
| Penicillium sp. | |||
| Cladosporium sp. 1, 2 | |||
| C. gracilis | |||
| Aspergillus sclerotiorum CBMAI 849 | Coastal Atlantic Ocean (São Paulo, Brazil) | pyrene and benzo[a]pyrene | [148] |
| Mucor racemosus CBMAI 847 | |||
| Tinctoporellus sp. CBMAI1061 | Marine sponges (Dragmacidonreticulatum and Amphimedon viridis) | PAHs, Pyrene | [149] |
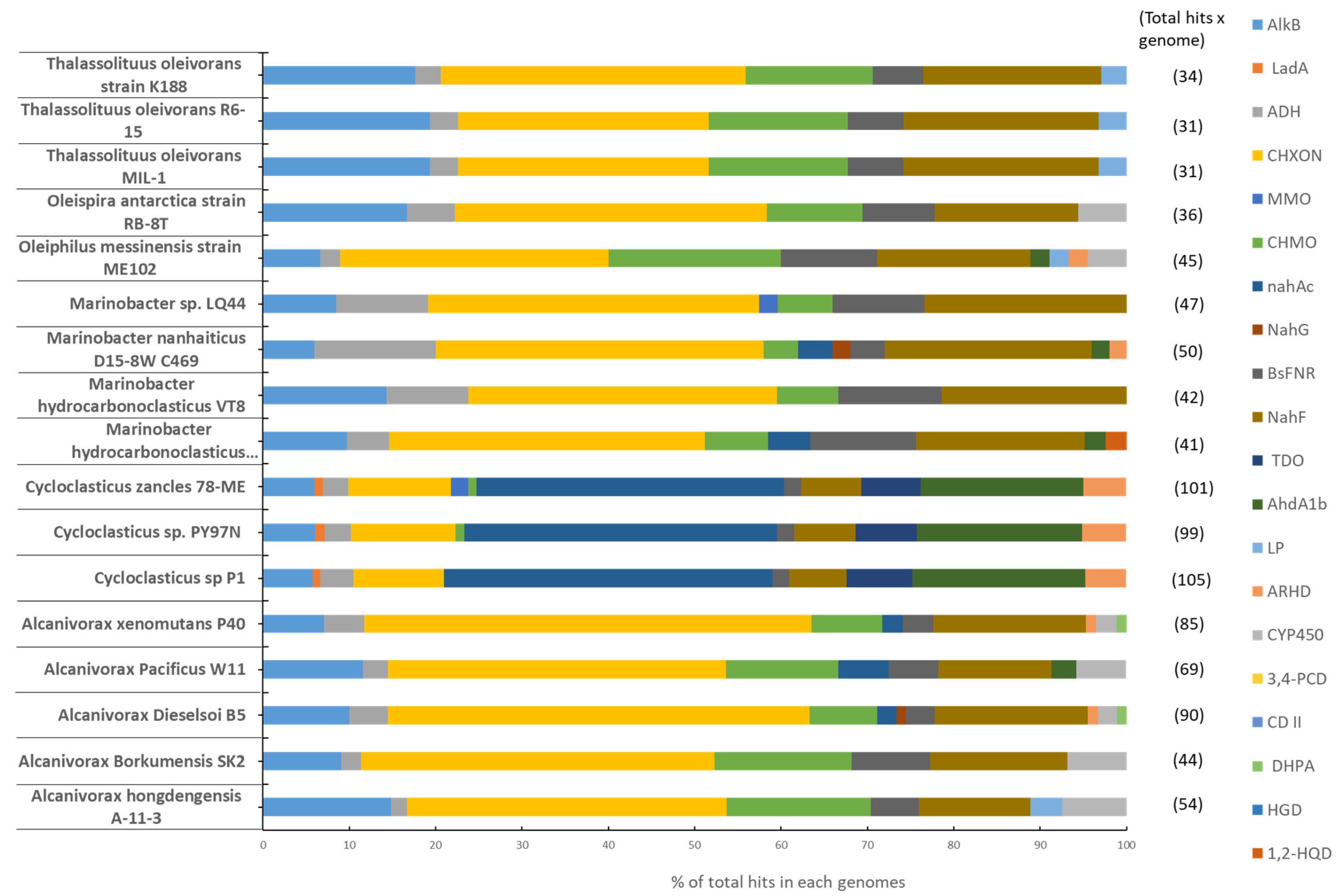
| Enzymes | EC Num. | Genomes | Access. Num. |
|---|---|---|---|
| Alkane hydroxylase | 1.14.15.3 | Alcanivorax hongdengensis A-11-3 | NZAMRJ000001 |
| Alkane monooxygenase LadA | 1.14.14.28 | Alcanivorax Borkumensis SK2 | NC008260 |
| Alcohol dehydrogenase | 1.1.1.2 | Alcanivorax Dieselsoi B5 | NC018691 |
| Cyclohexanol dehydrogenase | 1.1.1.245 | Alcanivorax Pacificus W11-5 | NZCP004387 |
| Methane monooxygenase | 1.14.13.25 | Alcanivorax xenomutans P40 | NZCP012331 |
| Cyclohexanone monooxygenase | 1.14.13.22 | Cycloclasticus sp. P1 | NC018697 |
| Naphthalene dioxygenase | 1.14.12.12 | Cycloclasticus sp. PY97N | NZCP023664 |
| Salicylate 1-monooxygenase | 1.14.13.1 | Cycloclasticus zancles 78-ME | NC021917 |
| Ferrodoxin reductase | 1.18.1.2 | M. hydrocarbonoclasticus ATCC49840 | NC017067 |
| Salicylaldehyde dehydrogenase | 1.2.1.65 | Marinobacter hydrocarbonoclasticus VT8 | NC008740 |
| Toluene dioxygenase | 1.14.12.11 | Marinobacter nanhaiticus D15-8W C469 | NZAPLQ000001 |
| Ethylbenzene dioxygenaseL-sub. | 1.14.12.18 | Marinobacter sp. LQ44 | NZCP014754 |
| Lipase | 3.1.1.3 | Oleiphilus messinensis strain ME102 | NZCP021425 |
| Aromatic ring-hydroxylating diox. | 1.14.12.3 | Oleispira antarctica strain RB-8 | FO203512 |
| Cytochrome P450 | 1.14.14.1 | Thalassolituus oleivorans MIL-1 | NC020888 |
| Protocatechuate 3,4-dioxygenase | 1.13.11.3 | Thalassolituus oleivorans R6-15 | NZCP006829 |
| Catechol 1,2-dioxygenase | 1.13.11.1 | Thalassolituus oleivorans strain K188 | NZ_CP017810.1 |
| 3,4-dihydroxyphenylacetate diox. | 1.13.11.15 | ||
| Homogentisate 1,2-dioxygenase | 1.13.11.5 | ||
| Hydroxyquinol 1,2-dioxygenase | 1.13.11.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dell’ Anno, F.; Rastelli, E.; Sansone, C.; Brunet, C.; Ianora, A.; Dell’ Anno, A. Bacteria, Fungi and Microalgae for the Bioremediation of Marine Sediments Contaminated by Petroleum Hydrocarbons in the Omics Era. Microorganisms 2021, 9, 1695. https://doi.org/10.3390/microorganisms9081695
Dell’ Anno F, Rastelli E, Sansone C, Brunet C, Ianora A, Dell’ Anno A. Bacteria, Fungi and Microalgae for the Bioremediation of Marine Sediments Contaminated by Petroleum Hydrocarbons in the Omics Era. Microorganisms. 2021; 9(8):1695. https://doi.org/10.3390/microorganisms9081695
Chicago/Turabian StyleDell’ Anno, Filippo, Eugenio Rastelli, Clementina Sansone, Christophe Brunet, Adrianna Ianora, and Antonio Dell’ Anno. 2021. "Bacteria, Fungi and Microalgae for the Bioremediation of Marine Sediments Contaminated by Petroleum Hydrocarbons in the Omics Era" Microorganisms 9, no. 8: 1695. https://doi.org/10.3390/microorganisms9081695