
Detection of Mycoplasma agalactiae in Ticks (Rhipicephalus bursa) Collected by Sheep and Goats in Sicily (South-Italy), Endemic Area for Contagious Agalactia

 , , , ,
, , , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
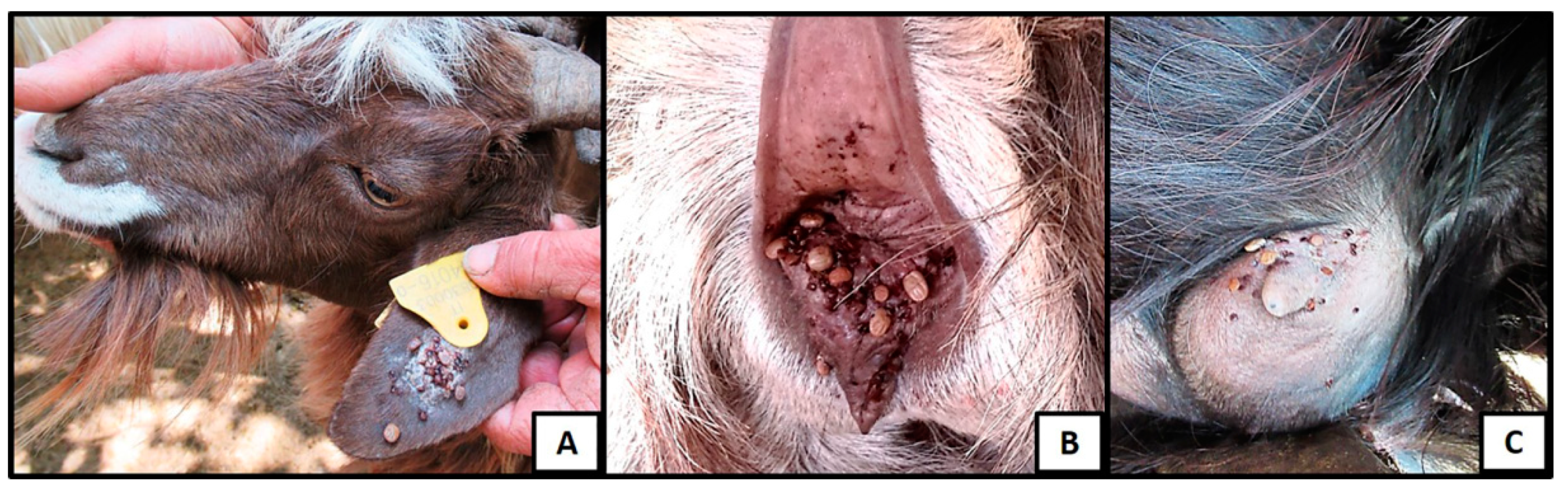
2.2. Study Design and Sampling
2.3. Tick Identification
2.4. Microbiological Analysis
2.5. Serological Analysis
2.6. DNA Extraction and PCR Amplification
2.7. DNA sequencing
3. Results
3.1. Ticks Identification
3.2. Microbiological Analysis
3.3. Molecular Analysis
3.4. Serological Analysis
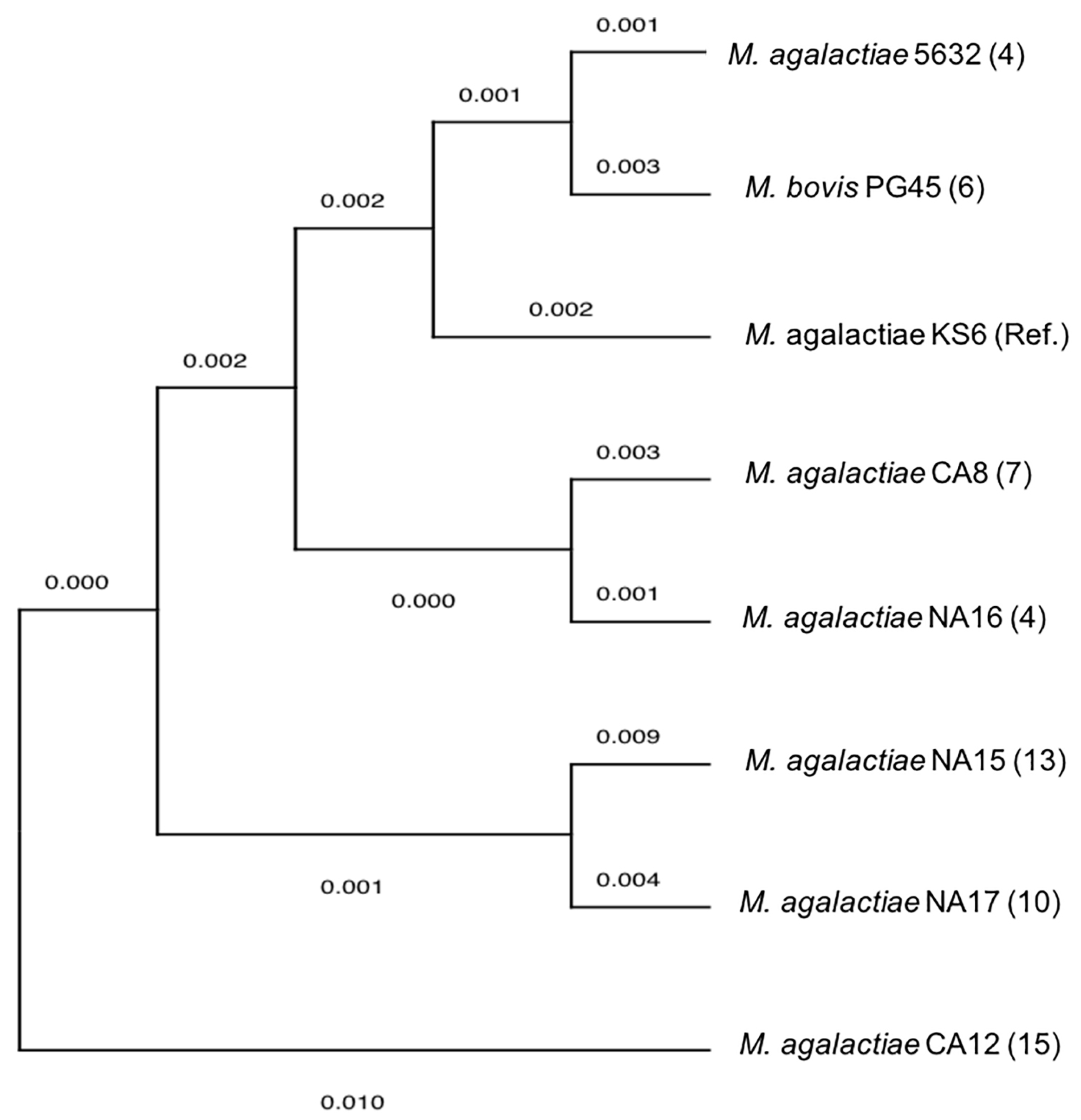
3.5. DNA Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- More, S.; Bicout, D.; Botner, A.; Butterworth, A.; Calistri, P.; De Koeijer, A.; Depner, K.; Edwards, S.; Garin-Bastuji, B.; Good, M.; et al. Vector-borne diseases. EFSA J. 2017, 15, 4793. [Google Scholar] [CrossRef] [Green Version]
- Guglielmone, A.A.; Robbins, R.G.; Apanaskevitch, D.A.; Petney, T.N.; Estrada-Pena, A.; Horak, I.G. The Hard Ticks of the World (Acari: Ixodida: Ixodididae); Springer: New York, NY, USA, 2014. [Google Scholar]
- McCoy, K.D.; Léger, E.; Dietrich, M. Host specialization in ticks and transmission of tick-borne diseases: A review. Front. Cell. Infect. Microbiol. 2013, 4, 57. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Peña, A.; Bouattour, A.; Camicas, J.L.; Walker, A.R. Ticks of Domestic Animals in the Mediterranean Region: A Guide to Identification of Species; University of Zaragoza: Zaragoza, Spain, 2004. [Google Scholar]
- Migliore, S.; Puleio, R.; Nicholas, R.A.J.; Loria, G.R. Mycoplasma agalactiae: The sole cause of classical contagious agalactia? Animals 2021, 15, 1782. [Google Scholar] [CrossRef]
- Neimark, H.; Johansson, K.E.; Rikihisa, Y.; Tully, J.G. Proposal to transfer some members of the genera Haemobartonella and Eperythrozoon to the genus Mycoplasma with descriptions of ‘Candidatus Mycoplasma haemofelis’, ‘Candidatus Mycoplasma haemomuris’, ‘Candidatus Mycoplasma haemosuis’ and ‘Candidatus Mycoplasma wenyonii’. Int. J. Syst. Evol. Microbiol. 2001, 51, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Nayak, N.C.; Blownik, M.K. Goat flea (order Siphonaptera) as a possible vector for the transmission of caprine mycoplasmal polyarthritis with septicaemia. Prev. Vet. Med. 1990, 9, 259–266. [Google Scholar] [CrossRef]
- Shaw, S.E.; Kenny, M.J.; Tasker, S.; Birtles, R.J. Pathogen carriage by the cat flea Ctenocephalides felis (Bouche) in the United Kingdom. Vet. Microbiol. 2004, 102, 183–188. [Google Scholar] [CrossRef]
- Woods, J.E.; Brewer, M.M.; Hawley, J.R.; Wisnewski, N.; Lappin, M.R. Evaluation of experimental transmission of Candidatus Mycoplasma haemominutum and Mycoplasma haemofelis by Ctenocephalides felis to cats. Am. J. Vet. Res. 2005, 66, 1008–1012. [Google Scholar] [CrossRef]
- Lappin, M.R.; Griffin, B.; Brunt, J.; Riley, A.; Burney, D.; Hawley, J.; Brewer, M.M.; Jensen, W.A. Prevalence of Bartonella species, haemoplasma species, Ehrlichia species, Anaplasma phagocytophilum, and Neorickettsia risticii DNA in the blood of cats and their fleas in the United States. J. Feline Med. Surg. 2006, 8, 85–90. [Google Scholar] [CrossRef]
- Willi, B.; Boretti, F.S.; Meli, M.L.; Bernasconi, M.V.; Casati, S.; Hegglin, D.; Puorger, M.; Neimark, H.; Cattori, V.; Wengi, N.; et al. Real-time PCR investigation of potential vectors, reservoirs, and shedding patterns of feline hemotropic mycoplasmas. Appl. Environ. Microbiol. 2007, 73, 3798–3802. [Google Scholar] [CrossRef] [Green Version]
- Kamrani, A.; Parreira, V.R.; Greenwood, J.; Prescott, J.F. The prevalence of Bartonella, hemoplasma, and Rickettsia felis infections in domestic cats and in cat fleas in Ontario. Can. J. Vet. Res. 2008, 72, 411–419. [Google Scholar]
- Foley, J.E.; Pedersen, N.C. Candidatus Mycoplasma haemominutum, a low-virulence erpierythrocytic parasite of cats. Int. J. Syst. Evol. Microbiol. 2001, 51, 815–817. [Google Scholar] [CrossRef] [Green Version]
- Willi, B.; Boretti, F.S.; Cattori, V.; Tasker, S.; Meli, M.L.; Reusch, C.; Lutz, H.; Hofmann-Lehmann, R. Identification, molecular characterization, and experimental transmission of a new hemoplasma isolate from a cat with hemolytic anemia in Switzerland. J. Clin. Microbiol. 2005, 43, 2581–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willi, B.; Boretti, F.S.; Baumgartner, C.; Cattori, V.; Meli, M.L.; Doherr, M.G.; Reusch, C.E.; Hofmann-Lehmann, R. Feline hemoplasmas in Switzerland: Identification of a novel species, diagnosis, prevalence, and clinical importance. Schweiz. Arch. Tierheilk. 2006, 148, 139–144. [Google Scholar] [CrossRef]
- Seneviratna, P.; Weerasinghe, N.; Ariyadasa, S. Transmission of Haemobartonella canis by the dog tick, Rhipicephalus sanguineus. Res. Vet. Sci. 1973, 14, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Novacco, M.; Meli, M.L.; Gentilini, F.; Marsilio, F.; Ceci, C.; Pennisi, M.G.; Lombardo, G.; Lloret, A.; Santos, L.; Carrapiço, T.; et al. Prevalence and geographical distribution of canine hemotropic mycoplasma infections in Mediterranean countries and analysis of risk factors for infection. Vet. Microbiol. 2010, 142, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Micsutka, A.; Meli, M.L.; Lutz, H.; Hofmann-Lehmann, R. Molecular investigation of transplacental and vector-borne transmission of bovine haemoplasmas. Vet. Microbiol. 2011, 152, 411–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagawa, M.; Matsumoto, K.; Inokuma, H. Molecular detection of Mycoplasma wenyonii and ‘Candidatus Mycoplasma haemobos’ in cattle in Hokkaido, Japan. Vet. Microbiol. 2008, 132, 177–180. [Google Scholar] [CrossRef]
- Ayling, R.D.; Bisgaard-Frantzen, S.; Adler, A.; Blowey, R.W.; Barlow, A.M.; Millar, M.F.; Van Der Burgt, G.M. Detection of’ Candidatus Mycoplasma haemobos’, Mycoplasma wenyonii and Anaplasma phagocytophilum from cattle in England. Vet. Rec. 2012, 170, 543. [Google Scholar] [CrossRef]
- Girotto, A.; Zangirólamo, A.F.; Bogado, A.L.; Souza, A.S.; Silva, G.C.; Garcia, J.L.; Vilas Boas, L.A.; Biondo, A.W.; Vidotto, O. Molecular detection and occurrence of’ Candidatus Mycoplasma haemobos’ in dairy cattle of Southern Brazil. Rev. Bras. Parasitol. Vet. 2012, 21, 342–344. [Google Scholar] [CrossRef]
- Hornok, S.; Sugár, L.; Fernández de Mera, I.G.; De La Fuente, J.; Horváth, G.; Kovács, T.; Micsutka, A.; Gönczi, E.; Flaisz, B.; Takács, N.; et al. Tick-and fly-borne bacteria in ungulates: The prevalence of Anaplasma phagocytophilum, haemoplasmas and rickettsiae in water buffalo and deer species in Central Europe, Hungary. BMC Vet. Res. 2018, 14, 98. [Google Scholar] [CrossRef]
- Santos, N.J.; Brito, D.R.; Abate, H.L.; Paixão, S.F.; Soares, E.D.; Vieira, T.S.; Garcia, J.L.; Vieira, R.F.; Vidotto, O. Hemotropic mycoplasmas infection in water buffaloes (Bubalus bubalis) from northeastern Brazil. Comp. Immunol. Microbiol. Infect. Dis. 2018, 56, 27–29. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Hu, Y.; Leng, C.; Shi, H.; Jiao, Z.; Chen, X.; Peng, Y.; Yang, H.; Kan, Y.; Yao, L. Molecular investigation of “Candidatus Mycoplasma haemobos” in goats and sheep in central China. Transbound. Emerg. Dis. 2019, 66, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Neimark, H.; Hoff, B.; Ganter, M. Mycoplasma ovis comb. nov. (formerly Eperythrozoon ovis), an eperythrocytic agent of haemolytic anaemia in sheep and goats. Int. J. Syst. Evol. Microbiol. 2004, 54, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Bergonier, D.; Berthelot, X.; Poumarat, F. Contagious agalactia of small ruminants: Current knowledge concerning epidemiology, diagnosis and control. Rev. Sci. Tech. 1997, 16, 848–873. [Google Scholar] [CrossRef] [PubMed]
- Loria, G.R.; Nicholas, R.A. Contagious agalactia: The shepherd’s nightmare. Vet. J. 2013, 198, 5–6. [Google Scholar] [CrossRef]
- Jay, M.; Tardy, F. Contagious agalactia in sheep and goats: Current perspectives. Vet. Med. Res. Rep. 2019, 10, 229–247. [Google Scholar] [CrossRef]
- Walker, J.B.; Keirans, J.E.; Horak, I.G. The genus Rhipicephalus (Acari, Ixodidae): A guide to the Brown Ticks of the World; Cambridge University Press: Cambridge, UK, 2000; p. 643. ISBN 9780521019774. [Google Scholar]
- Eterpi, M.; McDonnell, G.; Thomas, V. Decontamination efficacy against Mycoplasma. Appl. Microbiol. 2011, 52, 150–151. [Google Scholar] [CrossRef]
- Friis, N.F. Some recommendations concerning primary isolation of Mycoplasma suipneumoniae and Mycoplasma flocculare a survey. Nord. Vet. Med. 1975, 27, 337–339. [Google Scholar]
- Nicholas, R.; Baker, S. Recovery of mycoplasmas from animals. Methods Mol. Biol. 1998, 104, 37–43. [Google Scholar] [CrossRef]
- Pettersson, B.; Leitner, T.; Ronaghi, M.; Bölske, G.; Uhlen, M.; Johansson, K.E. Phylogeny of the Mycoplasma mycoides cluster as determined by sequence analysis of the 16S rRNA genes from the two rRNA operons. J. Bacteriol. 1996, 178, 4131–4142. [Google Scholar] [CrossRef] [Green Version]
- Torina, A.; Alongi, A.; Naranjo, V.; Estrada-Peña, A.; Vicente, J.; Scimeca, S.; Marino, A.M.; Salina, F.; Caracappa, S.; de la Fuente, J. Prevalence and genotypes of Anaplasma species and habitat suitability for ticks in a Mediterranean ecosystem. Appl. Environ. Microbiol. 2008, 74, 7578–7584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torina, A.; Khoury, C.; Caracappa, S.; Maroli, M. Ticks infesting livestock on farms in Western Sicily, Italy. Exp. Appl Acarol. 2006, 38, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Raele, D.A.; Galante, D.; Pugliese, N.; De Simone, E.; Cafiero, M.A. Coxiella-like endosymbiont associated to the Anatolian brown tick Rhipicephalus bursa in Southern Italy. Microbes Infect. 2015, 17, 799–805. [Google Scholar] [CrossRef]
- Enayati, A.A.; Asgarian, F.; Sharif, M.; Boujhmehrani, H.; Amouei, A.; Vahedi, N.; Boudaghi, B.; Piazak, N.; Hemingway, J. Propetamphos resistance in Rhipicephalus bursa (Acari, Ixodidae). Vet. Parasitol. 2009, 162, 135–141. [Google Scholar] [CrossRef]
- Ferrolho, J.; Antunes, S.; Santos, A.S.; Velez, R.; Padre, L.; Cabezas-Cruz, A.; Santos-Silva, M.M.; Domingos, A. Detection and phylogenetic characterization of Theileria spp. and Anaplasma marginale in Rhipicephalus bursa in Portugal. Ticks Tick Borne Dis. 2016, 7, 443–448. [Google Scholar] [CrossRef]
- Renneker, S.; Abdo, J.; Salih, D.E.A.; Karagenc, T.; Bilgic, H.; Torina, A.; Oliva, A.G.; Campos, J.; Kullmann, B.; Ahmed, J.; et al. Can Anaplasma ovis in small ruminants be neglected any longer? Transbound. Emerg. Dis. 2013, 60 (Suppl. 2), 105–112. [Google Scholar] [CrossRef]
- Masala, G.; Chisu, V.; Foxi, C.; Socolovschi, C.; Raoult, D.; Parola, P. First detection of Ehrlichia canis in Rhipicephalus bursa ticks in Sardinia Italy. Ticks Tick Borne Dis. 2012, 3, 396–397. [Google Scholar] [CrossRef]
- Agnone, A.; La Manna, M.; Sireci, G.; Puleio, R.; Usticano, A.; Ozdemir, U.; Nicholas, R.A.J.; Chiaracane, V.; Dieli, F.; Di Marco, V.; et al. A comparison of the efficacy of commercial and experimental vaccines for contagious agalactia in sheep. Small Rum. Res. 2013, 112, 230–234. [Google Scholar] [CrossRef]
- Loria, G.R.; Puleio, R.; Agnello, S.; Marogna, G.; Nicholas, R.A.J. Can vaccines for contagious agalactia reduce disease progression in infected animals: A preliminary study? Vet. Rec. Case Rep. 2018, 6, e000715. [Google Scholar] [CrossRef]
- Gomez-Martin, A.; de la Fe, C.; Amores, J.; Sánchez, A.; Contreras, A.; Paterna, A.; Buendía, A.J.; Corrales, J.C. Anatomic location of Mycoplasma mycoides subsp. capri and Mycoplasma agalactiae in naturally infected goat male auricular carriers. Vet. Microbiol. 2012, 157, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Agnello, S.; Chetta, M.; Vicari, D.; Mancuso, R.; Manno, C.; Puleio, R.; Console, A.; Nicholas, R.A.J.; Loria, G.R. Severe outbreaks of polyarthritis in kids caused by Mycoplasma mycoides subspecies capri in Sicily. Vet. Record. 2012, 170, 416. [Google Scholar] [CrossRef] [PubMed]
- Yeruham, I.; Hadani, A.; Galker, F. The life cycle of Rhipicephalus bursa Canestrini and Fanzago,1877 (Acarina: Ixodidae) under laboratory conditions. Vet. Parasitol. 2000, 89, 109–116. [Google Scholar] [CrossRef]
- de la Fuente, J.; Antunes, S.; Bonnet, S.; Cabezas-Cruz, A.; Domingos, A.G.; Estrada-Peña, A.; Johnson, N.; Kocan, K.M.; Mansfield, K.L.; Nijhof, A.M.; et al. Tick-pathogen interactions and vector competence: Identification of molecular drivers for tick-borne diseases. Front. Cell. Infect. Microbiol. 2017, 7, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedhoff, K.T. Transmission of Babesia, in Babesiosis of Domestic Animals and Man; Ristic, M., Ed.; CRC Press: Boca Raton, FL, USA, 1988; pp. 23–52. [Google Scholar]
- Lambert, M. Contagious agalactia of sheep and goats. Rev. Sci. Tech. 1987, 6, 699–711. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Farm | Locality | Farmed Species (n.) | First Report of CA | Sampled Individuals | Collected Ticks |
|---|---|---|---|---|---|
| A | Altofonte (PA *) | Sheep (111) Goat (75) | 2017 | 1 sheep 5 goats | 45 |
| B | Borgetto (PA *) | Sheep (171) Goat (2) | 2016 | 11 sheep | 41 |
| C | Santa Lucia del Mela (ME **) | Sheep (187) Goat (300) | 2018 | 8 goats | 66 |
| Total | 12 sheep and 13 goats | 152 |
| Farm | Tick Species | No. of Ticks | Total |
|---|---|---|---|
| A | R. bursa | 45 | 45 |
| B | R. bursa | 39 | 41 |
| R. turanicus | 1 | ||
| R. hylusitanium | 1 | ||
| C | R. bursa | 45 | 66 |
| R. sanguineus | 21 |
| Farm | ELISA (No. of Positive/All Samples) | Mycoplasma spp. Isolation from Milk (No. of Positive/All Samples) | Mycoplasma spp. Isolation from Ticks Pools (No. of Positive/All Samples) | Real-time Results from Tick Pools (No. of Positive/All Pools) | ||
|---|---|---|---|---|---|---|
| M. agalactiae | M. mycoides group | M. agalactiae/ M. mycoides | ||||
| A | 6/6 | 2/6 a | 0/15 | 3/15 | 2/15 | 0 |
| B | 4/11 | 0/11 | 2/21 a | 6/21 | 0 | 3/21 |
| C | 7/8 | 0/8 | 3/21 a | 11/20 | 0 | 5/20 |
| Tot. | 17/25 | 2/25 | 5/56 | 17 b/56 | 2 b/56 | 8 b/56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galluzzo, P.; Migliore, S.; Puleio, R.; Galuppo, L.; La Russa, F.; Blanda, V.; Tumino, S.; Torina, A.; Ridley, A.; Loria, G.R. Detection of Mycoplasma agalactiae in Ticks (Rhipicephalus bursa) Collected by Sheep and Goats in Sicily (South-Italy), Endemic Area for Contagious Agalactia. Microorganisms 2021, 9, 2312. https://doi.org/10.3390/microorganisms9112312
Galluzzo P, Migliore S, Puleio R, Galuppo L, La Russa F, Blanda V, Tumino S, Torina A, Ridley A, Loria GR. Detection of Mycoplasma agalactiae in Ticks (Rhipicephalus bursa) Collected by Sheep and Goats in Sicily (South-Italy), Endemic Area for Contagious Agalactia. Microorganisms. 2021; 9(11):2312. https://doi.org/10.3390/microorganisms9112312
Chicago/Turabian StyleGalluzzo, Paola, Sergio Migliore, Roberto Puleio, Lucia Galuppo, Francesco La Russa, Valeria Blanda, Serena Tumino, Alessandra Torina, Anne Ridley, and Guido R. Loria. 2021. "Detection of Mycoplasma agalactiae in Ticks (Rhipicephalus bursa) Collected by Sheep and Goats in Sicily (South-Italy), Endemic Area for Contagious Agalactia" Microorganisms 9, no. 11: 2312. https://doi.org/10.3390/microorganisms9112312