
Confounding Factors Influencing the Kinetics and Magnitude of Serological Response Following Administration of BNT162b2

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Analytical Procedures
2.3. Statistical Analysis
3. Results
3.1. Global Kinetics
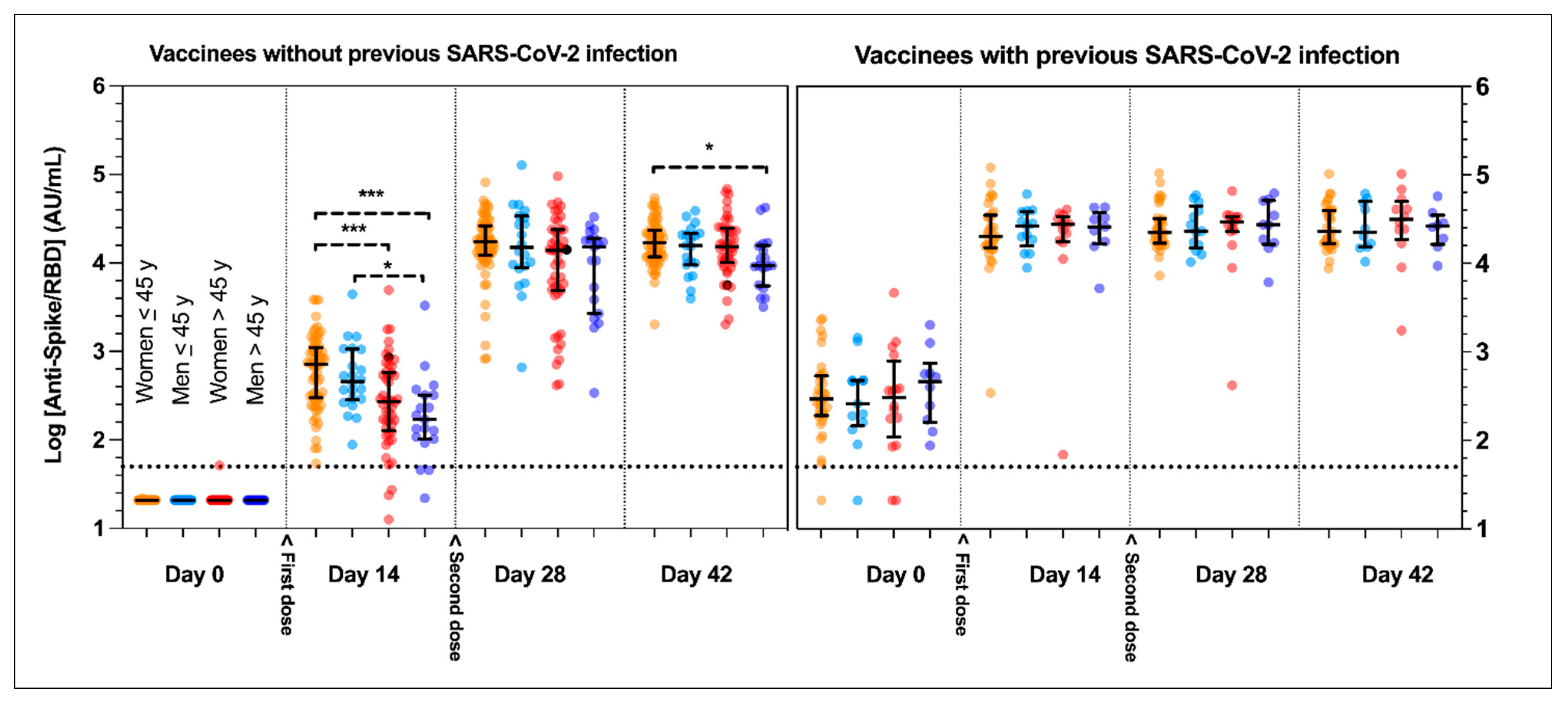
3.2. Influence of Previous Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection
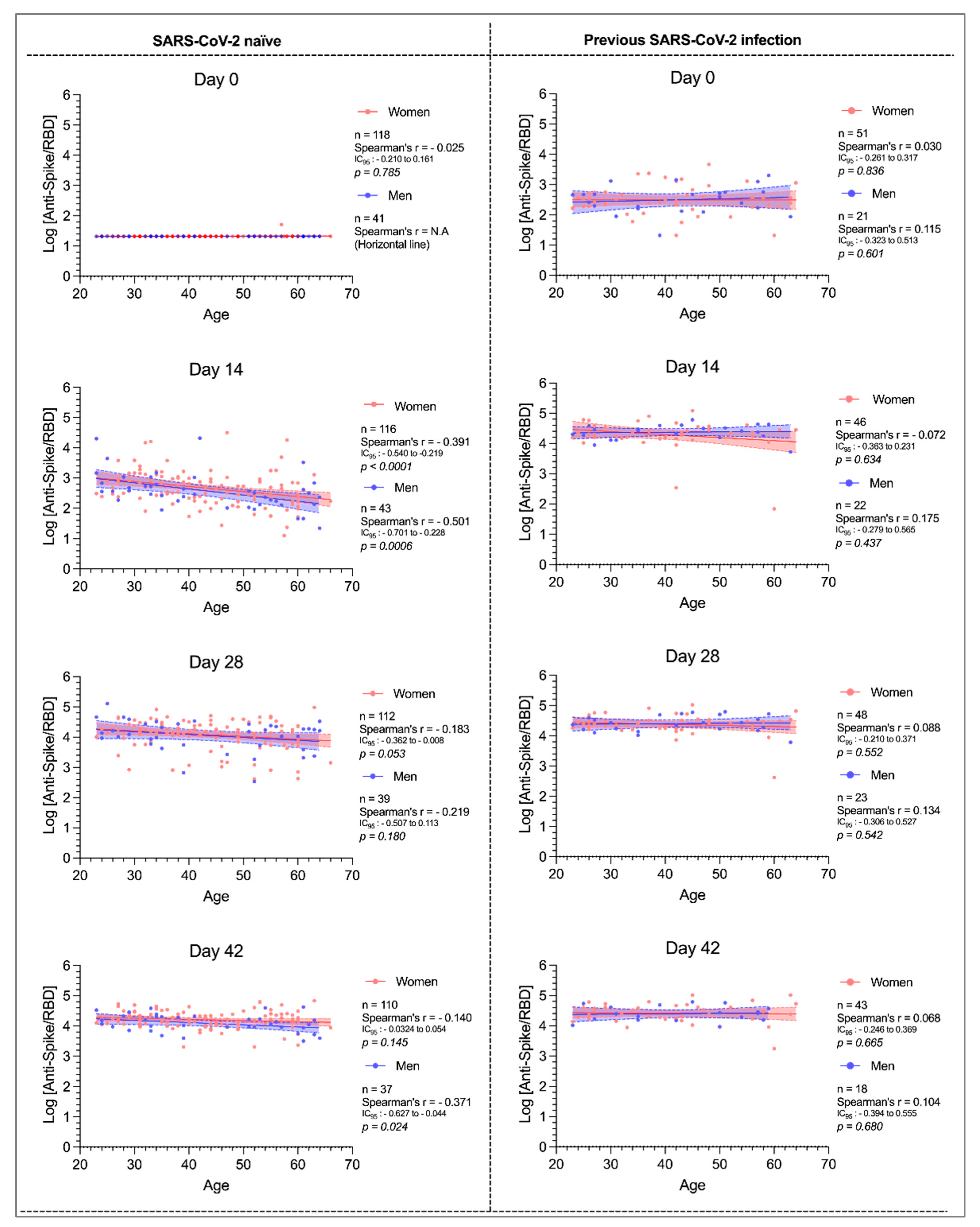
3.3. Influence of Age and Sex
3.4. Influence of ABO Blood Group
3.5. Other Potential Confounding Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, Y.; Pang, Y.; Lyu, Z.; Wang, R.; Wu, X.; You, C.; Zhao, H.; Manickam, S.; Lester, E.; Wu, T.; et al. The COVID-19 Vaccines: Recent Development, Challenges and Prospects. Vaccines 2021, 9, 349. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Favresse, J.; Bayart, J.-L.; Mullier, F.; Dogné, J.-M.; Closset, M.; Douxfils, J. Early antibody response in health-care professionals after two doses of SARS-CoV-2 mRNA vaccine (BNT162b2). Clin. Microbiol. Infect. 2021, in press. [Google Scholar] [CrossRef]
- Favresse, J.; Eucher, C.; Elsen, M.; Tré-Hardy, M.; Dogné, J.-M.; Douxfils, J. Clinical performance of the Elecsys electrochemiluminescent immunoassay for the detection of SARS-CoV-2 total antibodies. Clin. Chem. 2020, 66, 1104–1106. [Google Scholar] [CrossRef]
- Press Release: Une Seule Dose de Vaccin pour les Personnes Ayant déjà Eté Infectées par le SARS-CoV-2. Available online: https://www.has-sante.fr/jcms/p_3237456/en/une-seule-dose-de-vaccin-pour-les-personnes-ayant-deja-ete-infectees-par-le-sars-cov-2 (accessed on 9 March 2021).
- Manisty, C.; Otter, A.D.; Treibel, T.A.; McKnight, Á.; Altmann, D.M.; Brooks, T.; Noursadeghi, M.; Boyton, R.J.; Semper, A.; Moon, J.C. Antibody response to first BNT162b2 dose in previously SARS-CoV-2-infected individuals. Lancet 2021, 397, 1057–1058. [Google Scholar] [CrossRef]
- Strangfeld, A.; Schäfer, M.; Gianfrancesco, M.A.; Lawson-Tovey, S.; Liew, J.W.; Ljung, L.; Mateus, E.F.; Richez, C.; Santos, M.J.; Schmajuk, G.; et al. Factors associated with COVID-19-related death in people with rheumatic diseases: Results from the COVID-19 Global Rheumatology Alliance physician-reported registry. Ann. Rheum. Dis. 2021, in press. [Google Scholar] [CrossRef]
- Williamson, E.J.; Walker, A.J.; Bhaskaran, K.; Bacon, S.; Bates, C.; Morton, C.E.; Curtis, H.J.; Mehrkar, A.; Evans, D.; Inglesby, P.; et al. Factors associated with COVID-19-related death using OpenSAFELY. Nature 2020, 584, 430–436. [Google Scholar] [CrossRef]
- Poco, P.C.E.; Aliberti, M.J.R.; Dias, M.B.; Takahashi, S.D.F.; Leonel, F.C.; Altona, M.; Venys, A.L.; Shin-Ike, I.A.; Garcia, B.A.; Sumita, L.H.; et al. Divergent: Age, Frailty, and Atypical Presentations of COVID-19 in Hospitalized Patients. J. Gerontol. Ser. A 2021, 76, e46–e51. [Google Scholar] [CrossRef] [PubMed]
- Seidu, S.; Gillies, C.; Zaccardi, F.; Kunutsor, S.K.; Hartmann-Boyce, J.; Yates, T.; Singh, A.K.; Davies, M.J.; Khunti, K. The impact of obesity on severe disease and mortality in people with SARS-CoV-2: A systematic review and meta-analysis. Endocrinol. Diabetes Metab. 2021, 4, e00176. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Marriott, I.; Fish, E.N. Sex-based differences in immune function and responses to vaccination. Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giefing-Kröll, C.; Berger, P.; Lepperdinger, G.; Grubeck-Loebenstein, B. How sex and age affect immune responses, susceptibility to infections, and response to vaccination. Aging Cell 2015, 14, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Curtis, N. Factors That Influence the Immune Response to Vaccination. Clin. Microbiol. Rev. 2019, 32, e00084-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butterworth, M.; McClellan, B.H.; Aklansmith, M. Influence of sex on immunoglobulin levels. Nature 1967, 214, 1224–1225. [Google Scholar] [CrossRef] [PubMed]
- Grzelak, L.; Velay, A.; Madec, Y.; Gallais, F.; Staropoli, I.; Schmidt-Mutter, C.; Wendling, M.-J.; Meyer, N.; Planchais, C.; Rey, D. Sex differences in the evolution of neutralizing antibodies to SARS-CoV-2. J. Infect. Dis. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Korte, W.; Buljan, M.; Rösslein, M.; Wick, P.; Golubov, V.; Jentsch, J.; Reut, M.; Peier, K.; Nohynek, B.; Fischer, A. SARS-CoV-2 IgG and IgA antibody response is gender dependent; and IgG antibodies rapidly decline early on. J. Infect. 2021, 82, e11–e14. [Google Scholar] [CrossRef]
- Mehew, J.; Johnson, R.; Roberts, D.; Harvala, H. Convalescent plasma for COVID-19: Male gender, older age and hospitalisation associated with high neutralising antibody levels, England, 22 April to 12 May 2020. Eurosurveillance 2020, 25, 2001754. [Google Scholar] [CrossRef]
- Kontopoulou, K.; Ainatzoglou, A.; Nakas, C.; Papazisis, G. Immunogenicity after the First Dose of the BNT162b2 mRNA Covid-19 Vaccine: Real-World Evidence from Greek Healthcare Workers. 2021. Available online: http://dx.doi.org/10.2139/ssrn.3786138 (accessed on 18 June 2021).
- Jabal, K.A.; Ben-Amram, H.; Beiruti, K.; Batheesh, Y.; Sussan, C.; Zarka, S.; Edelstein, M. Impact of age, gender, ethnicity and prior disease status on immunogenicity following administration of a single dose of the BNT162b2 mRNA Covid-19 Vaccine: Real-world evidence from Israeli healthcare workers, December–January 2020. Eurosurveillance 2021, 26, 1–5. [Google Scholar] [CrossRef]
- Terpos, E.; Trougakos, I.P.; Apostolakou, F.; Charitaki, I.; Sklirou, A.D.; Mavrianou, N.; Papanagnou, E.-D.; Liacos, C.-I.; Gumeni, S.; Rentziou, G. Age-and gender-dependent antibody responses against SARS-CoV-2 in health workers and octogenarians after vaccination with the BNT162b2 mRNA vaccine. Am. J. Hematol. 2021, 96, E257–E259. [Google Scholar] [CrossRef]
- Höpping, A.M.; McElhaney, J.; Fonville, J.M.; Powers, D.C.; Beyer, W.E.; Smith, D.J. The confounded effects of age and exposure history in response to influenza vaccination. Vaccine 2016, 34, 540–546. [Google Scholar] [CrossRef]
- Castle, S.C. Clinical relevance of age-related immune dysfunction. Clin. Infect. Dis. 2000, 31, 578–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.H.; Kozlovsky, B.F.; Effros, R.B.; Grubeck-Loebenstein, B.; Edelman, R.; Sztein, M.B. Vaccination in the elderly: An immunological perspective. Trends Immunol. 2009, 30, 351–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prendecki, M.; Clarke, C.; Brown, J.; Cox, A.; Gleeson, S.; Guckian, M.; Randell, P.; Dalla Pria, A.; Lightstone, L.; Xu, X.-N. Effect of previous SARS-CoV-2 infection on humoral and T-cell responses to single-dose BNT162b2 vaccine. Lancet 2021, 397, 1178–1181. [Google Scholar] [CrossRef]
- Jalkanen, P.; Kolehmainen, P.; Häkkinen, H.; Huttunen, M.; Tähtinen, P.; Lundberg, R.; Maljanen, S.; Reinholm, A.; Tauriainen, S.; Pakkanen, S. COVID-19 mRNA Vaccine Induced Antibody Responses and Neutralizing Antibodies against Three SARS-CoV-2 Variants. 2021. Available online: https://assets.researchsquare.com/files/rs-343388/v1_stamped.pdf (accessed on 18 June 2021).
- Zhao, J.; Yang, Y.; Huang, H.; Li, D.; Gu, D.; Lu, X.; Zhang, Z.; Liu, L.; Liu, T.; Liu, Y. Relationship between the ABO Blood Group and the COVID-19 Susceptibility. Clin. Infect. Dis. 2020, 20, 1–4. [Google Scholar]
- Wu, Y.; Feng, Z.; Li, P.; Yu, Q. Relationship between ABO blood group distribution and clinical characteristics in patients with COVID-19. Clin. Chim. Acta 2020, 509, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Belančić, A.; Kresović, A.; Rački, V. Potential pathophysiological mechanisms leading to increased COVID-19 susceptibility and severity in obesity. Obes. Med. 2020, 19, 100259. [Google Scholar] [CrossRef] [PubMed]
- Dietz, W.; Santos-Burgoa, C. Obesity and its implications for COVID-19 mortality. Obesity 2020, 28, 1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, K.M.; Gray, C.M.; Bekker, L.-G. Is obesity a risk factor for vaccine non-responsiveness? PLoS ONE 2013, 8, e82779. [Google Scholar] [CrossRef] [Green Version]
- Bloch, E.M.; Patel, E.U.; Marshall, C.; Littlefield, K.; Goel, R.; Grossman, B.J.; Winters, J.L.; Shrestha, R.; Burgess, I.; Laeyendecker, O. ABO blood group and SARS-CoV-2 antibody response in a convalescent donor population. Vox Sang. 2021, in press. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variables | Total Subject Number (n = 231) |
|---|---|
| Age (median, range) | 43 (23–66) |
| ≤45 years of age (n, %) | 139 (40%) |
| Previous SARS-CoV-2 infection (n,%) | 47 (34%) |
| >45 years of age (n, %) | 92 (60%) |
| Previous SARS-CoV-2 infection (n,%) | 25 (27%) |
| Sex (n, %) | |
| Female | 169 (73%) |
| Previous SARS-CoV-2 infection (n,%) | 51 (30%) |
| Male | 62 (27%) |
| Previous SARS-CoV-2 infection (n,%) | 21(34%) |
| Previous SARS-CoV-2 infection (n, %) | 72 (31%) |
| Female (n, %) | 51 (71%) |
| Male (n,%) | 21 (29%) |
| ≤45 years of age (n, %) | 47 (65%) |
| >45 years of age (n, %) | 25 (35%) |
| BMI in kg/m2 (median, range) † | 23.7 (15.3–48.2) |
| ABO blood group (n, %) | |
| A | 76 (33%) |
| B | 9 (4%) |
| AB | 18 (8%) |
| O | 98 (42%) |
| Unknown | 30 (13%) |
| For female gender only | |
| Childbearing age (n, %) | 121 (72%) |
| Hormonal contraception | 78 (65%) |
| No hormonal contraception | 38 (31%) |
| Unknown | 5 (4%) |
| Menopausal (n, %) | 48 (28%) |
| Hormonal replacement therapy | 13 (27%) |
| No hormonal replacement therapy | 30 (63%) |
| Unknown | 5 (10%) |
| AU/mL | Women ≤45 Years | Men ≤45 Years | Women >45 Years | Men >45 Years | p-Value |
| Baseline | 21 † (21–21) | 21 † (21–21) | 21 † (21–21) | 21 † (21–21) | 0.839 |
| Day 14 | 718 (299–1111) | 459 (284–1062) | 271 (128–583) | 170 (103–323) | <0.0001 |
| Day 28 | 17343 (12,203–26,312) | 14962 (8803–33,829) | 13841 (4913–23,750) | 15244 (2674–19,048) | 0.0755 |
| Day 42 | 16843 (11,768–23,427) | 15784 (9530–21,602) | 15372 (10,071–24,685) | 9327 (5520–15,784) | 0.0269 |
| AU/mL | Women ≤45 Years | Men ≤45 Years | Women >45 Years | Men >45 Years | p-Value |
| Baseline | 293 (190–534) | 260 (146–477) | 304 (110–788) | 462 (160–742) | 0.849 |
| Day 14 | 20,055 (15,047–35,055) | 26,535 (15,722–38,628) | 27,721 (17,683–33,703) | 25,826 (16,659–37,683) | 0.927 |
| Day 28 | 22,270 (16,925–32,220) | 23,236 (14,921–44,441) | 29,584 (22,875–33,881) | 27,402 (16,448–51,377) | 0.905 |
| Day 42 | 23,057 (16,857–39,774) | 22,532 (15,244–50,159) | 31,359 (18,646–50,229) | 26,415 (16,388–34,968) | 0.876 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bayart, J.-L.; Morimont, L.; Closset, M.; Wieërs, G.; Roy, T.; Gerin, V.; Elsen, M.; Eucher, C.; Van Eeckhoudt, S.; Ausselet, N.; et al. Confounding Factors Influencing the Kinetics and Magnitude of Serological Response Following Administration of BNT162b2. Microorganisms 2021, 9, 1340. https://doi.org/10.3390/microorganisms9061340
Bayart J-L, Morimont L, Closset M, Wieërs G, Roy T, Gerin V, Elsen M, Eucher C, Van Eeckhoudt S, Ausselet N, et al. Confounding Factors Influencing the Kinetics and Magnitude of Serological Response Following Administration of BNT162b2. Microorganisms. 2021; 9(6):1340. https://doi.org/10.3390/microorganisms9061340
Chicago/Turabian StyleBayart, Jean-Louis, Laure Morimont, Mélanie Closset, Grégoire Wieërs, Tatiana Roy, Vincent Gerin, Marc Elsen, Christine Eucher, Sandrine Van Eeckhoudt, Nathalie Ausselet, and et al. 2021. "Confounding Factors Influencing the Kinetics and Magnitude of Serological Response Following Administration of BNT162b2" Microorganisms 9, no. 6: 1340. https://doi.org/10.3390/microorganisms9061340